
Analysis of the mouse Scnn1a promoter in cortical collecting duct cells and in transgenic mice
5
Promoter paper Analysis of the mouse Scnn1a promoter in cortical collecting duct cells and in transgenic mice 1 Stefan Kohler a ; 2 , Sylvain Pradervand a ; 2 , Chantal Verdumo a , Anne-Marie Me ¤rillat a , Marcelle Bens b , Alain Vandewalle b , Friedrich Beermann c , Edith Hummler a ; * a Institute of Pharmacology and Toxicology, University of Lausanne, Rue du Bugnon 27, CH-1005 Lausanne, Switzerland b INSERM 4478, Faculte ¤ de Me ¤dicine X, Bichat BP 416, Paris, France c ISREC (Swiss Institute for Experimental Cancer Research), CH-1066 Epalinges, Switzerland Received 26 February 2001; received in revised form 11 April 2001; accepted 11 April 2001 Abstract We have isolated and characterised the promoter of the mouse Scnn1a (KENaC) gene. Using transient transfections of serial deletion mutants into Scnn1a-expressing cells, we demonstrate that 1.56 kb of 5P upstream sequence is required for cell-specific expression and corticosteroid-mediated regulation. These 5P sequences are not sufficient to drive expression of a lacZ reporter gene or a rat Scnn1a cDNA in transgenic mice, where they failed to rescue Scnn1a deficiency. ß 2001 Elsevier Science B.V. All rights reserved. Keywords : Epithelial sodium channel ; Sodium transport ; Gene expression ; Promoter ; Transgenic mouse The amiloride-sensitive epithelial sodium channel (ENaC) consists of three homologous subunits (K, L and Q) encoded by the Scnn1a, Scnn1b and Scnn1c genes. ENaC mediates the rate-limiting step for sodium absorp- tion across epithelial cells that line the distal part of the renal tubule, the distal colon, the duct of several exocrine glands, and the lung airways. Mice de¢cient for KENaC develop a respiratory distress syndrome and fail to clear their lungs of £uid [1]. In the renal tubule, sodium reab- sorption by ENaC is one of the essential mechanisms in- volved in the regulation of sodium balance, blood volume and blood pressure [2,3]. Activity of the sodium channel is positively regulated by gluco- and mineralocorticoids. This is in part due to direct e¡ects on transcriptional activity of Scnn1a, Scnn1b, or/and Scnn1c gene expression [4]. In primary cultures of rat inner medullary collecting duct cells, aldosterone induced a 3^4-fold, and glucocorticoids a 5^8-fold increase of Scnn1a mRNA levels. Similarly, in A6 Xenopus kidney cells, Scnn1a mRNA was increased 4-fold by aldosterone after 24 h [5]. A genomic DNA fragment containing the ¢rst exon and the 5P £anking region of the mouse Scnn1a gene was iso- lated from a mouse genomic 129/Sv DNA library (Fig. 1A,B) [1]. Primer extension revealed two major and one minor transcription start sites at 534, 533 and 529 bp up- stream of the ATG (Fig. 1C). Several transcription initia- tion sites have been equally identi¢ed in the human and rat Scnn1a gene [6,7]. In the following, the ¢rst major transcription start site was referred to as +1 (Fig. 1B). Di¡erent lengths of 5P £anking sequence were fused to a CAT reporter gene and transfected into mouse renal cor- tical collecting duct cells (mpkCCDc14; [8]) and STO ¢- broblasts (Fig. 2). 1.56 kb of 5P £anking region enabled the strongest CAT activity. A signi¢cant activity was al- ready seen with a short promoter fragment of 76 bp, which contains one of the two TATA-like boxes (Fig. 2B). Extending the promoter fragment beyond 3762 bp (to 31558 bp) results in a further increase of CAT activity (Fig. 2B). This promoter fragment contains the highly conserved glucocorticoid-responsive element (GRE) con- sensus site (see below ; Fig. 3A) and four additional GRE half sites. Both the rat and human Scnn1a promoter contain a GRE that confers activation of a reporter gene by dexa- methasone in human, mouse and monkey cell lines [7,9] (Fig. 3B). In the mouse Scnn1a promoter, a putative GRE (AGAACAGAATGTCCT) which closely matches the 0167-4781 / 01 / $ ^ see front matter ß 2001 Elsevier Science B.V. All rights reserved. PII:S0167-4781(01)00228-7 * Corresponding author. Fax: +41-21-692-5355; E-mail : [email protected] 1 The nucleotide sequence reported in this paper has been submitted to the NIH database under accession no. AF346304. 2 These authors contributed equally to the work. Biochimica et Biophysica Acta 1519 (2001) 106^110 www.bba-direct.com
-
Upload
stefan-kohler -
Category
Documents
-
view
213 -
download
1
Transcript of Analysis of the mouse Scnn1a promoter in cortical collecting duct cells and in transgenic mice

Promoter paper
Analysis of the mouse Scnn1a promoter in cortical collecting duct cellsand in transgenic mice1
Stefan Kohler a;2, Sylvain Pradervand a;2, Chantal Verdumo a, Anne-Marie Merillat a,Marcelle Bens b, Alain Vandewalle b, Friedrich Beermann c, Edith Hummler a;*
a Institute of Pharmacology and Toxicology, University of Lausanne, Rue du Bugnon 27, CH-1005 Lausanne, Switzerlandb INSERM 4478, Faculte de Medicine X, Bichat BP 416, Paris, France
c ISREC (Swiss Institute for Experimental Cancer Research), CH-1066 Epalinges, Switzerland
Received 26 February 2001; received in revised form 11 April 2001; accepted 11 April 2001
Abstract
We have isolated and characterised the promoter of the mouse Scnn1a (KENaC) gene. Using transient transfections of serial deletionmutants into Scnn1a-expressing cells, we demonstrate that 1.56 kb of 5P upstream sequence is required for cell-specific expression andcorticosteroid-mediated regulation. These 5P sequences are not sufficient to drive expression of a lacZ reporter gene or a rat Scnn1a cDNAin transgenic mice, where they failed to rescue Scnn1a deficiency. ß 2001 Elsevier Science B.V. All rights reserved.
Keywords: Epithelial sodium channel; Sodium transport ; Gene expression; Promoter; Transgenic mouse
The amiloride-sensitive epithelial sodium channel(ENaC) consists of three homologous subunits (K, L andQ) encoded by the Scnn1a, Scnn1b and Scnn1c genes.ENaC mediates the rate-limiting step for sodium absorp-tion across epithelial cells that line the distal part of therenal tubule, the distal colon, the duct of several exocrineglands, and the lung airways. Mice de¢cient for KENaCdevelop a respiratory distress syndrome and fail to cleartheir lungs of £uid [1]. In the renal tubule, sodium reab-sorption by ENaC is one of the essential mechanisms in-volved in the regulation of sodium balance, blood volumeand blood pressure [2,3]. Activity of the sodium channel ispositively regulated by gluco- and mineralocorticoids. Thisis in part due to direct e¡ects on transcriptional activity ofScnn1a, Scnn1b, or/and Scnn1c gene expression [4]. Inprimary cultures of rat inner medullary collecting ductcells, aldosterone induced a 3^4-fold, and glucocorticoidsa 5^8-fold increase of Scnn1a mRNA levels. Similarly, inA6 Xenopus kidney cells, Scnn1a mRNA was increased4-fold by aldosterone after 24 h [5].
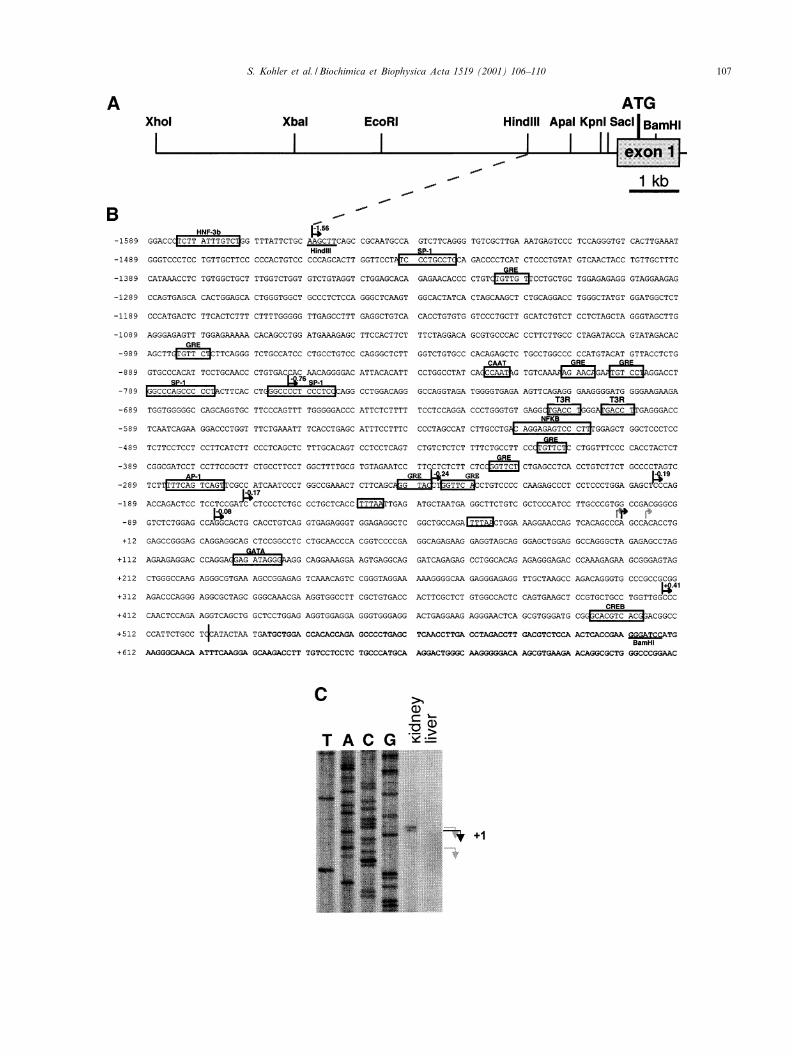
A genomic DNA fragment containing the ¢rst exon andthe 5P £anking region of the mouse Scnn1a gene was iso-lated from a mouse genomic 129/Sv DNA library (Fig.1A,B) [1]. Primer extension revealed two major and oneminor transcription start sites at 534, 533 and 529 bp up-stream of the ATG (Fig. 1C). Several transcription initia-tion sites have been equally identi¢ed in the human andrat Scnn1a gene [6,7]. In the following, the ¢rst majortranscription start site was referred to as +1 (Fig. 1B).Di¡erent lengths of 5P £anking sequence were fused to aCAT reporter gene and transfected into mouse renal cor-tical collecting duct cells (mpkCCDc14; [8]) and STO ¢-broblasts (Fig. 2). 1.56 kb of 5P £anking region enabledthe strongest CAT activity. A signi¢cant activity was al-ready seen with a short promoter fragment of 76 bp,which contains one of the two TATA-like boxes (Fig.2B). Extending the promoter fragment beyond 3762 bp(to 31558 bp) results in a further increase of CAT activity(Fig. 2B). This promoter fragment contains the highlyconserved glucocorticoid-responsive element (GRE) con-sensus site (see below; Fig. 3A) and four additionalGRE half sites.
Both the rat and human Scnn1a promoter contain aGRE that confers activation of a reporter gene by dexa-methasone in human, mouse and monkey cell lines [7,9](Fig. 3B). In the mouse Scnn1a promoter, a putative GRE(AGAACAGAATGTCCT) which closely matches the
0167-4781 / 01 / $ ^ see front matter ß 2001 Elsevier Science B.V. All rights reserved.PII: S 0 1 6 7 - 4 7 8 1 ( 0 1 ) 0 0 2 2 8 - 7
* Corresponding author. Fax: +41-21-692-5355;E-mail : [email protected]
1 The nucleotide sequence reported in this paper has been submitted tothe NIH database under accession no. AF346304.
2 These authors contributed equally to the work.
BBAEXP 91553 29-5-01
Biochimica et Biophysica Acta 1519 (2001) 106^110
www.bba-direct.com
BBAEXP 91553 29-5-01
S. Kohler et al. / Biochimica et Biophysica Acta 1519 (2001) 106^110 107

consensus sequence (AGAACANNNTGTTCT) is presentat 3811 bp (Fig. 3A). Putative GRE half sites are found at31324 bp, 3983 bp, 3416 bp, 3325 bp, 3234 bp and3241 bp (Fig. 1B). We therefore assessed hormone-medi-ated activation of expression from Scnn1a^CAT fusionconstructs. A 3-fold increase in CAT activity is alreadyseen with 236 bp of £anking sequences (Fig. 3B) whichsuggests that imperfect GRE half sites (like GGTTCTand TGTTCT) could equally respond to corticosteroidhormones. Using 1.56 kb of 5P sequence, a 6-fold (aldo-sterone-induced) and 8-fold (dexamethasone-induced)stimulation of CAT activity were obtained thus highlight-ing the importance of the consensus GRE site (Fig. 3B).The induction by corticosteroids is in accordance withstudies on human and rat Scnn1a promoter [7,10]. In hu-man lung cells, the corresponding GRE in the humanSCNN1A promoter was shown to bind glucocorticoidhormones and mediate ENaC-dependent Na� transportfollowing oxidative stress [9,11]. In comparison to therat and human SCNN1A promoter, the mouse Scnn1a
promoter shows high conservation of almost all identi¢edconsensus sequences, including CREs, TATA-like sequen-ces and the nucleotides comprising the three transcription-al start sites. We therefore suggested that 1.56 kb of 5P£anking sequences from the mouse Scnn1a gene containthe sequences necessary for stimulation by corticosteroidhormones.
From these results, we suggested that the Scnn1a pro-moter sequences could be useful in the mouse to speci¢-cally target transgene expression to Scnn1a-expressingtissues, as for example the cortical collecting duct orthe airways of the lung. To test this assumption, we¢rst analysed transgenic mouse lines carrying 1.56 kbof 5P sequences linked to the lacZ reporter gene(mScnn1a(31.56)^lacZ; not shown [12]). No X-gal stain-ing was detected in tissue obtained from four independentlacZ-transgenic lines [13]. Using reverse transcription(RT)-PCR, expression of Scnn1a(31.56)^lacZ was de-tected in kidney, colon, lung, heart and liver. The secondtransgenic experiment was performed to functionally re-
Fig. 1. The 5P £anking sequence of the mouse Scnn1a gene. A: Scheme of the 5P £anking region and the ¢rst exon containing the translation start site(ATG). Restriction sites used for generating the promoter fragments are indicated. B: Nucleotide sequence of 1589 bp of 5P region and part of exon 1.Scnn1a^CAT fusion plasmids were constructed using a XhoI site generated by PCR (Pfu Taq polymerase, Stratagene) 10 bp 5P of the ATG. Constructswere obtained using exonuclease digestion or restriction sites as indicated. A CAAT-like box (3826 bp) and TATA-like boxes (at 329 bp and 3149bp) are indicated. Potential binding sites for SP1 (31441 bp, 3789 bp and 3766 bp), T3R (3614 bp), NF-UB (3520 bp), AP-1 (3285 bp), GATA(+129 bp) and CREB (+495 bp) are indicated (identi¢ed by TFSEARCH). The position and size of the promoter fragments used for transient transfec-tion studies (see Fig. 3) are indicated (bent arrow), as well as the location of the engineered XhoI site (vertical bar). C: Primer extension from mRNAfrom mouse kidney revealed three transcription start sites. The major transcription start site at position +1 (black arrow) and two other transcriptionstart sites (grey arrow) were determined by primer extension using [K-32P]ATP end-labeled antisense primer 5P-CTCTAGCCCTGGCCTCCAGCTCCC-3P hybridised overnight at 60³C to 4 Wg of poly(A)� RNA as described [17].6
Fig. 2. Cell type-speci¢c expression from the mouse Scnn1a promoter. CAT activity of Scnn1a deletion constructs (A,B) in mouse cortical collectingduct (CCD) and ¢broblast (STO) cells. In ¢broblasts, a basal activity was observed with all promoter constructs. Transfection was performed usingTransFast1 (Promega). The plasmid molar ratio and total amount of transfected DNA were kept constant by supplementing with the pBLCAT6 basicvector [18]. Experiments were performed in triplicate, and 0.5 Wg of RSV-Luc plasmid [19] was included to normalise for transfection e¤ciency (Lucifer-ase Assay System, Promega). All plates were harvested 16 h after transfection. In A, the activity is expressed as relative CAT activity calculated by arbi-trarily setting the activity of pBLCAT6 basic vector to 1. In B, CAT activity of deletion series from the pScnn1a(31.56)^CAT constructs was arbitrarilyset to 100%.
BBAEXP 91553 29-5-01
S. Kohler et al. / Biochimica et Biophysica Acta 1519 (2001) 106^110108

veal correct Scnn1a expression in vivo. 1.56 kb of 5P £ank-ing sequences were linked to the rat Scnn1a cDNA(mScnn1a(31.56)^rScnn1a [14]) (Fig. 4A) and were bredinto a Scnn1a-de¢cient genetic background to reveal com-plementation of perinatal Scnn1a de¢ciency [1]. This strat-egy had been successfully used in a previous experiment[15]. In our study, two transgenic lines were analysed, butamongst 37 (line #1) and 32 o¡spring (line #2; Fig. 4B),no transgenic rescued animal could be identi¢ed, eventhough RT-PCR analysis revealed ubiquitous expressionof the transgene. Our results therefore suggest that 1.56
kb of 5P £anking sequences are not su¤cient to reproducethe endogenous Scnn1a expression pattern in vivo. Hence,further 5P or 3P regulatory sequences are required, whichmight be identi¢ed by co-injections of Scnn1a BACs (bac-terial arti¢cial chromosomes) with minimal reporter trans-genes [16]. Having identi¢ed the regulatory region willhelp to understand the mechanism underlying Scnn1agene expression, and ¢nally the implication of functionalENaC channels in electrolyte homeostasis.
This work has been supported by the Swiss NationalScience Foundation (Grant #31-52943.97 to E.H.). We
Fig. 3. Induction of the mouse Scnn1a promoter by corticosteroids. A: The GRE consensus sequence is highly conserved in human, mouse and ratScnn1a 5P sequence. A stretch of the mouse Scnn1a gene (positions 3821 to 3790 bp) containing the conserved GRE consensus sequence demonstrateshigh homology to rat (positions 3803 to 3772) and human (positions 32446 to 32415 bp) Scnn1a promoter sequences [10]. Identical nucleotide se-quences are indicated (grey boxes). B: Aldosterone (1 WM) and dexamethasone (100 nM) increase CAT activity in cells transfected with deletion con-structs. To maximise the response of cells to aldosterone (1036 M) or dexamethasone (1037 M) treatment, the cells were serum-starved for 24 h (2%stripped serum) before adding the steroid in a medium lacking serum, insulin, epidermal growth factor and triiodothyronine. pScnn1a(31.56)^CAT (un-stimulated) was arbitrarily set to 100% (right). *P6 0.05 versus unstimulated condition.
Fig. 4. Expression from 1.56 kb of Scnn1a regulatory sequence fails to rescue Scnn1a de¢ciency. A: Scnn1a(31.56)^rScnn1a. Scheme of the transgene(HindIII-NotI fragment) harbouring 1558 bp of 5P mouse Scnn1a £anking sequences (light grey line), the rat Scnn1a cDNA (dark grey box), and SV40small T antigen splice and polyadenylation sequences (SV40pA). Transgenic mice were identi¢ed by Southern blot analysis on tail DNA. Intron-span-ning primers used for RT-PCR analysis are indicated. B: Scnn1a^rScnn1a transgenic lines fail to rescue Scnn1a de¢ciency. The di¡erent genotypes fol-lowing mating of +/3U+/3Tg matings were determined as described previously [15]. For comparison, results of breeding of the CMV^rScnn1a trans-genic line are presented which functionally rescues mScnn1a de¢ciency [3,15]; +, wild-type; Tg, transgenic.
BBAEXP 91553 29-5-01
S. Kohler et al. / Biochimica et Biophysica Acta 1519 (2001) 106^110 109

thank Bernard C. Rossier for his continuous support onthis project.
References
[1] E. Hummler, P. Barker, J. Gatzy, F. Beermann, C. Verdumo, A.Schmidt, R. Boucher, B.C. Rossier, Nat. Genet. 12 (1996) 325^328.
[2] E. Hummler, J.-D. Horisberger, Am. J. Physiol. 276 (1999) G567^G571.
[3] E. Hummler, F. Beermann, J. Am. Soc. Nephrol. 11 (2000) S129^S134.
[4] K.A. Volk, R.D. Sigmund, P.M. Snyder, F.J. McDonald, M.J.Welsh, J.B. Stokes, J. Clin. Invest. 96 (1995) 2748^2757.
[5] A. May, A. Puoti, H.-P. Gaeggeler, J.-D. Horisberger, B.C. Rossier,J. Am. Soc. Nephrol. 8 (1997) 1813^1822.
[6] C.P. Thomas, S. Auerbach, J.B. Stokes, K.A. Volk, Am. J. Physiol.274 (1998) C1312^C1323.
[7] G. Otulakowski, B. Fa¢i, H.R. Bremner, H. O'Brodovich, Am. J.Respir. Cell Mol. Biol. 20 (1999) 1028^1040.
[8] M. Bens, V. Vallet, F. Cluzeaud, L. Pascual-Letallec, A. Kahn, M.Rafestin-Oblin, B.C. Rossier, A. Vandewalle, J. Am. Soc. Nephrol.10 (1999) 923^934.
[9] R. Sayegh, S.D. Auerbach, X. Li, R.W. Loftus, R.F. Husted, J.B.Stokes, C.P. Thomas, J. Biol. Chem. 274 (1999) 12431^12437.
[10] Y.H. Chow, Y. Wang, J. Plumb, H. O'Brodovich, J. Hu, Pediatr.Res. 46 (1999) 208^214.
[11] H.-C. Wang, M.D. Zentner, H.-T. Deng, K.-J. Kim, R. Wu, P.-C.Yang, D.K. Ann, J. Biol. Chem. 275 (2000) 8600^8609.
[12] E.H. Mercer, G.W. Hoyle, R.P. Kapur, R.L. Brinster, R.D. Palmiter,Neuron 7 (1991) 703^716.
[13] A. Schmidt, K. Tief, A. Foletti, A. Hunziker, D. Penna, E. Hummler,F. Beermann, Brain Res. Protoc. 3 (1998) 54^60.
[14] C.M. Canessa, J.D. Horisberger, B.C. Rossier, Nature 361 (1993)467^470.
[15] E. Hummler, P. Barker, C. Talbot, Q. Wang, C. Verdumo, B. Grubb,J. Gatzy, M. Burnier, J.-D. Horisberger, F. Beermann, R. Boucher,B.C. Rossier, Proc. Natl. Acad. Sci. USA 94 (1997) 11710^11715.
[16] R.J. DiLeone, G.A. Marcus, M.D. Johnson, D.M. Kingsley, Proc.Natl. Acad. Sci. USA 97 (2000) 1612^1617.
[17] S.L. McKnight, R. Kingsbury, Science 217 (1882) 316^324.[18] M. Boshart, M. Klu«ppel, A. Schmidt, G. Schu«tz, B. Luckow, Gene
110 (1992) 129^130.[19] J.R. De Wet, K.V. Wood, M. Deluca, D.R. Helinski, S. Subramani,
Mol. Cell. Biol. 7 (1987) 725^737.
BBAEXP 91553 29-5-01
S. Kohler et al. / Biochimica et Biophysica Acta 1519 (2001) 106^110110



![Activity of a maize ubiquitin promoter in transgenic riceWu [53], except that protoplasts were placed on 0.45~m Millipore filters (Millipore, Bedford, MA) over a layer of feeder nurse](https://static.fdocuments.in/doc/165x107/5feb6d5c683d19142d128b0d/activity-of-a-maize-ubiquitin-promoter-in-transgenic-rice-wu-53-except-that-protoplasts.jpg)














