
Analysis of Secondary Metabolites from Aspergillus ...784360/FULLTEXT04.pdf · Analysis of...
58
ACTA UNIVERSITATIS UPSALIENSIS UPPSALA 2015 Digital Comprehensive Summaries of Uppsala Dissertations from the Faculty of Pharmacy 195 Analysis of Secondary Metabolites from Aspergillus fumigatus and Penicillium nalgiovense Antimicrobial Compounds from Filamentous Fungi Isolated from Extreme Environments STEFAN SVAHN ISSN 1651-6192 ISBN 978-91-554-9154-3 urn:nbn:se:uu:diva-242611
Transcript of Analysis of Secondary Metabolites from Aspergillus ...784360/FULLTEXT04.pdf · Analysis of...

ACTAUNIVERSITATIS
UPSALIENSISUPPSALA
2015
Digital Comprehensive Summaries of Uppsala Dissertationsfrom the Faculty of Pharmacy 195
Analysis of Secondary Metabolitesfrom Aspergillus fumigatus andPenicillium nalgiovense
Antimicrobial Compounds from Filamentous FungiIsolated from Extreme Environments
STEFAN SVAHN
ISSN 1651-6192ISBN 978-91-554-9154-3urn:nbn:se:uu:diva-242611

Dissertation presented at Uppsala University to be publicly examined in BMC sal B22,Husargatan 4, Uppsala, Friday, 20 March 2015 at 13:00 for the degree of Doctor ofPhilosophy. The examination will be conducted in English. Faculty examiner: ProfessorDouglas Kinghorn.
AbstractSvahn, S. 2015. Analysis of Secondary Metabolites from Aspergillus fumigatus andPenicillium nalgiovense. Antimicrobial Compounds from Filamentous Fungi Isolatedfrom Extreme Environments. Digital Comprehensive Summaries of Uppsala Dissertationsfrom the Faculty of Pharmacy 195. 57 pp. Uppsala: Acta Universitatis Upsaliensis.ISBN 978-91-554-9154-3.
This thesis describes the cultivation and extraction of filamentous fungi isolated from extremeenvironments in the search for new antibiotic compounds. Filamentous fungi are a rich source ofmedicines including antibiotics, and it is believed that many currently unknown fungal speciesand bioactive fungal metabolites remain to be discovered.
Aspergillus fumigatus and Penicillium nalgiovense strains were isolated from an antibiotic-contaminated riverbed near Hyderabad, India, and soil taken from a penguin’s nest on PauleteIsland, Antarctica, respectively. It was anticipated that the extreme conditions within theseenvironments would exert unusual selective pressures on their filamentous fungi, possiblycausing the secretion of new bioactive compounds.
The cultivation, extraction and analysis of metabolites from the A. fumigatus strain resultedin the isolation of the antimicrobial substance gliotoxin. Subsequent investigations revealed thatthis strain’s secretion of gliotoxin was increased by as much as 65 % when it was cultivated inthe presence of pathogen-associated molecular patterns. These results indicate the existence of afungal receptor/signaling system for detecting nearby bacteria. The scope for using gliotoxin andthe related metabolite bis(methyl)gliotoxin as biomarker metabolites for diagnosing the lethalpulmonary condition invasive aspergillosis was also investigated. Bronchoalveolar lavage fluidfrom 42 patients with and without possible invasive aspergillosis was extracted and analyzed.The results obtained suggest that gliotoxin and bis(methyl)gliotoxin are not suitable markers fordiagnosing invasive aspergillosis.
Studies on the P. nalgiovense strain from Antarctica resulted in the isolation of the antifungalagent amphotericin B. The secretion of this compound increased when P. nalgiovense wascultured on a potato-dextrose agar enriched with coconut flakes rather than liquid RPMI 1640medium. This was the first time amphotericin B was isolated from any organism other than thebacterium Streptomyces nodosus.
The results presented in this thesis will be useful in the continuing search for novel bioactivecompounds, the diagnosis of fungal infections, and as a source of insight into the interactionsbetween microorganisms. Moreover, they show that even extensively studied fungal genera suchas Aspergillus and Penicillium are not completely understood and may produce unexpected orpreviously unknown bioactive metabolites under appropriate conditions.
Keywords: Aspergillus fumigatus, Penicillium nalgiovense, secondary metabolites, invasiveaspergillosis, elicitation, gliotoxin, bis(methyl)gliotoxin, amphotericin B
Stefan Svahn, Department of Medicinal Chemistry, Division of Pharmacognosy, Box 574,Uppsala University, SE-75123 Uppsala, Sweden.
© Stefan Svahn 2015
ISSN 1651-6192ISBN 978-91-554-9154-3urn:nbn:se:uu:diva-242611 (http://urn.kb.se/resolve?urn=urn:nbn:se:uu:diva-242611)

Do what works. Do what matters.


List of Papers
This thesis is based on the following papers, which are referred to in the text by their Roman numerals.
I Svahn, K. S., Göransson, U. El-Seedi, H., Bohlin, L., Larsson, J. D. G., Olsen, B., Chryssanthou, E. (2012) Antimicrobial activity of fil-amentous fungi isolated from highly antibiotic-contaminated river sediment. Infection Ecology and Epidemiology, 5;11591 – http//dx.doi.org/10.3402/iee.v2i0.11591.
II Svahn, K. S., Chryssanthou, E., Olsen, B., Bohlin, L., Göransson, U. (2015) Penicillium nalgiovense Laxa isolated from Antarctica is a new source of the antifungal metabolite amphotericin B. Fungal bi-ology and biotechnology, 2:1 Doi 10.1186/s40694-014-0011-x.
III Svahn, K. S., Göransson, U., Chryssanthou, E., Olsen, B., Sjölin, J., Strömstedt, A. A. (2014) Induction of gliotoxin secretion in Asper-gillus fumigatus by bacteria-associated molecules. PLoS ONE 9(4): e93685. Doi:101371/journal.pone.0093685
IV Svahn, K. S., Göransson, U., Chryssanthou, E., Bohlin, L., Janson, C., Olsen, B. Bis(methyl)gliotoxin and gliotoxin in bronchoalveolar lavage fluids are not suitable markers for invasive aspergillosis. Manuscript.
Reprints were made with permission from the respective publishers.

Author’s contribution
I. Major part in planning most experiments, reviewing the literature, cultivation, chemical analysis, evaluation, and writing. I was neither involved in sample collection nor the isolation of fungal species.
II. Major part in planning most experiments, reviewing the literature, cultivation, chemical analysis, evaluation, and writing. I was neither involved in sample collection nor the isolation of fungal species.
III. Major part in planning all experiments, reviewing the literature, cul-tivation, chemical analysis, evaluation, and writing.
IV. Major part in planning all experiments, reviewing the literature, chemical analysis, evaluation, and writing. I was neither involved in the collection of BAL samples nor diagnosing.

Contents
Introduction ..................................................................................................... 9 Pharmacognosy ........................................................................................... 9 The search for new antibiotics .................................................................... 9 Filamentous fungi and their metabolites .................................................. 11 Elicitation of fungal metabolites ............................................................... 13 Invasive aspergillosis ................................................................................ 14
Aim ................................................................................................................ 15
Fungal cultivation and analysis of metabolites ............................................. 16 Sampling and cultivation .......................................................................... 16 Fungal species ........................................................................................... 17 Extraction and isolation ............................................................................ 18 Identification and quantification of isolated compounds ......................... 20
Results and discussion ................................................................................... 22 Extraction of gliotoxin from A. fumigatus ................................................ 22 Extraction of amphotericin B from P. nalgiovense .................................. 27 Elicitation of gliotoxin production in A. fumigatus .................................. 30 The potential of gliotoxin and bis(methyl)gliotoxin as biomarkers for invasive aspergillosis ................................................................................ 34
Concluding remarks ...................................................................................... 39 Future perspectives ................................................................................... 41
Summary in Swedish ..................................................................................... 44 Mål och omfattning .................................................................................. 44 Isolering av gliotoxin från A. fumigatus ................................................... 45 Isolering av amphotericin B från P. nalgiovense ..................................... 45 Induktion av gliotoxinproduktion hos A. fumigatus ................................. 45 Gliotoxin och bis(metyl)gliotoxin som biomarkörer för invasiv aspergillos .................................................................................................................. 46 Framtida studier ........................................................................................ 46
Acknowledgements ....................................................................................... 49
References ..................................................................................................... 52

Abbreviations
A. fumigatus C. albicans E. coli
Aspergillus fumigatus Candida albicans Escherichia coli
ESBL FDA HPLC/UPLC LPS LTA
Extended spectrum betalactamase Food and drug administration High/Ultra performance liquid chromatography Lipopolysaccharide Lipoteichoic acid
MIC MRSA MS MS/MS m/z NMR PAMP P. aeruginosa P. nalgiovense PG RPMI medium SPE S. aureus
Minimum inhibitory concentration Methicillin-resistant Staphylococcus aureus Mass spectrometry or mass spectrometer Tandem mass spectrometry Mass-to-charge ratio Nuclear magnetic resonance Pathogen-associated molecular pattern Pseudomonas aeruginosa Penicillium nalgiovense Peptidoglycan Roswell Park Memorial Institute medium Solid phase extraction Staphylococcus aureus

9
Introduction
Pharmacognosy Pharmacognosy is the science of natural products used as drugs or for the production and discovery of drugs. The term is derived from the Greek words pharmacon (drug) and gnosis (knowledge). For millennia, humans have extracted and used substances of natural origin as penicillin, morphine, and caffeine to improve their quality of life. Many important medicines are natural products or derived from natural products [1]. More than one third (39.1 %) of all FDA-approved drugs are of natural origin, and 48.6 % of all cancer drugs registered from the 1940s until today are either natural prod-ucts or derivatives thereof [1]. With the development of modern analytical chemistry, microbiological cultivation methods, assays, and genetics during the last 50 years, the ability to isolate natural products from plants and mi-croorganisms has never been better [2,3]. Combined with molecular biology and computer modeling, this field of research will continue to be a signifi-cant source of useful bioactive compounds [4,5].
This in fact is illustrated by the recent discovery of the antibiotic texio-bactin, which was enabled by using a newly developed cultivation-independent method [6]. A diffusion culture chamber was used in the dis-covery, cultivation, and the evaluation of metabolites for a new bacterium, Eleftheria terrae. Texiobactin represents a new group of antibiotics and has activity against Gram-positive bacteria that are resistant to established anti-biotic agents. Discoveries of new antibiotic compounds are still made with traditional techniques. Anthracimycin was isolated in 2013 from a marine-derived Streptomyces strain. This metabolite represents a novel structure with activity against methicillin-resistant Staphylococcus aureus (MRSA) and Bacillus anthracis [7].
The search for new antibiotics Over the decades since their discovery, antibiotics have been overused and misused, resulting in increased levels of antibiotics in terrestrial and aquatic environments. The pharmaceutical industries, the meat industry, grain pro-duction, and health care have all contributed to this development [8]. One consequence of this spread of antibiotics into nature is the development and

10
spread of resistance genes among bacteria, as predicted by Alexander Flem-ing in 1945, when he was awarded the Noble Prize in medicine [9]. The spread of resistance is creating increasingly aggressive pathogens such as Pseudomonas aeruginosa, Klebsiella pneumoniae strains harboring plasmids encoding carbapanem-resistant metallo-beta-lactamase genes, MRSA, ex-tended spectrum beta-lactamase (ESBL) producing Escherichia coli, and Enterococcus faecium resistant against penicillin, vancomycin, and linezol-id. Even though these microbes present a global threat to humanity, the number of treatments effective against such resistant microorganisms is de-clining (Figure 1).
Figure 1. New antibacterial agents approved per 5-year period in the US from 1980 to 2009, and 2-year period from 2010-2012. Data from Center for Disease control and Prevention www.cdc.gov.
This worrying development is partly because the pharmaceutical industry has shown little interest in developing new antibiotics over the last two dec-ades. The collection of filamentous fungi and the investigation of their me-tabolites have played a central role in the search for drugs against infectious bacterial agents for decades.
This thesis describes studies on filamentous fungi collected from two ex-treme environments. The first environment was a location in the river Isa-kavagu near Hyderabad, India. The collection site is close to a wastewater treatment facility and has therefore been strongly affected by human activi-ty. This facility treats wastewater released by local pharmaceutical industries and has released alarmingly high quantities of fluoroquinolones and other antibiotics into its surroundings [10]. Metagenomic DNA sequencing indi-cated that the frequency of antibiotic resistance genes in samples from this environment was several times higher than in control samples obtained from the sediments of a Swedish river [11]. This suggests that the wastewater treatment plant is creating a man-made extreme environment in which mi-crobes are subject to unique selective pressures. Bacteria in this environment
# of
new
ant
ibac
teri
al a
gent
s
4
8
12
16
20
‘80-’84 ‘85-’90 ‘90-’94 ‘95-’99 ‘00-’04 ‘05-’09 ‘10-’12
Year interval

11
are exposed to the fluoroquinolones and therefore likely to develop re-sistance. Other microorganisms such as filamentous fungi must then adapt to both the high levels of antibiotics and the high population of fluoroquino-lone-resistant bacteria. This in turn creates a selection pressure favoring the development of fungal strains producing new classes of antimicrobial sub-stances or secreting larger quantities of known antibacterial metabolites.
The second sampling site was the soil from a penguin’s nest on Antarcti-ca. This site was less affected by human activity, but the harsh conditions make this environment unique, and its remoteness means that it is very diffi-cult to explore. Antarctica is one of the least explored terrestrial areas on Earth, and there have been few studies on its filamentous fungi or their se-cretion of bioactive substances [12]. The studied site’s high salinity, high UV-radiation, low temperature, low availability of water, and limited availa-bility of carbon and nitrogen may function as selective pressures favoring the secretion of bioactive compounds from the local microorganisms.
Several fungal species were isolated and identified from these two sites, but only two were chosen for up scaling. Based on their antimicrobial activi-ty, one Aspergillus fumigatus strain from the riverbed in Hyderabad, and one Penicillium nalgiovense strain from Antarctica were analyzed in this work.
Filamentous fungi and their metabolites Filamentous fungi are omnipresent organisms and break down organic mat-ter, releasing phosphorus, oxygen, nitrogen and carbon into the atmosphere and the soil. They also produce and secrete a wide variety of bioactive sec-ondary metabolites. Several of these metabolites have been developed into drugs, including antibiotics, vasodilators, immunosuppressants, anticancer, and anti-multiple sclerosis agents (Table 1).
Table 1. Medicines originating from filamentous fungi.
Medicine class Fungal origin Activity Reference Penicillins Penicillium rubens Antibiotic [13] Cephalosporins Cephalosporium Antibiotic [14] Griseofulvin Penicillium griseofulvum Antifungal [15] Cyclosporin A Tolypocladium inflatum Immunosuppressant [16] Fingolimod Isaria sinclairii Anti-multiple sclerosis [17] Lovastatin Brefeldin A
Aspergillus terrus Eupenicillium brefeldianum
Hypolipidemic Anticancer
[18] [19]

12
Two of the most important antibiotics are beta-lactams (Figure 2), including penicillins and cephalosporins.
Figure 2. Structures of the beta-lactams penicillin (left) and cephalosporins (right), where R1-3 represents a variety of side chains.
Drugs such as ciprofloxacin, gentamicin, aztreonam, ertapenem, and eryth-romycin are categorized within the fluoroquinolones, aminoglycosides, monobactams, carbapanems, and macrolides, respectively. Other antibiotics like vancomycin, linezolid, and tetracycline are not sorted into any of these categories. The mechanisms of action of antibiotic agents include affecting:
• Bacterial cell wall synthesis (beta-lactams, vancomycin, and baci-
tracin) [20]. Penicillins act by blocking the cross linking of pepti-doglycan via competitive inhibition of the transpeptidase enzyme. Cephalosporins disrupt the synthesis of the peptidoglycan layer of bacterial cell walls by competitive inhibition of penicillin bind-ing proteins. Vancomycin disrupts peptidoglycan cross-linkage [21].
• Nucleic acid synthesis (sulfonamides, trimethoprim, quinolones, and rifampin). Fluoroquinolones block the bacterial DNA replica-tion via inhibition of DNA gyrase [22].
• Protein synthesis. Tetracyclines, aminoglycosides, macrolides, linezolid, and streptogramins inhibit the 30s or 50s subunits in protein synthesis in bacteria, which leads to misreading of mRNA [23,24].
To date, more than 98,000 species of fungi have been characterized [25]. These represent only a fraction of the total number of fungal species in ex-istence, which is estimated to be anywhere between 0.6 and 9.9 million [25]. The low discovery rate of fungal species can partly be explained by the dif-ficulties encountered when cultivating filamentous fungi in laboratories. More than 99 % of all microorganisms in soil samples are reported to be impossible to cultivate under conventional laboratory conditions [26]. Clear-ly, this indicates that contemporary natural product isolation is not fully exploiting existing microbial biosynthetic capabilities. As such, there is still vast scope for studying microorganisms in order to identify new secondary metabolites.
N
O
S
HNR1
O
O OH
H
N
O
HNR2
O
HS
R3
O OH

13
The cultivation of filamentous fungi (and other microorganisms) usually begins with the inoculation of conidia into a growth medium, and carried out in shake flasks, bench top fermentors, or on agar plates. Modern advanced fermentors allow automated and controlled cultivation in batches of up to 100,000 L. As heterotrophic organisms, fungi require organic matter for growth and as a source of energy, and growth media can be based on e.g. potato-dextrose, malt extract, or soybean flour. Variables such as tempera-ture, humidity, pH, and shaking speed all affect the metabolism and growth of the filamentous fungi. Environmental biology has been revolutionized by the development of PCR-related and culture-independent techniques, such as metagenomics and metaproteomics. These modern analytical tools have lead to many new discoveries since they permit the analysis of biological diversi-ty almost to completion even in complex samples [27-29]. Furthermore, metaproteomic techniques can provide information about metabolic activity in cultures grown in e.g. 15N-enriched media. New bioactive microbial pep-tides can be discovered in this way by exploiting automated liquid chroma-tography and tandem mass spectrometry (LC-MS/MS) and comparing the results obtained to databases of known peptides, thus intersecting with the field of pharmacognosy [30].
Elicitation of fungal metabolites Bacteria and fungi are two phyla with a very long common history, and it is reasonable to assume that they have developed receptor systems to detect each other’s presence. Fungi, like all other organisms, have receptors that perceive their surroundings, although only a few such receptors have yet been characterized [31]. One such group of receptors in mammals is the Toll-like receptors, which detect pathogen-associated molecular patterns (PAMPs) and induces immune responses [32]. PAMPs are various bacterial cell wall and cell membrane components, and include lipopolysaccharide (LPS), peptidoglycan (PG), and lipoteichoic acid (LTA). Gram-negative bacteria are associated with LPS and PG, while Gram-positive bacteria are associated with PG and LTA. When such bacteria infect mammals, these molecules are released into the host, which senses them using pattern recog-nition receptors expressed on dendritic cells, macrophages, and neutrophils [33,34]. No equivalent fungal system has yet been identified.
The elicitation experiments in this thesis aimed to induce or provoke a metabolic response from A. fumigatus during cultivation. I propose that re-ceptor systems have been developed in filamentous fungi to detect and re-spond to nearby bacteria, just like in eukaryotes like humans, mice, and in-sects [34].

14
Invasive aspergillosis Invasive aspergillosis is a pulmonary disease that occurs when pathogenic Aspergillus sp. invades the lungs and destroys their tissues [35]. It is very rare in otherwise healthy individuals, but presents a significant risk to im-munosuppressed patients such as individuals with HIV, hematological dis-orders, cirrhosis, chronically obstructive pulmonary disease, and transplant recipients [36,37]. During infection, the conidia germinate in the alveoli and perforate the tissue, and the fungus secretes metabolites such as gliotoxin to kill the surrounding tissue and deflect the host’s immune response [38-40].
After having surveyed the metabolism of A. fumigatus in Paper I, and given the slight imperfections in existing diagnostic methods, I saw great potential in studying A. fumigatus metabolites in bronchoalveolar lavage (BAL) specimen from patients with invasive aspergillosis. The quantitative analysis of A. fumigatus metabolites in BAL samples with LC-MS is likely to be a useful means of diagnosing invasive aspergillosis

15
Aim
This thesis focuses on the analysis of antimicrobial secondary metabolites, secreted by A. fumigatus and P. nalgiovense. Initial studies on the metabo-lites produced by these species led to studies on their elicitation with patho-gen-associated molecular patterns and the scope for diagnosing invasive aspergillosis or other fungal diseases by screening bronchoalveolar lavage samples for gliotoxin and bis(methyl)gliotoxin. Specific aims were:
• To develop methods for isolating and cultivating filamentous fun-
gi from extreme environments (I and II). • To extract, isolate, and elucidate the structures of secreted bioac-
tive compounds from fungal strains showing strong antimicrobial activity (I and II).
• To elicitate fungal production of bioactive compounds by modu-lating growth conditions (II) and by culturing A. fumigatus with bacterial cell wall and cell membrane associated molecules (III).
• To improve the diagnosis of invasive aspergillosis by determining A. fumigatus metabolites in human bronchoalveolar lavage fluid (IV).
The overall aim of this thesis is to place the results presented in Papers I-
IV in a broader context, and to provide suggestions for future studies on bioactive secondary metabolites from filamentous fungi.

16
Fungal cultivation and analysis of metabolites
The cultivation, extraction, and isolation of bioactive metabolites from A. fumigatus and P. nalgiovense strains are illustrated in figure 3.
Sampling and cultivation The fungal strains were isolated from either a river sediment soil sample from the Isakavagu river near Hyderabad, India (Paper I) or the soil from a penguin’s nest on Paulete island in Antarctica (Paper II). The soil samples were immediately frozen and transported to the Karolinska University La-boratory in Stockholm in an unbroken freeze chain. The filamentous fungi were isolated by using sterile cotton swabs to spread material from the sam-ples over Sabouraud dextrose agar, potato-dextrose agar, or malt agar plates and then incubating the plates until colonies became visible. To ensure puri-ty, the fungal strains were re-cultured on potato-dextrose. The A. fumigatus strain (Paper I) was identified by microscopic examination while the P. nalgiovense strain (Paper II) was sent for identification by genotypic and phenotypic methods at the Fungal Biodiversity Centre in Utrecht, the Neth-erlands. The isolates were stored at -70 °C until required for cultivation and extraction. During the screening of fungal bioactive metabolites, the fila-mentous fungi were cultivated in RPMI 1640 broth or on solid potato-dextrose medium (Figure 4) [41,42]. The incubation time varied from one to four weeks.
Isolated fungi
Solid medium culture Liquid medium culture
Extraction and filtration
Preparative HPLC
HPLC separation
Identification with
MS & NMR
Filtration of supernatant
Preparative SPE
Assay
Assay
Figure 3. Work flow of assay-guided extraction of fungal metabolites. The right hand route was used for A. fu-migatus cultivated in liquid media (Paper I). The left hand route was only used for P. nalgiovense cultivat-ed on solid media (Paper II). In Paper III and IV, the extracts were only filtered and diluted prior to LC-MS analysis.

17
An assay for antimicrobial activity was used to screen extracts for bioactive fungal metabolites. The extracts were serially diluted in a 96-well plate, at dilutions ranging from 1/1 to 1/32. A suspension of bacteria or fungi was added to each well containing diluted extract, after which the plate was in-cubated for 48 hours and inspected to assess growth. An extract was consid-ered to show antimicrobial activity if the growth of the microorganisms was inhibited at any dilution. This assay was initially performed at the Depart-ment of Clinical Microbiology, Karolinska University Laboratory (Paper I) using both yeast and selected bacteria: Candida albicans ATCC 90028, Escherichia coli ATCC 25922, ESBL-producing E. coli 4.09, Staphylococ-cus aureus ATCC 29213, MRSA 2.08, Pseudomonas aeruginosa 97.09 (re-sistant against beta-lactam antibiotics, gentamicin, amikacin, ceftazidime, and ciprofloxacin), vancomycin-resistant Enterococcus van A V4932, and carbapanem-resistant Klebsiella pneumoniae 91.07. After 2010, these exper-iments were performed at the division of Pharmacognosy of Uppsala Uni-versity, using the following microorganisms: E. coli ATCC 25922, S. aureus ATCC 29213, P. aeruginosa ATCC 27853, and C. albicans ATCC 90028. The inhibitory activity of the fungal extracts was quantified by determining their minimum inhibitory concentration (MIC), i.e. the lowest concentration of the antimicrobial substance capable of inhibiting the growth of a microor-ganism [43].
Fungal species The work in this thesis included analysis of secondary metabolites from two strains of A. fumigatus and P. nalgiovense (Figure 4). Molds belonging to
Figure 4. Fungal cultivation. Left: Cultures of A. fumigatus (left tube) and P. nalgiovense (right tube) on potato-dextrose agar. Middle: P. nalgiovense cultured in RPMI 1640 medium at inoculation (left tube) and after 14 days (right tube). Right: P. nalgiovense cultivated on potato-dextrose agar en-riched with coconut flakes after 1, 3, 5, and 7 days (counting clockwise starting from top right plate).

18
the genus Aspergillus are ubiquitous saprophytic filamentous fungi that thrive in many environments. Certain Aspergillus species such as A. fumiga-tus, are also pathogenic and infect human lungs. The 10.000-15.000 liters of air that a typical person inhales each day are estimated to contain a few hun-dred A. fumigatus conidia [44,45]. The conidia are 2-3 µm in diameter and can therefore penetrate deep into the lungs, reaching the alveoli. A. fumiga-tus is also heat tolerant, resistant to oxidative stress, has a high growth rate, and can survive on various nutrients [46-48]. This combination of traits makes A. fumigatus one of the most pathogenic molds known [49]. Conse-quently, its metabolism, biology, and pathogenicity have been reviewed extensively [48,50], and its genome was sequenced in 2005 [51].
P. nalgiovense was first discovered in 1932 by O. Laxa [52] and is today used in the meat industry as a starting culture for sausages, producing a white-greyish coating on the surface of the meat. It is related to P. camem-bert and P. roquefortii, which are used in cheese production. P. nalgiovense is considered suitable for cheese and sausage production because it is not known to produce appreciable quantities of mycotoxins or bioactive metabo-lites other than penicillin [53] and peptaibols [54]. Penicillin was discovered in 1929 [13] and originates from the Penicillium genus, as its name suggests.
Extraction and isolation The four studies presented in this thesis used somewhat different procedures for the extraction and isolation of fungal metabolites. The standard proce-dure outlined above (Figure 3) reflects the approach adopted in Papers I and II, but a simplified extraction protocol was used in Papers III and IV. In general, the isolation of the fungal metabolites and their characterization were achieved using high or ultra high performance liquid chromatography (HPLC or UPLC), UV-detection, and mass spectrometry (MS). HPLC is a chromatographic technique that can be used analytically and preparative to separate diverse types of compounds based on their size, charge, and polari-ty, among other things. Liquid chromatographs and MS are commonly cou-pled together as a convenient way of separating and characterizing com-pound mixtures. In Paper I, A. fumigatus was cultured in RPMI 1640 medi-um and then filtered to separate the mycelial growth from the liquid. RPMI 1640 contains several ions (Na+, Cl-, K+, etc.) and D-glucose, which were separated from the secondary metabolites using C18 SPE columns augment-ed with 10 g of packing material. The columns were conditioned with 10 % aqueous acetonitrile containing 0.05 % formic acid, and the metabolites were eluted with 80 % aqueous acetonitrile containing 0.05 % formic acid. In Paper II, P. nalgiovense was cultured both in RPMI 1640 and on solid media. Three fungal extracts were considered in this work: one from the filtrate of the liquid culture, one from the mycelial growth, and a methanol

19
extract of the cultures grown on solid media. The mycelial growth and agar-based cultures were extracted twice with methanol (1:3 v/v). Fungal metabo-lites may have been produced but not secreted from the fungal cells during the cultivation. In order to disrupt the fungal cell walls and membranes, thus increase the extraction efficiency, the mycelial growth was put in an ultra-sonic bath during the extraction. In Paper III, the A. fumigatus cultures were simply filtered and then diluted in 10 % aqueous acetonitrile containing 0.05 % trifluoroacetic acid. In Paper IV, the BAL samples were extracted twice with dichloromethane (1:3 v/v), evaporated, filtered through a 0.22 µm filter centrifuge tubes, and dissolved in methanol before LC-MS analysis.
Preparative and analytical HPLC was performed on an ÄKTA basic HPLC system with a UV-900 detector. Depending on the size of the extract to be separated, separation was achieved using either a semi-preparative Jupiter C18 250 × 10 mm column or an analytical Jupiter C18 250 × 4.6 mm column (both with a particle size of with 5 µm). Different gradients of water and up to 60 % acetonitrile were used for separation (Figure 5); in most case the gradient elution was performed over 40 minutes but elution periods of up to 120 minutes were used for some extracts. All of the extracts discussed in Papers I, II and III were fractionated by HPLC, and the separated fractions were examined in antimicrobial assays. Active fractions were further ana-lyzed by MS and NMR. All BAL samples were frozen at -20 °C after collec-tion, then thawed and extracted twice with dichloromethane (1:3 v/v) for a total time of 120 minutes. The pooled extracts were evaporated and dis-solved in methanol before UPLC-MS analysis, and the chromatograms were compared to data for reference substances.
Figure 5. An illustrative HPLC-UV analysis of an extract from a S. aureus culture recorded at 215 nm. The acetonitrile content of the eluent is increased from 5 % to 60 % over the course of the analysis to elute hydrophobic substances.
Minutes0 5 10 15 20 25 30
500
1000
1500
2000
Inte
nsity
(mA
U)
60 % Acetonirile
5 % Acetonitrile

20
Identification and quantification of isolated compounds The structural elucidation of isolated compounds is essential in natural prod-uct research. Two particularly useful techniques for this purpose are MS and nuclear magnetic resonance (NMR) spectroscopy, which provide different and complementary types of structural information. NMR spectroscopy re-lies on the magnetic properties of the atom nuclei in a molecule. Each com-pound generates a unique spectrum that reveals the chemical surrounding of each atom. This, in turn, often leads to complete structure elucidation, which cannot generally be obtained by MS. The NMR measurements relevant to this thesis were obtained from a Varian 400 MHz AS400 spectrometer or a Bruker 600 MHz spectrometer equipped with a smartprobe. An MS is a highly versatile and flexible detector with a large number of applications, generating valuable structural information such as accurate mass, fragmenta-tion pattern, and isotopic signature of the analyzed compound in short time with high accuracy and sensitivity.
An electrospray ionization ion source was used in this thesis in combina-tion with a quadrupole-time-of-flight mass analyzer to detect and analyze extracted fungal metabolites. The liquid samples were pumped into a capil-lary during the electrospray ionization, subjected to a sheath gas and an elec-tric field. The sheath gas transforms the liquid into small charged aerosols and eventually into free ions, which were accelerated into the quadrupole-time-of-flight mass analyzer. The quadrupole consists of four parallel rods on which a direct current and a radio frequency are applied, shifting the po-larity of the rods. This creates an oscillating electric field, which regulates the m/z window. The MS plots the signals from the gas phase ions based on their mass-to-charge (m/z) ratio. This ratio reflects the time it takes for the ions to pass through an electric field and hit the microchannel plate detector. During tandem MS (MS/MS) analyses, the ions were led through a collision cell to be fragmented by collision with argon gas before reaching the detec-tor. Fragmentation patterns are characteristic or even unique for each com-pound, thus an important part of the compound identification. Other com-monly used MS instruments and applications include separation techniques such as gas chromatography and ion mobility spectrometry, ion sources such as fast atom bombardment and matrix assisted laser desorption/ionization, mass selectors such as ion traps and triple quadrupoles, and detectors such as electron multipliers and orbitraps.
An MS can detect much lower amounts of sample compared to a UV-detector. This allows a smaller inner diameter of the HPLC column. The capillary HPLC system (C18 150 × 1.0 mm column having a 5 µm particle size) used in Paper I was an instrument with excellent qualities for the pre-sented work. However, it was replaced with a nanoACQUITY UPLC system (250 mm × 75 µm column having a 1.7 µm particle size), which improved the sensitivity and selectivity in Papers II, III, and IV.

21
In Paper III a Mann-Whitney test was used to compare the gliotoxin con-centration in four different groups (for each group n=40, significance threshold P<0.05) of cultures; one reference group and three groups that had been exposed to different PAMPs. The reference group was cultivated ac-cording to the standard protocol while the other three were cultivated ac-cording to the standard protocol but using media containing LPS, PG, or LTA at 5 µg L-1.

22
Results and discussion
The four studies presented in this thesis involved the extraction and analysis of fungal metabolites with antimicrobial activity. Papers I and II had similar aims: to isolate fungal species from soil or sediment samples collected in extreme environments, followed by extraction, isolation, and identification of their secreted bioactive metabolites. I analyzed the metabolites secreted from a strain of the filamentous fungus P. nalgiovense in Paper II, and from the filamentous fungi A. fumigatus in Papers I, III, and IV. The results pre-sented in Paper I formed the basis of the studies presented in Papers III and IV.
Extraction of gliotoxin from A. fumigatus The first extreme environment was a riverbed sample from the river Isa-kavagu, near Hyderabad, India. The river was contaminated with high levels of ciprofloxacin and other antibiotics, and the spread of antibiotic genes among bacterial species were higher that in a control sample from a Swedish river sediment [10,11]. Sixty-one strains of filamentous fungi (Table 3) were isolated from the riverbed sample and several of them showed antimicrobial activity in the initial assays against Gram-positive and Gram-negative bacte-ria (Table 4).

23
Table 3. Fungal diversity of the 61 strains of filamentous fungi isolated from the antibiotic-contaminated river sediment.
Species Number of strains Alternaria sp. 2
Aspergillus clavatus 1 A. flavus 6
A. fumigatus 6 A. glaucus 1
A. niger 8 A. terrus 4
Aspergillus sp. 5 Fusarium sp. 5
Mucor sp. 5 Paecilomyces crustaceus 2
Penicillium sp. 1 Phoma glomerata 1
Phoma sp. 4 Rhizopus sp. 3
Trichoderma sp. 5 Unidentified zygomycete 2
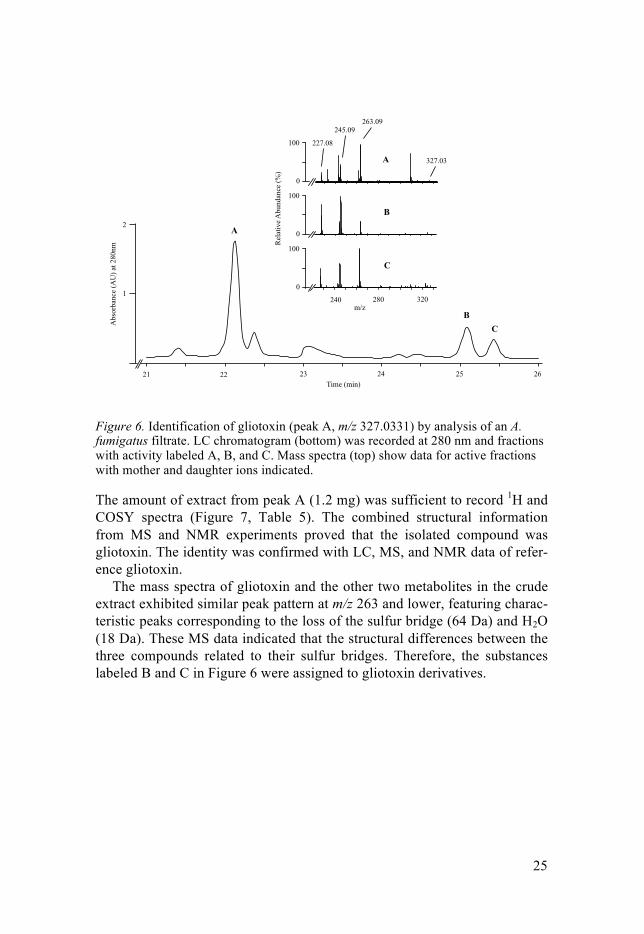
Many of the isolated fungal species qualified for further studies, but A. fu-migatus strain 1 was chosen since it had the most significant activity. The filtered culture of this strain revealed antimicrobial effect against Enterococ-cus van A, MRSA, ESBL-producing E. coli, and P. aeruginosa in dilution down to 1:512, 1:256, 1:16, and 1:1, respectively, and down to 1:8 against C. albicans. The preparative work led to the isolation of three fractions with antimicrobial activity (Figure 6).

Tabl
e 4.
The
bac
teric
idal
eff
ects
of t
he fu
ngal
cul
ture
filtr
ates
on
susc
eptib
le a
nd re
sist
ant b
acte
rial s
train
s.
Spec
ies
Isol
ate
no.
S. a
ureu
s A
TC
C 2
9213
E
. col
i A
TC
C 2
5922
E. f
aeci
um
VR
E v
an A
C
CU
G 3
6804
K. p
neum
onia
e 91
.07
MR
SA 2
.08
E. c
oli E
SBL
4.
09
A. f
umig
atus
1,
6, 1
3, 4
4, 5
0 +
+ +
+ +
+ A.
fum
igat
us
46
+ +
+ +
+ no
As
perg
illus
sp.
15
+ no
+
+ +
no
A. f
lavu
s 2,
5, 2
1, 5
8 +
+ +
+ +
+ A.
fla
vus
4 +
+ +
+ +
no
A. fl
avus
61
+
no
+ no
+
no
A. n
iger
28
, 30,
31,
32,
34
, 37,
56
+ no
+
no
+ no
Tric
hode
rma
sp.
12, 6
0 +
no
+ no
+
no
+ in
dica
tes t
hat t
he fi
ltrat
e re
duce
d ba
cter
ial g
row
th b
y ≥9
9.98
% re
lativ
e to
that
seen
in c
ontro
l cul
ture
s; n
o =
no b
acte
ricid
al a
ctiv
ity.
25
Figure 6. Identification of gliotoxin (peak A, m/z 327.0331) by analysis of an A. fumigatus filtrate. LC chromatogram (bottom) was recorded at 280 nm and fractions with activity labeled A, B, and C. Mass spectra (top) show data for active fractions with mother and daughter ions indicated.
The amount of extract from peak A (1.2 mg) was sufficient to record 1H and COSY spectra (Figure 7, Table 5). The combined structural information from MS and NMR experiments proved that the isolated compound was gliotoxin. The identity was confirmed with LC, MS, and NMR data of refer-ence gliotoxin.
The mass spectra of gliotoxin and the other two metabolites in the crude extract exhibited similar peak pattern at m/z 263 and lower, featuring charac-teristic peaks corresponding to the loss of the sulfur bridge (64 Da) and H2O (18 Da). These MS data indicated that the structural differences between the three compounds related to their sulfur bridges. Therefore, the substances labeled B and C in Figure 6 were assigned to gliotoxin derivatives.
Abs
orba
nce
(AU
) at 2
80nm
22 23 24 25 2621Time (min)
A
BC
2
1
Rel
ativ
e Abu
ndan
ce (%
)
100
0
A
B
C
227.08
245.09263.09
327.03
240 280 320m/z
100
0
100
0

26
Figure 7. NMR analysis of gliotoxin, dissolved in CDCl3, using tetramethylsilane as internal standard. Data from this COSY spectrum led to the structure elucidation and identification of gliotoxin in Paper I.
Table 5. NMR data for extracted gliotoxin and reference gliotoxin dissolved in CDCl3, recorded at 400 MHz.
Position Extracted gliotoxin Reference gliotoxin δH (J in Hz) COSY δH (J in Hz) COSY
2a 3.19, s 3.13, s 3a 4.81, s 4.75, s
3a-OH 3.15, s 3.64, br s 5a 4.41, d (13) 6 4.35, d (12) 6 6 4.27, d (13) 5a 4.18, dd (12, 7.6) 5a
6-OH 5.59, s 5.63, s 7 5.78, d (8.8) 8 5.71, d (8.4) 8 8 5.93, dd (8.8, 4.8) 7, 9 5.86, dd (8.4, 4.8) 7, 9 9 5.99, m 8 5.92, m 8, 10A
10A 3.75, dm (18) 10B 3.68, dm (18) 9, 10B 10B 2.95, d (18) 10A 2.88, d (18) 10A
The results from Paper I prove the concept of bio-assay guided screening and isolation of bioactive compounds from a filamentous fungus in an ex-treme environment. Several strains of filamentous fungi secreted metabolites with activity against some of the clinically most important multi-resistant bacterial strains, against which there are few or no effective treatment op-tions. The bioassay-guided approach used in this work is not especially la-bor-intensive but makes it possible to screen for antimicrobial compounds with activity against multiple microbes and to semi-quantitatively assess the
6 5 4 3 2
ppm
6
5
4
3
2
ppmH10A/H10B
H5/H6
H9/H8
N N
HOH
O
O
Me
OH
S S
HHH
H
H
H
H9/H8 H10A/H10B
H5/H6

27
activity of the identified metabolites due to the incorporation of multiple dilution steps. If sensitive and selective LC-MS detection is incorporated into the process, it represents a soil-to-spectrum approach to isolating new bioactive compounds. No previous attempts to isolate bioactive compounds originating from filamentous fungi collected from antibiotic-contaminated environments have been reported.
Current screening methods for the identification of antimicrobials are very diverse, and the results of a given screening campaign may be strongly dependent on the approach and methods adopted. It is possible that the ap-proach used in this work selected for specific microorganisms and metabo-lites. The studied environment may thus harbor other filamentous fungi that secrete secondary metabolites with bioactivity, only not detected with the chosen methods. However, the isolation and identification of gliotoxin con-firms the potential and value of this approach. Gliotoxin belongs to the epipolythiodioxopiperazines, which are a group of fungal secondary metabo-lites with broad activity, due to their reactive disulfur-bridged diketopipera-zine ring [55,56]. The molecule’s toxicity in mammalian cells is closely connected to the bridge, easily forming covalent linkages with other biomol-ecules [57]. The mechanism of action is not completely understood, but in-cludes the generation of reactive oxygen species by redox cycling [58] (Fig-ure 8) and attaching to proteins with cysteine or other susceptible thiol resi-dues that causes inactivation of e.g. alcoholdehydrogenase and creatine ki-nase, respectively [59,60]. For these reasons, gliotoxin is not considered a suitable drug candidate.
Figure 8. Generation of reactive oxygen species by redox cycling between the re-duced (left) and oxidized (right) forms of gliotoxin.
There is considerable potential for conducting similar studies in future be-cause there are vast numbers of unexplored and man-made extreme envi-ronments to investigate.
Extraction of amphotericin B from P. nalgiovense The second soil sample that was investigated was from Paulete Island in Antarctica, a harsh and hostile environment for most organisms. This envi-
N N
OHH
O
O
Me
OH
S SN N
OHH
O
O
Me
OH
SH
SH
2 O2 2 O2- . 2 H2O2 + HO .
28
ronment is not only extremely cold and inhospitable, but rather unexplored with respect to filamentous fungi and their secondary metabolites.
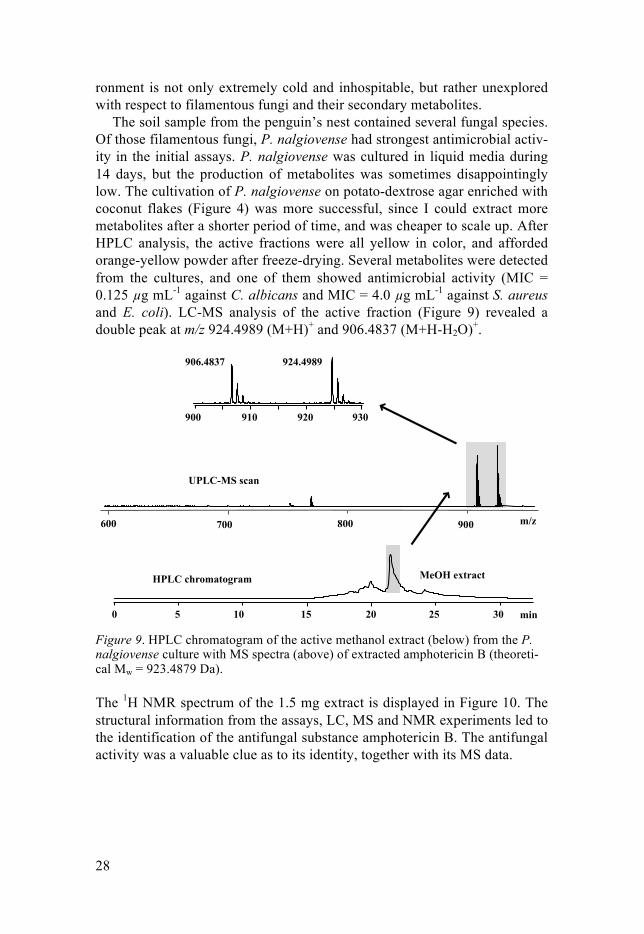
The soil sample from the penguin’s nest contained several fungal species. Of those filamentous fungi, P. nalgiovense had strongest antimicrobial activ-ity in the initial assays. P. nalgiovense was cultured in liquid media during 14 days, but the production of metabolites was sometimes disappointingly low. The cultivation of P. nalgiovense on potato-dextrose agar enriched with coconut flakes (Figure 4) was more successful, since I could extract more metabolites after a shorter period of time, and was cheaper to scale up. After HPLC analysis, the active fractions were all yellow in color, and afforded orange-yellow powder after freeze-drying. Several metabolites were detected from the cultures, and one of them showed antimicrobial activity (MIC = 0.125 µg mL-1 against C. albicans and MIC = 4.0 µg mL-1 against S. aureus and E. coli). LC-MS analysis of the active fraction (Figure 9) revealed a double peak at m/z 924.4989 (M+H)+ and 906.4837 (M+H-H2O)+.
Figure 9. HPLC chromatogram of the active methanol extract (below) from the P. nalgiovense culture with MS spectra (above) of extracted amphotericin B (theoreti-cal Mw = 923.4879 Da).
The 1H NMR spectrum of the 1.5 mg extract is displayed in Figure 10. The structural information from the assays, LC, MS and NMR experiments led to the identification of the antifungal substance amphotericin B. The antifungal activity was a valuable clue as to its identity, together with its MS data.
600 800 m/z
MeOH extract
UPLC-MS scan
HPLC chromatogram
0 5 10 15 20 25 30 min
700 900
906.4837 924.4989
900 930920910

29
Figure 10. 1H NMR spectrum of amphotericin B extracted from P. nalgiovense, and dissolved in DMSO-d6. The peaks at 2.5 and 3.3 ppm were assigned to DMSO and residual methanol, respectively.
Since amphotericin B contains 74 protons, the amount of material was not sufficient for complete proton assignment on this 600 MHz NMR spec-trometer. The crude material was therefore identified by visual comparison to a reference NMR spectrum that was convincingly similar, and MS data that produced peaks at m/z 924.4897 and 906.4729 for reference amphoteri-cin B. All spectroscopic data matched that reported previously for amphoter-icin B [61,62].
The results show that a strain of P. nalgiovense, isolated from the soil from a penguin’s nest on Antarctica, now is the second known organism being able to produce amphotericin B, after the bacterium S. nodosus. The secretion of this metabolite was noticeably enhanced when the fungus was cultivated on solid medium enriched with coconut flakes, compared to the liquid medium RPMI 1640.
P. nalgiovense has been cultivated and analyzed by several research groups that have isolated other substances, such as peptaibols, nalgiovensin, diaporthins, dipodazin, and penicillins but not amphotericin B [53,63-65]. None of the other metabolites were detected in the present study. This meta-bolic difference may depend on cultivation procedures, choice of extraction solvent, or possibly genetic differences in the fungal strains. Culture media in other studies included Czapek yeast autolysate agar, malt extract agar, oatmeal agar, and the glucose-based Raulin-Thom solution followed by ex-traction with petroleum, chloroform-methanol mixtures, or ethyl acetate. Even if P. nalgiovense strains produced amphotericin B in other studies, this metabolite may have gone unnoticed through the analysis since antifungal activity of the extracted compounds was not studied. To my knowledge, this is the first study, which use potato-dextrose agar enriched with coconut flakes for cultivation of P. nalgiovense.
The mode of action of amphotericin B is based on its affinity to ergoste-rol, which is a sterol found in fungal cell membranes. By binding to ergoste-rol with the polyene side oriented outwards, amphotericin B creates a trans-
6 5 4 3 2ppm
1
O
OH
O
O O
OHHO
NH2
HO O O
OH
OH OH
OH
OH OH
OH

30
membrane channel. The fungal cell then becomes disrupted, due to the leak-age of ions (calcium, potassium, protons, and chloride) and other small mol-ecules [66]. Although it is uncertain if amphotericin B is the primary cause, this impairment is followed by cell death [67].
No attempts were made in this study to investigate the biosynthetic path-ways for the amphotericin B production, but it is possible that the ability to produce amphotericin B is unique for this strain of P. nalgiovense, implying a genetic explanation. Lateral gene transfer between bacteria and filamen-tous fungi is not unknown, and the genes involved in the biosynthesis of amphotericin B in S. nodosus, or another amphotericin B producing organ-ism, may have been transferred into the genome of P. nalgiovense [68,69]. Alternatively, a different biosynthetic pathway to produce amphotericin B may have been developed by P. nalgiovense, compared to S. nodosus. Final-ly, a more practical explanation could be that amphotericin B has been pro-duced in other studies, although in too low amount to be detected.
Based on the results in Papers II, it is not unreasonable to expect a differ-ent set of secreted metabolites depending on cultivation procedure. Even though the possibilities of varying a culture procedure are many, this study highlighted the importance of development and re-evaluation of cultivation methods in the search for bioactive compounds from filamentous fungi. In this article, I presented a differentiated cultivation and isolation procedure (compared to paper I), and managed to significantly increase the production of the antifungal metabolite amphotericin B for the chosen fungus.
In conclusion, I showed that P. nalgiovense is untapped of its bioactive metabolites. This is promising in terms of exploiting other known and un-known fungal species, of which there are many.
Elicitation of gliotoxin production in A. fumigatus As pointed out above, the choice of medium played a significant role in the metabolism for the filamentous fungi studied in this thesis. Instead of chang-ing medium (as in Paper II), a different kind of adjustment of the cultivation condition was performed in Paper III. Under the assumption that surrounding bacteria would stimulate or trigger the filamentous fungus to produce certain metabolites, I added LPS, LTA, and PG (Figure 11) in the RPMI 1640 me-dium to simulate the presence of bacteria, and observed the changes in the metabolism of A. fumigatus.

31
Figure 11. Pathogen associated molecular patterns. Top left: Lipopolysaccharide. EtN = ethanolamine, Gal = D-galactose, Glu = glucose, Hep = L-glycero-D-manno-heptose, KDO = 3-deoxy-D-manno-oct-2-ulosonic acid, P = phosphate. The lipid A, and polysaccharide structure varies among species of Gram-negative bacteria, but the major components are typically the same (lipid A, KDO-core, O-antigen). Top right: Peptidoglycan from S. aureus, built up by N-Acetylmuramic acid (left) and N-Acetylglucosamin (right) units, where the tetra peptide chain contains of L-alanine, D-glutamine, L-lysine, and D-alanine. A 5-glycine interbridges tetrapep-tides between N-Acetylmuramic acid units to add stability to the cell wall in Gram-positive bacteria. Below: Lipoteichoic acid is bound to membrane lipids in Gram-positive bacteria and partially responsible for the negative charge of the cell surface.
The experiments included in Pater III were carried out in three steps and built on the cultivation procedure of A. fumigatus developed in Paper I. The first step was to determine within which concentration intervals the PAMPs affected the fungus. The three PAMPs were individually added to the cul-tures at 0.3, 1, 5, 10, and 100 µg per mL and measured after seven days (Figure 12).
HO OP
O O
HO
O
O-O
NH3+H3C
PO
O
O-
O
HO
HO OH
O O
HO
HO OH
O
O
O
C15H31
O
O
C15H31
45 -50
O
O
O
O
O
NH
O
O
O
OO
O HN OP
O
OHO-
O
OH
O
HO
HO
HepPEtN
Hep
KDO
KDO
PEtN
P Hep
Glu
Glu
Glu Hep
Gal
KDO
O-Antigen
O-HO
O
PO
Lipid A
Core oligo-saccharide
O
OO
HO
O*
NHO
HO
OH
O
O
Tetrapeptide
NH
O
O
*
Lipopolysaccharide
Peptidoglycan
Lipoteichoic acid

32
Figure 12. The influence of PAMPs on the secretion of gliotoxin in A. fumigatus after seven days of incubation. The levels of gliotoxin are normalized against a con-trol group (38 µM) and the error bars show the standard error of the reported mean (n=3). The levels of gliotoxin were increased by up to 85 %, starting from a PAMP concentration at 0.1 µg mL-1.
The levels of secreted gliotoxin increased by up to 85 %, starting from a PAMP concentration at 0.1 µg mL-1. The increase of gliotoxin production stopped at about 10 µg PAMP per mL.
The second step was to measure how the gliotoxin concentration varied over a 13-day period. I added 0.3, 1.0, or 5.0 µg of LPS, PG, or LTA per mL RPMI 1640 and sampled the cultures on day 1 to 5 and day 7, 9, 11, and 13. The levels of gliotoxin peaked at day 5-9 (Figure 13). Based on these results, the future cultures were sampled after seven days of incubation.
Figure 13. The influence of PG on gliotoxin production of A. fumigatus during an incubation period of 13 days. The graph illustrates how the concentration of gliotox-in varies over time, peaks between days five and eleven, and increases when PG was added to 0.3, 1, or 5 µg mL-1 to the liquid culture. The error bars indicate the stand-ard error of the reported mean (n=3).
-10
10
30
50
70
90
0.1 0.3 1 5 10 100
PG
LPS
LTA
PG, LPS and LTA
Glio
toxi
n in
crea
se (%
)
µg/ml
0
5
10
15
20
25
30
Gliot
oxin
(µM
)
Days
Control0.3 µg PG/ml1 µg PG/ml5 µg PG/ml
1 2 3 4 5 7 9 11 13

33
The third step was to show that these increased gliotoxin levels were not a matter of random variations, but a significant and measurable difference. Four equally large groups of samples (n=40 per group) were incubated with 5 µg of PG, LPS, or LTA per mL RPMI added. The gliotoxin production was correlated to the presence of PAMPs, and increased with up to 65 % compared to untreated cultures. The Mann-Whitney test clearly indicated differences in gliotoxin concentration between the four groups: PLPS=0.000035, PLTA=0.000015, and PPG=4.5*10-9 (Figure 14).
Figure 14. The difference in gliotoxin concentration between four different sets of culture (n=4*40). A Mann-Whitney test showed that the gliotoxin concentration was significantly lower (P<0.0001 for all groups, indicated with an *) in the control group (n=40) compared to groups of cultures enriched with LPS, PG or LTA at 5 µg mL-1. Error bars indicate standard error of mean.
Based on the results in Paper III there is no doubt that there are ways to af-fect the metabolism for A. fumigatus. A. fumigatus increases the secretion of gliotoxin when cultured in a medium supplemented with PAMPs. The addi-tion of LPS, LTA, and PG significantly increased the production of gliotox-in, which supports the hypothesis that there exists a receptor/signaling sys-tem in A. fumigatus that detects PAMPs and up regulates the secretion of gliotoxin. The nature and characteristics of such receptors are subject for further studies, but in mammals, the PAMPs are detected via Toll-like recep-tors and cause activation of host defense systems and immunoregulatory responses [70,71].
This interaction may be extrapolated to the biological environment in human lungs. Since molds of the genus Aspergillus are common in both indoor and outdoor air, almost all humans are daily exposed to airborne As-pergillus spp. conidia [44]. Consequently, most people are at the risk of fun-gal colonization, mainly in the respiratory tract. Individuals with e.g. chronic obstructive pulmonary disease are often colonized with Gram-negative or Gram-positive bacteria, which release LPS, LTA, and PG [72]. If the levels of these PAMPs in infected mucosal surfaces are locally comparable to the levels, which have been studied here, this will possibly increase the gliotoxin
0
10
20
30
40
50
60
70
80
90
Control LPS LTA PG
Glio
toxi
n (µ
M)
* *
*

34
production in surrounding A. fumigatus hyphae as well. As a consequence, the fungal infection will be exacerbated. Support for this theory may be found in a study of aspergillosis patients, which were treated for a co-infection of P. aeruginosa, where treatment for the bacterial infection simul-taneously diminished the presence of A. fumigatus in their lungs [73].
In a larger perspective, this study also gave insight to the inter species communication between filamentous fungi and bacteria. Applications of the described work could be useful in any industry or facility extracting metabo-lites or other products from filamentous fungi.
The potential of gliotoxin and bis(methyl)gliotoxin as biomarkers for invasive aspergillosis This study evaluated the occurrence of gliotoxin and bis(methyl)gliotoxin in BAL specimens, collected from patients being admitted at the Department of Pulmonary and Allergic Diseases at the Uppsala Akademiska Hospital. The Regional Ethical Review Board (No. 2014/157) and the Uppsala Biobank (Bio bank No. 827) approved all collection and storage of BAL samples included in this work.
Only 0.3 % of the spores in hospitals are A. fumigatus spores, but this species causes about 90 % of all systemic infections due to Aspergillus [74]. Invasive aspergillosis is a particularly problematic type of Aspergillus infec-tion, with mortalities ranging from 60 to 90 % [45,75]. The symptoms of invasive aspergillosis are not specific and several of the methods used are not conclusive individually. Physicians therefore consider a combination of clinical symptoms, radiological and microbiological findings to diagnose invasive aspergillosis (Table 6).
Table 6. Advantages and drawbacks of different methods for diagnosing invasive aspergillosis.
Type Advantages Drawbacks Reference Culture Certain evidence for the presence of A.
fumigatus. Complementary to histopat-hology.
Low sensitivity, requires biopsy
[76,77]
Histopathology Fast. Complementary ot culture-based methods.
Requires biopsy.
Galactomannan assay
Approved by FDA. Good alternative to biopsy (serum).
Not selective. Vari-able sensitivity.
[78]
Beta-D-glycan assay
Good separation proven/probable from no invasive aspergillosis. Approved by FDA.
Low specificiy for A. fumigatus.
[76,79-81]
Imaging Non-invasive. Rapid. No specificity. Radiation.
LC-MS analysis Fast. Selective. Sensitive. Undeveloped. [82,83]

35
BAL samples collected from 42 patients were analyzed for the presence of gliotoxin and bis(methyl)gliotoxin. The patients were diagnosed with differ-ent pulmonary diseases, and 21 (50 %) met the requirements of possible invasive aspergillosis (Table 7). Gliotoxin was not detected in any sample, but bis(methyl)gliotoxin was detected in 10 (24 %) at concentrations be-tween 0.32 to 18 µg mL-1. Bis(methyl)gliotoxin was detected in five (24 %) patients with possible invasive aspergillosis at concentrations between 0.32 and 8.1 µg mL-1. In the group of 21 patients without possible invasive asper-gillosis, bis(methyl)gliotoxin was detected in five (24 %) at concentrations ranging from 1.4 to 18.0 µg mL-1. The highest concentration of bis(methyl)gliotoxin was found in the sample from a pneumonia patient (No. 17). Furthermore, one more ion was observed eluting at 26 min 10 sec at m/z 357.3029 in 32 (76 %) of all 42 samples (Figure 15). The most abundant peak of this ion was at m/z 357.3029, which is close to the one of bis(methyl)gliotoxin at. m/z 357.0963, although eluting at 34 min 40 sec. Marker 1 was a doubly charged ion (M+2H)2+, indicating a molecular weight of 712.59 Da, unlike bis(methyl)gliotoxin which weighs 356.0865 Da and is singly charged (M+H)+.

36
Table 7. Distribution of bis(methyl)gliotoxin and Marker 1 in 42 BAL samples.
ID Diagnosea IA status Bis(methyl)glio-toxin (µg mL-1)b
Marker 1b
1 LTR Possible - + 2 SA - + 3 USID Possible 0.32 + 4 PN Possible - + 5 Cough Possible - - 6 SPN, AML Possible - + 7 SA - + 8 LF Possible - + 9 PN, AML Possible 8.1 -
10 AL Possible 5.4 - 11 SPN 3.4 + 12 PN - - 13 AC 2.4 + 14 SPN Possible - - 15 PN, MY Possible - + 16 SA - + 17 PN 18.0 - 18 SPN - - 19 SPN, PP Possible 2.6 + 20 DY Possible - + 21 BR, COPD - + 22 CLL Possible - + 23 Cough - + 24 LC, AC - - 25 LF Possible - + 26 HE - + 27 LC, AC - + 28 Cough 10.3 + 29 PN, PA - + 30 LF Possible - + 31 HE - + 32 LC, AC - + 33 HE, PN Possible - + 34 SA Possible - + 35 LC - - 36 Cough Possible 5.3 + 37 SPN Possible - - 38 Cough Possible - + 39 LC - + 40 LC, AC 1.4 + 41 CLL, Cough Possible - + 42 BR, Cough - -
a) Abbreviations: LTR = lung transplant recipient; SA = sarcoidosis; USID = unclear system-atic inflammatory disease; PN = pneumonia; SPN = solitary pulmonary nodule; AML = acute myeloid leukemia; LF = lung fibrosis; AL = alveolite; AC = adenocarcinoma; MY = myelo-ma; PP = pleural plaques; DY = dyspnea; BR = bronchiectasis; COPD = chronic obstructive pulmonary disease; CLL = chronic lymphocytic cancer; LC = lung cancer; HE = hemoptysis; PA = pulmonary atelectasis. b) - = not detected, + = detected.

37
Figure 15. MS chromatogram (mass window 357 to 359 m/z) and spectra of marker 1 and bis(methyl)gliotoxin. Marker 1 was an unidentified compound detected in 76 % of the BAL sample extracts. The retention time and MS data of these two sub-stances show that they are two different substances, an observation that a non-chromatographic based analysis might not have provided.
Marker 1 was detected in 76 % of the samples, spread among patients both with and without possible invasive aspergillosis. The biological origin and complete structure elucidation and origin of this marker 1 were not investi-gated further, as the amount available was not sufficient for e.g. NMR analy-sis.
This article presented a fast, sensitive, and accurate method to extract and analyze gliotoxin and bis(methyl)gliotoxin as well as other metabolites in human BAL samples. The method was optimized to assist and improve the diagnosis of invasive aspergillosis, by determining the levels of gliotoxin and bis(methyl)gliotoxin in BAL specimens. This study included analysis of BAL samples from 42 patients with various pulmonary diseases, whereof 21 with possible invasive aspergillosis.
The results do not support the use of the metabolites gliotoxin and bis(methyl)gliotoxin as biomarkers for diagnosing invasive aspergillosis. It is more likely that the use of these metabolites may give rise to false nega-tives or false positive test results, since gliotoxin was not detected in any of these samples, and bis(methyl)gliotoxin was found both in patients diag-nosed with possible invasive aspergillosis and those without. The highest concentration of bis(methyl)gliotoxin was found in a patient not diagnosed with invasive aspergillosis. The UPLC-MS equipment was a suitable choice in this study, since I could easily separate an unknown substance, marker 1, at m/z 357.3029, from bis(methyl)gliotoxin at m/z 357.0963, eluting more than eight minutes from each other. This separation would not be as easy (depending on accuracy) in an experiment lacking a chromatographic sys-tem, such as matrix associated laser desorption/ionization MS analyzers or direct-injection MS applications.
m/z357 358 359
Marker 1 Bis(methyl)gliotoxin
100 20 30 40 50 min
LC-MS chromatogram
m/z357 358 359
N N
OHH
O
O
Me
OH
S
S
Me
Me

38
The higher reactivity of gliotoxin could explain why it was not detected in any sample in this study. Its reactive ability makes it able to penetrate into, or attach to surrounding tissue in the lung, and thus not extractable from the lung tissues during bronchoscopy [55]. If the levels of gliotoxin were too low to be detected with the state-of-the-art equipment used here, it is certain-ly valid to question the value of using it as a marker metabolite for invasive aspergillosis.
Other groups have evaluated the potential of gliotoxin and bis(methyl)gliotoxin in serum as marker metabolites in invasive aspergillosis patients, and only in a few BAL samples [83,84]. Cerqueira et al. studied 30 serum sample and discovered gliotoxin in 8 of 30 samples at levels up to 138.5 ng mL-1 (0.424 µM). Domingo et al. extracted 32 serum samples and 2 BAL samples and detected bis(methyl)gliotoxin in 10 (8 serum and 2 BAL) samples up to 49.84 µg mL-1, and gliotoxin in 2 (1 serum, 1 BAL) samples up to 0.25 µg mL-1. These studies could not establish a reliable correlation between the levels of gliotoxin and bis(methyl)gliotoxin in serum or BAL and the risk of invasive aspergillosis.
Chemical analysis of BAL specimens with UPLC-MS is sensitive, accu-rate, selective, and fast – abilities that other well used diagnostic methods do not completely possess. However, an improvement of invasive aspergillosis diagnosis based on the detection of gliotoxin and bis(methyl)gliotoxin in BAL continues to be questionable. This study illustrates that this chemical analysis is not yet useful in terms of diagnosing invasive pulmonary asper-gillosis. Gliotoxin and bis(methyl)gliotoxin should be studied in a larger and preferably more diverse group of patients before the benefit of these metabo-lites could be evaluated in invasive aspergillosis diagnostics. I suggest that future attempts of chemical analysis of body fluids include a larger number of BAL samples from individuals with proven invasive aspergillosis, or fo-cus on serum and blood samples, which, so far, seem to be a more successful approach.

39
Concluding remarks
The current work involved analyses of bioactive secondary metabolites iso-lated from two fungal strains, A. fumigatus and P. nalgiovense, collected from extreme environments. The effects of different cultivation procedures on fungal metabolism were investigated, along with the scope for using the fungal metabolites gliotoxin and bis(methyl)gliotoxin as biomarkers for di-agnosing invasive aspergillosis. The key outcomes of these studies were:
• The development of a method for isolating filamentous fungi from extreme environments and for extracting and isolating antimicro-bial metabolites produced by such filamentous fungi (I and II).
• The isolation of the antibiotic metabolite gliotoxin from A. fu-migatus cultures (I).
• The discovery that P. nalgiovense is able to produce the antifungal metabolite amphotericin B (II).
• The discovery of a new metabolic response in A. fumigatus in-duced by exposure to bacterial cell wall components and mem-brane-associated molecules (III).
• An assessment of the scope for improving the diagnosis of inva-sive pulmonary aspergillosis by determining the levels of the A. fumigatus metabolites gliotoxin and bis(methyl)gliotoxin in clini-cal bronchoalveolar lavage samples (IV).
In Paper I, a strain of A. fumigatus was isolated from a riverbed down-stream a wastewater treatment plant. This strain secreted the highly antibi-otic diketopiperazine gliotoxin. Gliotoxin was first discovered in the 1950s, and has displayed toxicity against mammalian cells, and therefore not been considered a drug candidate. Paper I is the first study presenting a bioassay guided screening of bioactive compounds from filamentous fungi isolated from heavily antibiotic-contaminated environments. Both methodology and results from this work led to new ideas and experiments presented in the following three studies.
Paper II describes the isolation of a P. nalgiovense strain from a soil sample taken from a penguin’s nest on Antarctica. The hostile environment of this continent makes life extremely difficult for all organisms, and few studies on filamentous fungi from Antarctica and their metabolites have been reported. This fungal strain secreted amphotericin B, an antifungal substance

40
discovered in the 1950s but never previously isolated from any organism other than S. nodosus. The extraction yield of amphotericin B was noticeably higher when the isolated strain was cultivated on a solid medium compared to a liquid medium.
In Paper III, it was discovered that the secretion of gliotoxin from A. fu-migatus could be increased (by up to 65 %) by supplementing the liquid growth medium with pathogen-associated molecular patterns originating from Gram-negative and Gram-positive bacterial cell walls and membranes. This kind of elicitation has been studied in mammals, but its effect on fungal metabolism has been less extensively explored. These successful elicitation experiments made me conclude that A. fumigatus had an unknown signaling system for detecting bacterial presence. In this particular case, it is of rele-vance also for clinical treatment as aspergillosis is usually accompanied by bacterial co-infections. Rather than the predominant view that fungal infec-tion is clearing a path for subsequent bacterial co-infections, these results explained how the presence of bacteria could elevate gliotoxin production in A. fumigatus, which, in turn, makes the otherwise harmless fungus invasive.
In Paper IV, the scope for exploiting the secretion of gliotoxin and bis(methyl)gliotoxin by A. fumigatus to diagnose invasive pulmonary asper-gillosis was explored. Invasive aspergillosis is sometimes difficult to diag-nose accurately and promptly. Other groups have proposed that these metab-olites could be used for diagnosis, but our studies on BAL samples suggest that this is unlikely. The results presented in Paper IV suggest that future MS-based diagnostic methods should incorporate a chromatographic step to ensure adequate separation of the extracted metabolites.
Finding new bioactive compounds is the goal for most researchers in the field of pharmacognosy. For a natural product scientist, Nature is a laborato-ry with a vast number of micro- and macro-scale environmental niches, from which to collect, extract, and identify new bioactive compounds. When screening for bioactive compounds in nature, there are differences between cultivation, extraction, and analysis of filamentous fungi compared to analy-sis of plants and living organisms. In studies of microorganisms, one sample the size of a Falcon tube can contain an uncountable number of species of bacteria and filamentous fungi. Isolation, cultivation and analysis of all spe-cies in such a sample takes years, and it is impossible to predict which or-ganisms and metabolites you end up studying. The same diversity is not encountered when working with larger plants or organisms, since such stud-ies usually intend to focus on one or a few species at the time.
For sure, both approaches require samples from nature on places that may be difficult to attain. The collection of plants may be limited to a certain time of the year or a specific area. Collection sites for filamentous fungi may also be remote and cumbersome to reach. In Paper II, for example, a soil sample was obtained from a penguin’s nest on Antarctica. However, once the fila-mentous fungi are isolated, they can repeatedly be cultivated and extracted in

41
the laboratory within only days or weeks. Cultivation can be varied easily and optimized depending on the metabolism and growth of the organism. Such fast, flexible, and repeatable analysis cannot be done as easily in stud-ies of plants and other organisms.
Nonetheless, plant material collected from nature has grown “naturally”, in its preferred and unique environment. This implies that its metabolism is correlated to its interaction with surrounding (and competing) organisms. Consequently, secondary metabolites have likely been secreted for a reason, which usually is desired from a pharmacological perspective. For microor-ganisms cultivated in the laboratory, the same thing cannot be said. An iso-lated strain of a filamentous fungus is in several ways far from its natural habitat. Temperature, nutrients, light conditions, pH, temperature, water and oxygen availability, surrounding organisms, etc. are different from the natu-ral environment. For this reason, some preparation and adjustment of the cultivation media should be taken into consideration in the search for bioac-tive compounds from filamentous fungi, as presented in Papers II and III.
Filamentous fungi and other organisms depend on the production of bio-active compounds to survive predators or competing organisms. They cannot rely on nutrition depots or cell-specialization, and therefore benefit by hav-ing metabolic pathways that are downregulated under most conditions but activated when necessary for defense against competing organisms. Conse-quently, it is likely that filamentous fungi do not secrete their full spectrum of antimicrobial agents when cultivated under standard laboratory growth conditions. The exploitation of elicitation mechanisms that either activate dormant genes or increase the secretion of known metabolites is useful for exploring fungal metabolism, and such mechanisms have considerable po-tential in the identification and production of potential drugs.
The interaction of microorganisms is an exciting field of research, alt-hough the microbial natural environment is difficult to completely mimic in a laboratory. Studies of co-cultivation of filamentous fungi, plants, and bac-teria have been done but the knowledge about how these organisms interact with each other when co-cultured is rather limited [85-87]. This reflects the problems and questions that arise when studying co-cultures. In the experi-ments with filamentous fungi and bacterial cell wall material, I had to put a substantial effort into optimizing culturing conditions, and to demonstrate repeatability.
Future perspectives The lowest hanging fruits on the tree of antimicrobial metabolites have al-ready been collected. How can today’s researchers climb higher? The devel-opment of genome mining [88] and culture-independent screening methods [6] are two exciting scientific branches, which expand the borders of natural

42
product research. Genome mining aims to display genetic information such as gene clusters that code for bioactive compounds not expressed during cultivation. The use of culture-independent screening tools, such as diffusion culture chambers, has led to the recent discovery of texiobactin, which repre-sents a new class of antibacterial compounds. If these two methodologies would lead to the discovery and cultivation of only a fraction of all unknown microbial species and their secondary metabolites, much would be gained.
Co-cultivation of bacteria and filamentous fungi is another alluring tech-nique. Such experiments would give new insights in how filamentous fungi deflect and kill bacterial species, which would be a promising start for dis-covery of new antibiotics. However, an experiment that includes several living organisms, will not be successful unless the medium, growth condi-tions, and cultivation time are optimized or at least investigated. Considering how many variables these factors include, this might be challenging. Fur-thermore, if co-culturing would be a successful approach to induce metabolic responses and wake up dormant biosynthetic pathways, the biological origin of the secreted metabolites must somehow be proved. Nonetheless, once all these issues have been addressed, many, if not all, known species of filamen-tous fungi and other microorganisms would be of interest to cultivate and study yet again.
The exploration of both new and extreme environments should also be continued, regardless of cultivation method. High environmental concentra-tions of antibiotics are likely to be found in locations other than that just downstream of the particular wastewater treatment plant introduced in Paper I. Hopefully, additional studies will be conducted in other places where an-thropogenic activity has created unusual conditions or selective pressures, especially with culture-independent methods.
However, if terrestrial microorganisms cannot meet the urgent need of se-creting new antibiotics, it would be logical to focus more on marine organ-isms. The oceans are the biggest ecosystems on the planet and still largely uncharted with respect to bioactive compounds [89,90]. The development within marine natural product research during the last twenty years has re-sulted in its recognition as a sub category within natural product chemistry. While both traditional and modern cultivation techniques have enabled the discovery of various novel antibiotic compounds [6,7], my expectation is that oceans, lakes and other marine environments have high potential to har-bor easily accessible caches of bioactive substances [91].
I hope that the interdisciplinary collaborations between pharmacists, chemists, microbiologists, and engineers will fully explore these caches and unveil their bounty, leading the crossbreeding of natural product chemistry, bioinformatics, molecular biology, and genetics towards a deeper under-standing of microbial metabolism [2,88,92,93]. If the ongoing development of modern chemistry and new cultivation techniques and tools is applied in

43
natural product research, the progress with isolation of antibiotics from ter-restrial and marine microorganisms will have a bright future.
In conclusion, filamentous fungi have a broad and far-reaching capacity to reveal new bioactive metabolites. The results I have presented will be useful in the ongoing search for new antibiotics and the development of di-agnostic techniques of invasive aspergillosis. In the emerging post-antibiotic era, it will be essential to renew the focus on discovery and development of new antibiotics.

44
Summary in Swedish
Trådsvampar utsöndrar substanser, s.k. sekundära metaboliter, för att på-verka sin omgivning. Under det senaste 90 åren har trådsvampar och deras sekundära metaboliter nyttjats bl.a. inom läkemedelsindustrin för utveckling av antibiotika. Ett exempel är substansen penicillin, vilken isolerades från trådsvampen Penicillium rubens 1929. Denna upptäckt är en av de största i medicinsk historia och har räddat livet på ett stort antal människor. Många sjukdomsalstrande bakteriestammar har dock utvecklat resistens mot flera eller alla typer av tillgängliga antibiotika och dessvärre har läkemedelsföre-tagen satsat mindre resurser på utveckling av antibiotika de senaste två de-cennierna. En av orsakerna är att sjukdomsalstrande bakterier utvecklar och sprider resistenta gener snabbare än vad nya läkemedel kan upptäckas. Trots det så måste letandet efter ny antibiotika prioriteras i framtiden. En värld utan verksam antibiotika är ett stort kliv bakåt för den globala sjukvården och för mänskligheten, helt enkelt ett mardrömsscenario.
Mål och omfattning Målet med denna avhandling var att analysera sekundära metaboliter utsönd-rade av trådsvampar. Följande mål sattes upp:
• Isolera och studera trådsvampar från jordprover hämtade från ex-trema miljöer (Delarbete I och II).
• Odla de isolerade trådsvamparna från jordproverna, samt extra-hera och identifiera deras bioaktiva sekundära metaboliter (Delar-bete I och II).
• Undersöka hur olika odlingsmetoder påverkar svamparnas meta-bolism (Delarbete II och III).
• Kartlägga förekomsten av metaboliter härstammande från A. fumigatus i bronksköljvätska från patienter i syfte att förbättra diagnostiken av invasiv aspergillos (Delarbete IV).
Avhandlingen syftar till att sätta samman de fyra delarbetena i ett större sammanhang, samt att argumentera för att fortsätta att studera trådsvampar och deras sekundära metaboliter.

45
Isolering av gliotoxin från A. fumigatus Jordbanksprover från en flod nedströms ett reningsverk i Hyderabad, Indien, analyserades i Delarbete I. Flodvattnet innehöll ovanligt mycket antibiotika – bl.a. upp till 31 mg L-1 av ciprofloxacin. En sådan hög koncentration trig-gar bakteriers resistensutveckling och jag antog att även andra organismer (t.ex. trådsvampar) skulle utveckla nya metaboliter för att konkurrera med dessa bakterier. Vi isolerade 61 arter av trådsvampar från detta jordbanks-prov och flera av dem utsöndrade substanser med aktivitet mot flera resi-stenta bakteriestammar. Den A. fumigatus stammen som hade starkast aktivi-tet odlades i större skala och jag extraherade och identifierade den sekundära metaboliten gliotoxin som hade antimikrobiell aktivitet.
Detta delarbete byggde upp en arbetsmetod för att analysera extrema mil-jöer i sökandet efter antibiotiska substanser från trådsvampar. Metoderna och experimenten i detta delarbete utgjorde grunden för det preparativa och ana-lytiska arbetet i alla andra delarbeten.
Isolering av amphotericin B från P. nalgiovense Levnadsvillkoren för mikroorganismer på Antarktis är unika och denna miljö kan ha selekterat för utvecklingen av nya bioaktiva sekundära metaboliter hos trådsvampar. Studier av dessa organismer från Antarktis är relativt be-svärliga att genomföra och få har presenterats hittills. Från ett jordprov från ett pingvinbo på ön Paulete i Antarktis (Delarbete II) isolerade vi trådsvam-pen P. nalgiovense, vars extrakt hade hämmande effekt på andra trådsvam-par. Trådsvampen utsöndrade substansen amphotericin B, en sekundär meta-bolit som aldrig tidigare har isolerats från denna trådsvamp. Amphotericin B upptäcktes på 1950-talet som en sekundär metabolit producerad av bakterien Streptomyces nodosus och används idag i behandlingen av svampinfektioner.
Jag kan se tre förklaringar till att just denna stam av P. nalgiovense kan producera amphotericin B. Det kan antingen bero på att generna som kodar för produktionen av amphotericin B har överförts från S. nodosus (eller nå-gon annan organism) till P. nalgiovense, vilken använder den för att försvara sig mot andra svampar. Den kan även ha utvecklat förmågan att producera amphotericin B via en annan biosyntesväg. Kanske är det mest troligt att trådsvampen har producerat amphotericin B sedan den upptäcktes på 1920-talet, men ingen har noterat det tidigare.
Induktion av gliotoxinproduktion hos A. fumigatus I delarbete III undersökte jag om det gick att trigga en stam av A. fumigatus till att ändra sin metabolism med ändrade odlingsförutsättningar. För att

46
simulera närvaron av bakterier, tillsatte vi vanligt förekommande cellväggs- och cellmembranselement från bakterier när vi odlade trådsvampen. Resulta-tet blev att trådsvampen utsöndrade upp till 65 % mer av substansen glio-toxin. Detta samband kan innebära att det finns ett receptorsystem hos svampen som reagerar på närvaron av bakterier. Majoriteten av A. fumigatus receptorer är ännu inte identifierade och effekten av bakterierelaterade mole-kyler på svampar och deras metabolism har aldrig studerats på detta sätt tidigare.
Dessa resultat kan vara till nytta för industrier och forskningsenheter som odlar trådsvampar och isolerar deras metaboliter. De ger också en insikt i hur trådsvampar och bakterier interagerar med varandra.
Gliotoxin och bis(metyl)gliotoxin som biomarkörer för invasiv aspergillos Jag analyserade bronksköljvätska från patienter på Lung- och Allergiavdel-ningen på Uppsala Akademiska Sjukhus i syfte att förbättra diagnostiken av den potentiellt dödliga lungsjukdomen invasiv aspergillos (Delarbete IV). Syftet var att utreda om metaboliterna gliotoxin och bis(metyl)gliotoxin, vilka härstammar från A. fumigatus, kunde användas som biomarkörer för invasiv aspergillos. Diagnostiken av denna lungsjukdom är i dagsläget fullt möjligt, men är i vissa fall för osäker eller för sen. De 42 patienterna hade olika lungrelaterade sjukdomar och 21 st. (50 %) uppfyllde kraven för att ha möjlig invasiv aspergillos.
Analyserna visade att ingen av dessa substanser verkar vara lämpliga markörer för diagnostik av invasiv aspergillos. Gliotoxin fanns inte i några detekterbara mängder i något av proverna och bis(metyl)gliotoxin påvisades i prover från patienter både med och utan möjlig invasiv aspergillos. Andra studier av blod- och serumprover har visat sig vara mer framgångsrika än denna i att relatera koncentrationer av gliotoxin och bis(metyl)gliotoxin med invasiv aspergillos. Innan denna diagnostiska metod av bronksköljvätska ska bli användbar, behövs det nya studier på en större grupp patienter, helst med bevisad invasiv aspergillos.
Framtida studier Att upptäcka och isolera nya substanser med effekt mot kliniskt relevanta mikroorganismer är utmanande men nödvändigt. Odling och extraktion av trådsvampar har gjorts i snart hundra år men dessvärre mynnar alltför få försök ut i upptäckter av nya antibiotiska substanser. Detta återspeglas i de ökade problemen med multi- och omniresistenta bakterier som drabbar

47
mänskligheten världen över idag. Antibiotiska substanser från de trådsvam-par och mikroorganismer som var enklast att odla har redan exploaterats. Frukterna som hängde lägst i antibiotikaträdet är plockade och uppätna. Hur ska morgondagens forskare nå till de högre grenarna? Det finns många ar-gument för att fortsätta studera trådsvampar och deras metaboliter:
• Mer än 90 % av alla trådsvampsarter beräknas vara oupptäckta,
vilket gör dem till en outtömd källa av metaboliter [25]. • Förbättrade odlings- och analystekniker möjliggör upptäckt och
kultivering av okända mikroorganismer och deras metaboliter [6,93].
• Det finns många outforskade, extrema eller exceptionella provtag-ningsställen att leta efter antibiotika utsöndrade av trådsvampar i. Till exempel är stora delar av haven ännu outforskade [92].
Metoder som överbryggar problemet med okultiverbara mikroorganismer är ett stort steg framåt inom detta forskningsområde [6]. Ny isolering- och kul-tiveringsutrustning möjliggör upptäckande av nya organismer och deras me-taboliter. Dessutom har s.k. ”genome mining” pekats ut som ett mycket po-tentiellt forskningsområde. Denna metod baseras på den ökade tillgänglig-heten för att analysera mikrobers genom, och därmed göra kvalificerade gissningar om vilka naturprodukter som de har kapacitet att producera [88]. Möjligheten att analysera gener som vanligtvis inte används av mikroorgan-ismer under traditionell kultivering kan leda till en revitalisering av natur-produktskemi. Dessa två metoder är kapabla att öppna upp skattkistor av antibiotika och andra bioaktiva metaboliter som ligger gömda i marken och i haven. Även om bara några procent av alla okända trådsvampar och bakte-rier kan börja analyseras så skulle mycket vara vunnet.
Samkultivering av levande trådsvampar och bakterier är ett annat område som ännu är i utvecklingsstadiet. Att efterlikna trådsvamparnas naturliga miljö när man kultiverar dem är en stor utmaning. Ett steg i den riktningen är att samodla levande trådsvampar och bakterier och studera hur de interage-rar. Det skulle ge en inblick i vilka metaboliter trådsvampar utsöndrar i sin vanliga miljö för att bekämpa bakterier. Dock måste sådana metoder optime-ras så att alla samodlade organismer kan växa i mediet och dessutom säker-ställa vilken metabolit som utsöndrats av vilken organism. Här har forsk-ningen knappt lämnat startgroparna [86,87].
Om detta ändå inte skulle inducera upptäckten av ny antibiotika så finns det många miljöer kvar att studera. Jordens yta består av 70 % vatten och många havsbottnar är helt outforskade med avseende på marina trådsvampar och deras bioaktiva substanser, speciellt på djupare vatten. Även om tråd-svampar isolerade från jordprover hittills har studerats i större utsträckning än de isolerade från marina miljöer, så har de sistnämnda fått mer och mer uppmärksamhet de senaste årtiondena [91]. Tack vare utvecklingen av dyk-

48
ning, undervattensfarkoster, samt biologiska och kemiska analysmetoder under de senaste 50 åren, har jordens största ekosystem aldrig varit mer till-gängligt än nu.
Denna avhandling visar att trådsvampar är en källa till kända och okända metaboliter, och deras metabolism går att påverka med ändrade odlingsbe-tingelser. Mina resultat kan vägleda sökandet efter nya bioaktiva substanser, användas inom diagnosticering av invasiv aspergillos samt för att förstå kommunikationen mellan trådsvampar och dess omgivning.

49
Acknowledgements
This work was carried out at the Division of Pharmacognosy, Department of Medicinal Chemistry, Faculty of Pharmacy, Uppsala University. I want to reach out and show my sincere gratitude to all of you who have made this period of my life outstanding and very special. I thank myself for having the courage, endurance and brains to complete this PhD project. Supervisors Prof Lars Bohlin, for leading and guiding this project from start to goal. Prof Björn Olsen, for your visionary mind and clear leadership. Prof Ulf Göransson, for being a truly committed scientist, catalyzing my development as a scientist, and a good friend (too bad I was a vegetarian when you served that wild boar). Dr Erja Chryssanthou, for your critical and constructive assistance, which elevated my scientific output several levels. Colleagues (and friends) Dr Adam Strömstedt, for fruitful cooperation with the bacteria lab and being a cornucopia of “jokes” of both high and low quality. PhD student Erik Ja-cobsson, for sharing the q-tof maintenance work load, sharing office with me i.e. dealing with my temporary rage attacks (Jag blir galen! Jäkla skräp!), my noise issues and constant blabbing. PhD student Camilla Eriksson for being a fantastic friend and always helping out. PhD student Sung-kyu Park for being part of this trip from day one. And astonishing rhetorical skills in Mafia. PhD student Delgerbat Boldbaatar for always being kind and caring. And for being an excellent bad guy in Mafia. PhD student Sohaib Malik for the much-needed text editing of the thesis. Prof Anders Backlund, for being protective, honest and kind. Prof Joakim DG Larsson and Prof Jan Sjölin for significant scientific contribution to Pa-

50
pers I and III, respectively. Dr Hesham El-Seedi, for the NMR-introduction, Egyptian handshake and help with keeping dreams alive. Dr Anita Persson, who led me onto the right path without knowing it, and for the nice sofa (the cats love it too). And for the bottle of whiskey – it came in very handy! Dr Robert Burman, for the introduction to the q-tof (I hate it and love it). Former and current colleagues at the division: Maj, Kerstin (tack för danserna), Paco, Cessie, Christina, Muaaz, Elisabet, Astrid, Taj, Javed (when is the next Pharmacognosy cricket game?), Sunithi, Sonny, Nastiia, Teshome, Mariam, Josefine, Jenny, Eleonora (keep it simple, stupid!), An-tonella, Vanessa, Stig, Natalie, Debashish, Chinos, Rachelle, Lotta, Momo, Blazej, Aidia, Eman, Irfan, Usama, Hanna W på Karolinska, Erik H, Frans, Siv, Karam, Åke (lycka till på nya stället), Patrik M, plus everybody at the division of Analytical Pharmaceutical Chemistry: professors Curt Pettersson and Douglas Westerlund, Axel, Alexander, Anna, Annika, Alfred, Albert, Annelie, Ida, Jacob, Micke, Ylva, Victoria, Olle and Jafar. Friends and family My twin soul Dr Anders Herrmann, for being a superb friend and helper when things started to get difficult. When is our next Åland trip? My best flat mate ever Dr Anna Koptina, for always being there – I would not have come this far without your support. Вся́кому о́вощу своё вре́мя. Anna-Lena Hedman for all the fun in good times and the support in bad times. My gaming-friend #1 Alle Chen for the extended weekends in Liljeholmen with Agricola, Civ 4, good food and conversation about essential things in life (pizza, gaming and chicks). My gaming crew #1 in Uppsala: Alle Kini, Mattias and Johan S. The Agrico-la weekends kept me floating when all else dragged me down. The mendi-cant and the milking stool rules! My band Hybris – the most important band in the world - with Alle Kini, Johan A (min överman), Niclas, Erik, Mackan, Guitar-Magnus (min favorit-basist), Marit, Emma, Anna Zerpe med Z, saxofon-Johan, Mitsurugi-Miron, and Helena. And Lia for delivering much needed coffee and Greek honey cakes and sandwiches, thus making the Sunday mornings the best time of the week. Our fantastic concerts and mostly good-spirited rehearsals will always be on my mind. Becky Reid, for rapid proof reading and the guided tour of London. Marika Reis, for being a great support and the nice weekends in Malmö. Dr Mattis Fondell, to whom I should have listened more often.

51
Also Veronica, Daniel, Jonna, Fredrik G, Anna H, Valeria, Johan E, Johan V, Lennart, Inger, Julia, kurator-Linda på Wärmlands nation, hela Rolfberga IK, hela Samba Sao Miguel, Jennifer, Lily, David, Shawn & Josephine, Melly at Scuba Junkie in Kota Kinabalu, Brian S for good diving company, Suzanne and Inge for accomodation and fun in the Netherlands, and all other friends for being part of this journey. My two spoiled and nutty cats Luke and Leya, for always being around (sleeping) and looking cute or very macho. Miss you! My sisters Fredrika and Cecilia with families (Johan, Nils, Theodor, Niclas, Victor, Elin och lill-Bosse) for nice family dinners and bbq:s. My dear parents Anneli and Bosse for giving me all the necessary lessons early in life. Cooking is chemistry, and anyone can cook. “Alla saker står inte överst och längst fram!”

52
References
1. Newman DJ, Cragg GM (2012) Natural Products As Sources of New Drugs over the 30 Years from 1981 to 2010. J Nat Prod 75: 311-335.
2. Bohlin L, Goransson U, Alsmark C, Weden C, Backlund A (2010) Natural products in modern life science. Phytochem Rev 9: 279-301.
3. Cragg GM, Newman DJ (2013) Natural products: a continuing source of novel drug leads. Biochim Biophys Acta 1830: 3670-3695.
4. Rosen J, Gottfries J, Muresan S, Backlund A, Oprea TI (2009) Novel chemical space exploration via natural products. J Med Chem 52: 1953-1962.
5. El-Elimat T, Zhang X, Jarjoura D, Moy FJ, Orjala J, et al. (2012) Chemical Diversity of Metabolites from Fungi, Cyanobacteria, and Plants Relative to FDA-Approved Anticancer Agents. ACS Med Chem Lett 3: 645-649.
6. Ling LL, Schneider T, Peoples AJ, Spoering AL, Engels I, et al. (2015) A new antibiotic kills pathogens without detectable resistance. Nature 517: 455-459.
7. Jang KH, Nam SJ, Locke JB, Kauffman CA, Beatty DS, et al. (2013) Anthracimycin, a potent anthrax antibiotic from a marine-derived actinomycete. Angew Chem Int Ed Engl 52: 7822-7824.
8. Lapworth DJ, Baran N, Stuart ME, Ward RS (2012) Emerging organic contaminants in groundwater: A review of sources, fate and occurrence. Environ Pollut 163: 287-303.
9. Andersson DI, Hughes D (2011) Persistence of antibiotic resistance in bacterial populations. FEMS Microbiol Rev 35: 901-911.
10. Larsson DG, de Pedro C, Paxeus N (2007) Effluent from drug manufactures contains extremely high levels of pharmaceuticals. J Hazard Mater 148: 751-755.
11. Kristiansson E, Fick J, Janzon A, Grabic R, Rutgersson C, et al. (2011) Pyrosequencing of antibiotic-contaminated river sediments reveals high levels of resistance and gene transfer elements. PLoS One 6: e17038.
12. Cowan DA, Makhalanyane TP, Dennis PG, Hopkins DW (2014) Microbial ecology and biogeochemistry of continental Antarctic soils. Front Microbiol 5.
13. Fleming A (1929) On the antibacterial of cultures of a Penicillium with special reference to their use in isolation of B. influenzae. Brit J of Exp Pathol 10: 226-236.
14. Bo G (2000) Giuseppe Brotzu and the discovery of cephalosporins. Clin Microbiol Infect 6 Suppl 3: 6-9.
15. Oxford AE, Raistrick H, Simonart P (1939) Studies in the biochemistry of micro-organisms: Griseofulvin, C(17)H(17)O(6)Cl, a metabolic product of Penicillium griseofulvum Dierckx. Biochem J 33: 240-248.
16. Borel JF, Feurer C, Gubler HU, Stahelin H (1976) Biological effects of cyclosporin A: a new antilymphocytic agent. Agents Actions 6: 468-475.
17. Kluepfel D, Bagli J, Baker H, Charest MP, Kudelski A (1972) Myriocin, a new antifungal antibiotic from Myriococcum albomyces. J Antibiot (Tokyo) 25: 109-115.

53
18. Endo A (1992) The Discovery and Development of Hmg-Coa Reductase Inhibitors. J Lipid Res 33: 1569-1582.
19. Tamura G, Ando K, Suzuki S, Takatsuki A, Arima K (1968) Antiviral activity of brefeldin A and verrucarin A. J Antibiot (Tokyo) 21: 160-161.
20. Tomasz A (1979) The mechanism of the irreversible antimicrobial effects of penicillins: how the beta-lactam antibiotics kill and lyse bacteria. Annu Rev Microbiol 33: 113-137.
21. Barna JC, Williams DH (1984) The structure and mode of action of glycopeptide antibiotics of the vancomycin group. Annu Rev Microbiol 38: 339-357.
22. Drlica K, Malik M, Kerns RJ, Zhao X (2008) Quinolone-mediated bacterial death. Antimicrob Agents Chemother 52: 385-392.
23. Vakulenko SB, Mobashery S (2003) Versatility of aminoglycosides and prospects for their future. Clin Microbiol Rev 16: 430-450.
24. Luzzatto L, Apirion D, Schlessinger D (1968) Mechanism of action of streptomycin in E. coli: interruption of the ribosome cycle at the initiation of protein synthesis. Proc Natl Acad Sci U S A 60: 873-880.
25. Scheffers BR, Joppa LN, Pimm SL, Laurance WF (2012) What we know and don't know about Earth's missing biodiversity. Trends Ecol Evol 27: 501-510.
26. Torsvik V, Ovreas L (2002) Microbial diversity and function in soil: from genes to ecosystems. Curr Opin Microbiol 5: 240-245.
27. Venter JC, Remington K, Heidelberg JF, Halpern AL, Rusch D, et al. (2004) Environmental genome shotgun sequencing of the Sargasso Sea. Science 304: 66-74.
28. Shendure J, Ji H (2008) Next-generation DNA sequencing. Nat Biotechnol 26: 1135-1145.
29. Rothberg JM, Leamon JH (2008) The development and impact of 454 sequencing. Nat Biotechnol 26: 1117-1124.
30. Belnap CP, Pan C, Denef VJ, Samatova NF, Hettich RL, et al. (2011) Quantitative proteomic analyses of the response of acidophilic microbial communities to different pH conditions. ISME J 5: 1152-1161.
31. Grice CM, Bertuzzi M, Bignell EM (2013) Receptor-mediated signaling in Aspergillus fumigatus. Front Microbiol 4: 26.
32. Kawai T, Akira S (2010) The role of pattern-recognition receptors in innate immunity: update on Toll-like receptors. Nat Immunol 11: 373-384.
33. Blasius AL, Beutler B (2010) Intracellular toll-like receptors. Immunity 32: 305-315.
34. Hoffmann JA, Kafatos FC, Janeway CA, Ezekowitz RA (1999) Phylogenetic perspectives in innate immunity. Science 284: 1313-1318.
35. Walsh TJ, Anaissie EJ, Denning DW, Herbrecht R, Kontoyiannis DP, et al. (2008) Treatment of aspergillosis: clinical practice guidelines of the Infectious Diseases Society of America. Clin Infect Dis 46: 327-360.
36. Segal BH, Walsh TJ (2006) Current approaches to diagnosis and treatment of invasive aspergillosis. Am J Resp Crit Care 173: 707-717.
37. Patterson TF (2005) Advances and challenges in management of invasive mycoses. Lancet 366: 1013-1025.
38. Scharf DH, Heinekamp T, Remme N, Hortschansky P, Brakhage AA, et al. (2012) Biosynthesis and function of gliotoxin in Aspergillus fumigatus. Appl Microbiol Biotechnol 93: 467-472.
39. Pardo J, Urban C, Galvez EM, Ekert PG, Muller U, et al. (2006) The mitochondrial protein Bak is pivotal for gliotoxin-induced apoptosis and a critical host factor of Aspergillus fumigatus virulence in mice. J Cell Biol 174: 509-519.

54
40. Comera C, Andre K, Laffitte J, Collet X, Galtier P, et al. (2007) Gliotoxin from Aspergillus fumigatus affects phagocytosis and the organization of the actin cytoskeleton by distinct signalling pathways in human neutrophils. Microbes Infect 9: 47-54.
41. Arseculeratne SN, de Silva LM, Wijesundera S, Bandunatha CH (1969) Coconut as a medium for the experimental production of aflatoxin. Appl Microbiol 18: 88-94.
42. Chang PK, Skory CD, Linz JE (1992) Cloning of a gene associated with aflatoxin B1 biosynthesis in Aspergillus parasiticus. Curr Genet 21: 231-233.
43. Wiegand I, Hilpert K, Hancock RE (2008) Agar and broth dilution methods to determine the minimal inhibitory concentration (MIC) of antimicrobial substances. Nat Protoc 3: 163-175.
44. Latge JP (1999) Aspergillus fumigatus and aspergillosis. Clin Microbiol Rev 12: 310-335.
45. Balloy V, Chignard M (2009) The innate immune response to Aspergillus fumigatus. Microbes Infect 11: 919-927.
46. Bardana EJ, Jr. (1981) The clinical spectrum of aspergillosis--part 1: epidemiology, pathogenicity, infection in animals and immunology of Aspergillus. Crit Rev Clin Lab Sci 13: 21-83.
47. Araujo R, Rodrigues AG (2004) Variability of germinative potential among pathogenic species of Aspergillus. J Clin Microbiol 42: 4335-4337.
48. Askew DS (2008) Aspergillus fumigatus: virulence genes in a street-smart mold. Curr Opin Microbiol 11: 331-337.
49. Dagenais TRT, Keller NP (2009) Pathogenesis of Aspergillus fumigatus in Invasive Aspergillosis. Clin Microbiol Rev 22: 447-465.
50. Rementeria A, Lopez-Molina N, Ludwig A, Vivanco AB, Bikandi J, et al. (2005) Genes and molecules involved in Aspergillus fumigatus virulence. Rev Iberoam Micol 22: 1-23.
51. Nierman WC, Pain A, Anderson MJ, Wortman JR, Kim HS, et al. (2005) Genomic sequence of the pathogenic and allergenic filamentous fungus Aspergillus fumigatus. Nature 438: 1151-1156.
52. Stiller ET, Vandeputte J, Wachtel JL (1955) Amphotericins A and B, antifungal antibiotics produced by a streptomycete. II. The isolation and properties of the crystalline amphotericins. Antibiot Annu 3: 587-591.
53. Andersen SJ, Frisvad JC (1994) Penicillin production by Penicillium nalgiovense. Lett Appl Microbiol 19: 486-488.
54. Frisvad JC, Smedsgaard J, Larsen TO, Samson RA (2004) Mycotoxins, drugs and other extrolites produced by species in Penicillium subgenus Penicillium. Stud Mycol: 201-241.
55. Trown PW, Bilello JA (1972) Mechanism of Action of Gliotoxin - Elimination of Activity by Sulfhydryl Compounds. Antimicrob Agents Ch 2: 261-265.
56. Rodriguez PL, Carrasco L (1992) Gliotoxin - Inhibitor of Poliovirus Rna-Synthesis That Blocks the Viral-Rna Polymerase-3dpol. J Virol 66: 1971-1976.
57. Gardiner DM, Waring P, Howlett BJ (2005) The epipolythiodioxopiperazine (ETP) class of fungal toxins: distribution, mode of action, functions and biosynthesis. Microbiol 151: 1021-1032.
58. Bernardo PH, Brasch N, Chai CL, Waring P (2003) A novel redox mechanism for the glutathione-dependent reversible uptake of a fungal toxin in cells. J Biol Chem 278: 46549-46555.

55
59. Waring P, Sjaarda A, Lin QH (1995) Gliotoxin inactivates alcohol dehydrogenase by either covalent modification or free radical damage mediated by redox cycling. Biochem Pharmacol 49: 1195-1201.
60. Hurne AM, Chai CL, Waring P (2000) Inactivation of rabbit muscle creatine kinase by reversible formation of an internal disulfide bond induced by the fungal toxin gliotoxin. J Biol Chem 275: 25202-25206.
61. McNamara CM, Box S, Crawforth JM, Hickman BS, Norwood TJ, et al. (1998) Biosynthesis of amphotericin B. J Chem Soc Perk T 1: 83-87.
62. Sowinski P, Pawlak J, Borowski E, Gariboldi P (1992) 1H NMR Model Studies of Amphotericin-B - Comparison of X-Ray and NMR Stereochemical Data. Magn Reson Chem 30: 275-279.
63. Larsen TO, Breinholt J (1999) Dichlorodiaportin, diaportinol, and diaportinic acid: three novel isocoumarins from Penicillium nalgiovense. J Nat Prod 62: 1182-1184.
64. Raistrick H, Ziffer J (1951) Studies in the Biochemistry of Micro-Organisms .84. The Colouring Matters of Penicillium-Nalgiovensis-Laxa .1. Nalgiovensin and Nalgiolaxin - Isolation, Derivatives and Partial Structures. Biochem J 49: 563-574.
65. Sorensen D, Larsen TO, Christophersen C, Nielsen PH, Anthoni U (1999) Dipodazine, a diketopiperazine from Penicillium dipodomys. Phytochemistry 51: 1181-1183.
66. Adler-Moore J, Proffitt RT (2002) AmBisome: liposomal formulation, structure, mechanism of action and pre-clinical experience. J Antimicrob Chemoth 49: 21-30.
67. Baginski M, Czub J (2009) Amphotericin B and Its New Derivatives - Mode of Action. Curr Drug Metab 10: 459-469.
68. Jaramillo VD, Sukno SA, Thon MR (2015) Identification of horizontally transferred genes in the genus Colletotrichum reveals a steady tempo of bacterial to fungal gene transfer. BMC Genomics 16: 2.
69. Hall C, Brachat S, Dietrich FS (2005) Contribution of horizontal gene transfer to the evolution of Saccharomyces cerevisiae. Eukaryot Cell 4: 1102-1115.
70. Mogensen TH (2009) Pathogen recognition and inflammatory signaling in innate immune defenses. Clin Microbiol Rev 22: 240-273.
71. Janeway CA, Jr., Medzhitov R (2002) Innate immune recognition. Annu Rev Immunol 20: 197-216.
72. Kousha M, Tadi R, Soubani AO (2011) Pulmonary aspergillosis: a clinical review. Eur Respir Rev 20: 156-174.
73. Baxter CG, Rautemaa R, Jones AM, Webb AK, Bull M, et al. (2013) Intravenous antibiotics reduce the presence of Aspergillus in adult cystic fibrosis sputum. Thorax 68: 652-657.
74. Brakhage AA, Langfelder K (2002) Menacing mold: The molecular biology of Aspergillus fumigatus. Annu Rev Microbiol 56: 433-455.
75. Maschmeyer G, Haas A, Cornely OA (2007) Invasive Aspergillosis - Epidemiology, diagnosis and management in immunocompromised patients. Drugs 67: 1567-1601.
76. De Pauw B, Walsh TJ, Donnelly JP, Stevens DA, Edwards JE, et al. (2008) Revised definitions of invasive fungal disease from the European Organization for Research and Treatment of Cancer/Invasive Fungal Infections Cooperative Group and the National Institute of Allergy and Infectious Diseases Mycoses Study Group (EORTC/MSG) Consensus Group. Clin Infect Dis 46: 1813-1821.

56
77. Cuenca-Estrella M, Bassetti M, Lass-Florl C, Racil Z, Richardson M, et al. (2011) Detection and investigation of invasive mould disease. J Antimicrob Chemoth 66: i15-i24.
78. Leeflang MM, Debets-Ossenkopp YJ, Visser CE, Scholten RJ, Hooft L, et al. (2008) Galactomannan detection for invasive aspergillosis in immunocompromized patients. Cochrane Database Syst Rev: CD007394.
79. Kontoyiannis DP, Marr KA, Park BJ, Alexander BD, Anaissie EJ, et al. (2010) Prospective surveillance for invasive fungal infections in hematopoietic stem cell transplant recipients, 2001-2006: overview of the Transplant-Associated Infection Surveillance Network (TRANSNET) Database. Clin Infect Dis 50: 1091-1100.
80. Odabasi Z, Mattiuzzi G, Estey E, Kantarjian H, Saeki F, et al. (2004) Beta-D-glucan as a diagnostic adjunct for invasive fungal infections: validation, cutoff development, and performance in patients with acute myelogenous leukemia and myelodysplastic syndrome. Clin Infect Dis 39: 199-205.
81. Obayashi T, Yoshida M, Mori T, Goto H, Yasuoka A, et al. (1995) Plasma (1-->3)-beta-D-glucan measurement in diagnosis of invasive deep mycosis and fungal febrile episodes. Lancet 345: 17-20.
82. Puri A, Ahmad A, Panda BP (2010) Development of an HPTLC-based diagnostic method for invasive aspergillosis. Biomed Chromatogr 24: 887-892.
83. Domingo MP, Colmenarejo C, Martinez-Lostao L, Mullbacher A, Jarne C, et al. (2012) Bis(methyl)gliotoxin proves to be a more stable and reliable marker for invasive aspergillosis than gliotoxin and suitable for use in diagnosis. Diagn Micr Infec Dis 73: 57-64.
84. Cerqueira LB, de Francisco TM, Gasparetto JC, Campos FR, Pontarolo R (2014) Development and validation of an HPLC-MS/MS method for the early diagnosis of aspergillosis. PLoS One 9: e92851.
85. Moreno PR, van der Heijden R, Verpoorte R (1993) Effect of terpenoid precursor feeding and elicitation on formation of indole alkaloids in cell suspension cultures of Catharanthus roseus. Plant Cell Rep 12: 702-705.
86. Xu XL, Lee RTH, Fang HM, Wang YM, Li R, et al. (2008) Bacterial peptidoglycan triggers Candida albicans hyphal growth by directly activating the adenylyl cyclase Cyr1p. Cell Host Microbe 4: 28-39.
87. Flores-Sanchez IJ, Pec J, Fei J, Choi YH, Dusek J, et al. (2009) Elicitation studies in cell suspension cultures of Cannabis sativa L. J Biotechnol 143: 157-168.
88. Bachmann BO, Van Lanen SG, Baltz RH (2014) Microbial genome mining for accelerated natural products discovery: is a renaissance in the making? J Ind Microbiol Biot 41: 175-184.
89. Mayer AMS, Rodriguez AD, Taglialatela-Scafati O, Fusetani N (2013) Marine Pharmacology in 2009-2011: Marine Compounds with Antibacterial, Antidiabetic, Antifungal, Anti-Inflammatory, Antiprotozoal, Antituberculosis, and Antiviral Activities; Affecting the Immune and Nervous Systems, and other Miscellaneous Mechanisms of Action. Mar Drugs 11: 2510-2573.
90. Omarsdottir S, Einarsdottir E, Ogmundsdottir HM, Freysdottir J, Olafsdottir ES, et al. (2013) Biodiversity of benthic invertebrates and bioprospecting in Icelandic waters. Phytochem Rev 12: 517-529.
91. Overy DP, Bayman P, Kerr RG, Bills GF (2014) An assessment of natural product discovery from marine (sensu strictu) and marine-derived fungi. Mycology 5: 145-167.

57
92. Gerwick WH, Moore BS (2012) Lessons from the Past and Charting the Future of Marine Natural Products Drug and Discovery and Chemical Biology. Chem Biol 19: 1631-1631.
93. Mi Q, Pezzuto JM, Farnsworth NR, Wani MC, Kinghorn AD, et al. (2009) Use of the in vivo hollow fiber assay in natural products anticancer drug discovery. J Nat Prod 72: 573-580.

Acta Universitatis UpsaliensisDigital Comprehensive Summaries of Uppsala Dissertationsfrom the Faculty of Pharmacy 195
Editor: The Dean of the Faculty of Pharmacy
A doctoral dissertation from the Faculty of Pharmacy, UppsalaUniversity, is usually a summary of a number of papers. A fewcopies of the complete dissertation are kept at major Swedishresearch libraries, while the summary alone is distributedinternationally through the series Digital ComprehensiveSummaries of Uppsala Dissertations from the Faculty ofPharmacy. (Prior to January, 2005, the series was publishedunder the title “Comprehensive Summaries of UppsalaDissertations from the Faculty of Pharmacy”.)
Distribution: publications.uu.seurn:nbn:se:uu:diva-242611
ACTAUNIVERSITATIS
UPSALIENSISUPPSALA
2015


















