
Analysis of Response Calls to Diverse Ground Predators ...
47
Taylor University Pillars at Taylor University Master of Environmental Science esis Collection 2006 Analysis of Response Calls to Diverse Ground Predators from ree Geographically Separate Florida Scrub-Jay (Aphelocoma coerulescens) Subpopulations Sarah A. (Bales) Douglass Follow this and additional works at: hps://pillars.taylor.edu/mes Part of the Environmental Sciences Commons is esis is brought to you for free and open access by Pillars at Taylor University. It has been accepted for inclusion in Master of Environmental Science esis Collection by an authorized administrator of Pillars at Taylor University. For more information, please contact [email protected]. Recommended Citation (Bales) Douglass, Sarah A., "Analysis of Response Calls to Diverse Ground Predators from ree Geographically Separate Florida Scrub-Jay (Aphelocoma coerulescens) Subpopulations" (2006). Master of Environmental Science esis Collection. 3. hps://pillars.taylor.edu/mes/3
Transcript of Analysis of Response Calls to Diverse Ground Predators ...

Taylor UniversityPillars at Taylor University
Master of Environmental Science Thesis Collection
2006
Analysis of Response Calls to Diverse GroundPredators from Three Geographically SeparateFlorida Scrub-Jay (Aphelocoma coerulescens)SubpopulationsSarah A. (Bales) Douglass
Follow this and additional works at: https://pillars.taylor.edu/mes
Part of the Environmental Sciences Commons
This Thesis is brought to you for free and open access by Pillars at Taylor University. It has been accepted for inclusion in Master of EnvironmentalScience Thesis Collection by an authorized administrator of Pillars at Taylor University. For more information, please contact [email protected].
Recommended Citation(Bales) Douglass, Sarah A., "Analysis of Response Calls to Diverse Ground Predators from Three Geographically Separate FloridaScrub-Jay (Aphelocoma coerulescens) Subpopulations" (2006). Master of Environmental Science Thesis Collection. 3.https://pillars.taylor.edu/mes/3

ANALYSIS OF RESPONSE CALLS TO DIVERSE GROUND PREDATORS FROM
THREE GEOGRAPHICALLY SEPARATE FLORIDA SCRUB-JAY (APHELOCOMA
COERULESCENS) SUBPOPULATIONS
SARAH A. BALES1*, JAN M. REBER2, M. REBECCA BOLT3, and ROBERT T. REBER4
1Department of Earth and Environmental Science, 236 West Reade Ave, Taylor
University, Upland, Indiana 46989
2Department of Biology, 236 West Reade Ave, Taylor University, Upland, Indiana 46989
3The Dynamac Corporation, DYN-1, Kennedy Space Center, Florida 32899; and
4Department of Earth and Environmental Science, 236 West Reade Ave, Taylor
University, Upland, Indiana 46989
*Email: [email protected]

2
ABSTRACT
Dialectic variation occurs in many bird species. Different factors have been
investigated regarding dialectic variation, including cultural and genetic transmission of
songs or calls, and geographic separation. In this study, the predator alarm call of the
Florida Scrub-Jay to ground predators was recorded in three geographically separate
subpopulations across Florida—Canaveral National Seashore/Merritt Island National
Wildlife Refuge, Lyonia Preserve and Leisure Lakes/Lake June in Winter Scrub State
Park—to examine any dialectic occurrence between these three subpopulations.
Additionally, many bird species are recognized as having highly evolved predator
recognition systems, often with different calls for specific predators. The Florida Scrub-
Jay has exhibited such varied call responses to predators. Three different ground predator
treatments, an artificial snake, a live snake, and a live cat, were used to elicit alarm calls.
Responses to each predator were then compared within a subpopulation. This study
revealed that significant differences existed between subpopulations responding to the cat
and artificial snake treatment, but no significant differences existed between
subpopulations for the live snake treatment. These results indicate dialectic divergence
between subpopulations in response to some species of ground predators, but not to the
snake, with which the birds have evolved and are most familiar. Florida Scrub-Jay
responses to the three ground predators were significantly different within two
subpopulations, at Leisure Lakes/Lake June in Winter Scrub State Park and Lyonia
Preserve, but not within the Canaveral National Seashore/Merritt Island National Wildlife
Refuge subpopulation. Results from this study could be valuable to scientists, wildlife
managers, and communities striving to improve their conservation efforts.

3
Key words: predator alarm call, ground predators, Florida Scrub-Jay, dialectic
variation, geographic separation, habitat fragmentation, conservation

4
INTRODUCTION
The Florida Scrub-Jay, Aphelocoma coerulescens, was officially designated in
1987 as Threatened due to a major decline in its population, an approximately 25-50
percent decrease over the previous decade (USFWS 1999). This decline in the Florida
Scrub-Jay (FLSJ) population was attributed to significant habitat loss, fragmentation, and
degradation that resulted primarily from an increase in urbanization, agriculture, and lack
of scrub management (USFWS 1999).
Florida Scrub-Jay families consist of a monogamous pair with additional
“helpers” that are typically previous offspring, and often number between two to eight in
total family size (McGowan and Woolfenden 1989). The risk of potential predation
decreases when birds live in groups because of the increased protection sentinels add.
This sentinel system involves an individual bird that is not foraging, but rather stands
guard, exchanging this task with other birds of the same family group (McGowan and
Woolfenden 1989). Particular behaviors, such as mobbing and alarm calls, are elicited
when a potential threat is spotted by a sentinel who then initiates an alarm call.
Florida Scrub-Jays exhibit mobbing behavior through loud calling, scolding, and
visual cues, described by Francis et al. (1989), to draw attention to possible predators.
They often mob predators on the ground, but hide from raptors spotted in the air
(McGowan and Woolfenden 1989). By drawing attention to a predator their survivability
increases. This is especially important because predation is the largest contributing factor
to FLSJ mortality, particularly during nesting (Woolfenden and Fitzpatrick 1984,
Fitzpatrick et al. 1991; Bowman and Woolfenden 2001; Thorington and Bowman 2003).
Snakes are considered to be a major nest predator on both nestlings and fledglings

5
(Schaub et al. 1992; Woolfenden and Fitzpatrick 1996). Free-ranging (including feral)
cats are increasingly becoming a threat to all birds, including the FLSJ. In a report by the
Florida Fish and Wildlife Conservation Commission (2001), it is estimated that free-
ranging cats in Florida may be killing 68 million birds per year.
The decline in FLSJ populations is not only attributed to high predation risks but
also to habitat fragmentation. Habitat fragmentation often disrupts and isolates smaller
portions of a population resulting in metapopulations and eventually subpopulations. As
these subpopulations become more isolated from one another, due to urbanization or fire
suppression, the risk of extinction will increase (Woolfenden and Fitzpatrick 1991; Stith
et al. 1996; Breininger et al. 1999). Once isolated, genetic variation may occur, which
can also result in dialect formation among spatially separated species. Song dialects have
been extensively studied and numerous hypotheses posed about their adaptive
significance (MacDougall-Shackleton and MacDougall-Shackleton 2001).
Many bird species are known to have different song and/or call dialects, including
the White-crowned Sparrow (Baker and Cunningham 1985; Cunningham et al. 1987),
Orange-fronted Parakeet (Bradbury et al. 2001), Song Sparrow (Harris and Lemon 1971),
Northern Cardinal (Lemon 1966), and Bewick’s Wren (Kroodsma 1974). Conclusions
from these studies indicate that song dialects may be influenced by factors that include
cultural learning, genetic transmission, and geographic separation. Relatively few studies
have examined the acoustical and genetic differences between populations of the FLSJ.
Cultural learning has been described as non-genetic song-learning, from
generation to generation (Liaolo et al. 2001). Songs or calls can be learned over time
(cultural-evolution) and the errors that occur in this learning process may contribute to

6
song variability (Lynch and Baker 1994). Genetic mutations that arise, as well as with
population size and migration, could largely affect call diversity within a population
(Lynch and Baker 1993). The influence geographic separation has on dialectic variation
also may involve cultural and genetic transmission factors. With the lack of interaction
between neighboring populations, these cultural and genetic factors could escalate the
rate of dialectic divergence. For example, if a population became more isolated from
another population, they are less likely to share songs and genes. If one of these
populations were to learn or establish songs that became distinctly different through time,
they would no longer share similar songs and, therefore, potentially not reproduce. This
could result in further genetic isolation. In addition, different environmental pressures
may influence song transmission within populations in geographically separate areas
(Lynch and Baker 1993). Possible environmental pressures may include different
predators or habitat types, factors pertinent to this study.
Dispersal distances also may contribute to population and dialectic variation. The
Florida Scrub-Jay is a non-migratory species with intense natal philopatry, thereby
remaining near its natal territory throughout life (Stith et al. 1996; Fitzpatrick et al. 1999).
Fitzpatrick et al. (1999) found that FLSJ disperse, on average, about 1.6 km from their
natal territory. This dispersal rate is noted as being one of the most restricted among all
bird species (Stenzler and Fitzpatrick 2002). Fitzpatrick et al. (1999) also found that
FLSJ are more likely to disperse within the corridors formed by historic sand ridges
found across Florida. The xeric oak scrub ecosystem (optimal FLSJ habitat) is associated
with Florida’s ancient dune systems that parallel the coasts of Florida (Woolfenden and
Fitzpatrick 1996). These sand ridges include the Lake Wales Ridge area, and both

7
coastal (Gulf and Atlantic) areas. The xeric oak scrub ecosystem has been described by
Menges (1999) in detail along with specific habitat requirements of the FLSJ. Patches in
this ecosystem that provide optimum habitat contain: low oak-dominated species about 2
m in height or less, minimal pine canopy, and open sandy patches for caching acorns
(Fitzpatrick et al. 1991; Breininger et al. 1995; Menges 1999; Bowman and Woolfenden
2002).
The levels of threat presented to birds by predators are species specific, and their
response (behaviorally and acoustically) has evolved to recognize and address diverse
predators and their associated danger. The Florida Scrub-Jay particularly exemplifies a
social unit which responds differently to different predator-related threats. The FLSJ has
been extensively studied, largely because of its unique family structure that involves
helpers, a cooperative breeding strategy, non-migratory nature, and specific habitat
requirements (Woolfenden and Fitzpatrick 1990). Most studies have examined FLSJ life
history patterns while few have actually analyzed their vocalizations, especially predator-
elicited calls.
An early study by Barbour (1977) categorized the FLSJ vocal repertoire and
recognized differences between their response to ground and aerial predators. The FLSJ
response to a ground predator is referred to as the “predator screech scold,” while the
response to an aerial predator is noted as an “aerial predator alarm weep” (Barbour 1977).
The response to an aerial predator is not only acoustically different but behaviorally
different than to a ground predator (Barbour 1977). Other animal species, such as the
California ground squirrel and Gunnison’s prairie dog, also have different responses to
aerial and ground predators (Placer and Slobodchikoff 2000). One aspect of this present

8
study focuses on the FLSJ acoustic response to ground predators, and the term “predator
screech scold” or “predator alarm call” are used interchangeably herein.
In another study (Elowson and Hailman 1991), FLSJ predator-elicited calls to
both ground and aerial predators were classified through dichotomous sorting. In
Elowson and Hailman’s (1991) work, predator-elicited calls of the FLSJ showed complex
variation encompassing twelve different call types and subtypes in response to various
aerial and ground predators. In particular, the predator alarm calls to snakes clustered
into a ‘flat’ call type, with four subtypes. For further description of the different call
types and subtypes see Elowson and Hailman (1991).
Another study examined the “hiccup” call between three sub-regional
populations across Florida (Russel unpubl. data 2005). Hiccup calls are exclusive to
females and often made in territorial disputes, for further description see Barbour (1977).
Interestingly, for the first time, Russel (unpubl. data 2005) found dialectic variation in the
hiccup call between all three sub-regional populations.
A highly developed predator recognition system also has evolved in other bird
species such as Carolina Wrens (Morton and Shalter 1977), Black-capped Chickadees
(Apel and Ficken 1981; Baker and Becker 2002), Tufted Titmice (Waite and Grubb
1987), and Lapwings (Walters 1990). These studies revealed some bird species have
diverse responses to various predators. These responses include different call structures
related to the level of threat presented by a predator (Morton and Shalter 1977; Baker and
Becker 2002) or distinct call types to various predators (Walters 1990). Some other
examples in the animal kingdom that incorporate specific behavioral and/or acoustical
signals for predator recognition include vervet monkeys (Cheney and Seyfarth 1990;

9
Brown et al. 1992), baboons (Fischer et al. 2001), and prairie dogs (Ackers and
Slobodchikoff 1999). The potential risk of predation decreases as a social unit cooperates
to watch for and detect predators.
Due to its specific habitat requirements, limited dispersal range, and increasing
fragmentation of its optimal habitat, the FLSJ is at high risk for further population
isolation. This may lead to not only greater dialectic variation but genetic isolation
between populations and, therefore, potentially to speciation or extinction. The focus of
this study was two-fold: to investigate possible dialectic variation in the alarm call
responses between three geographically separate FLSJ subpopulations, and to determine
any significant differences in FLSJ responses to three different ground predators. This
study hopes to contribute to the understanding of call types, dialectic variances, adaptive
significance, and conservation management for the FLSJ.
METHODS
Study sites and populations.—There are five major sub-regions of the Florida
Scrub-Jay found in geographically separate locations, three of which are considered “core
populations” (Fig. 1) (Fitzpatrick et al. 1994 cited in USFWS 1999). This study focused
on three subpopulations within the core populations from June to August 2005. All three
sub-regional areas—the Atlantic Coast Subregion, Ocala Subregion, and Lake Wales
Ridge Subregion—occur along major sand ridges (Fitzpatrick et al. 1994 cited in USFWS
1999). The first study site was at Canaveral National Seashore/Merritt Island National
Wildlife Refuge (CANA/MINWR) and Kennedy Space Center (KSC), referred to as CA
hereafter. This rural study site was located in the Atlantic Coast Subregion. The second
study site was located at Leisure Lakes (LL) near Lake June in Winter Scrub State Park,

10
Lake Placid, Florida. This semi-rural study site was located in the Lake Wales Ridge
Subregion. The third study site, in the Ocala Subregion, was a 360-acre preserve called
Lyonia Preserve (LY). This preserve was located behind the Deltona Regional Library in
the city of Deltona, Florida. All three sites consisted of xeric habitats that ranged from
high quality managed oak scrub, to partially managed scrubby flatwoods, and restored
scrub habitat from sand pine and oak stands, respectively.
At each study site, six to eight families were randomly chosen and recorded.
Typically, these families were easily identified because most or all members were banded
with a US Fish and Wildlife metal band and a color band pattern issued by previous
researchers. This allows for identifying which family particular birds belong to. The
Florida Scrub-Jays were habituated to human observers at all sites.
Experimental procedure and equipment.—One artificial and two live predators
were used to elicit predator alarm calls. An Eastern Corn Snake, Elaphe guttata, and a
domestic cat, Felis catus, were used for the two live predator stimuli. An artificial blow-
up snake was used as the third stimulus. The model snake mimicked an Eastern Indigo,
Drymarchon couperi, in size and coloration. It was chosen because it is a natural snake
predator of the FLSJ. All trials were conducted in the mornings from 0730 to 1200.
Under no circumstances were the live animals involved in this study harmed in any way,
and at no time did they pose any harm to the FLSJ.
Three of the six to eight designated families were randomly chosen to be recorded
the first of a two-day period, and another three were recorded on the next day. This
schedule was repeated after a 24 h wait period. If all three families were not recorded on
a given day, a recording of the missed family was attempted on the third day. In some

11
cases, families were recorded over a three day period with a 24 hour wait period followed
by either two or three more days of recording. This procedure was repeated for each
treatment at each site. The exception occurred at Leisure Lakes where it was necessary to
record two families each day over a consecutive four day period. The initial failure of the
randomly selected families to appear on the first day was most likely due to the onset of
the acorn season (August). These families were possibly actively foraging and caching
acorns (R. Bowman, pers. comm.), which are an important food source and critical to
FLSJ survival (Menges 1999).
The artificial snake was the first stimulus exhibited to FLSJ families, followed by
the live Eastern Corn Snake and, finally, a house cat. The artificial snake was placed on a
saw palmetto or oak scrub species with a string attached to produce movement. The trial
began when at least one adult FLSJ was present. The trial ended when the birds ceased
scolding and apparently lost interest or simply flew away. Trial duration resulted in an
average of 3 minutes of recording time.
The live Eastern Corn Snake was placed in a Plexiglas and wood-structured cage
9 ¾ x 10 x 32 inches (w x h x l) that was set on a small foldable table and placed within
the scrub but still conspicuous to the birds. The trial ended when the birds flew away
after scolding, or when the snake was removed for its own well-being (heat stress), but
usually after about 3 minutes of recording.
Two male, spayed and de-clawed, gray littermates were used interchangeably at
CA and LY. A third cat, a black male that was spayed and de-clawed, was used at
Leisure Lakes. A cat carrier was utilized to transport and hold each cat until the top was
opened for exposure to the birds. Cats were leashed and attached to the carrier at all

12
times in the field. A cat was placed in a noticeable area near scrub when one or more
adult FLSJ were present. The trial ended when either the birds flew away or if the cat
became unsettled and was removed for its own well-being. Again, this resulted in an
average of 3 minutes of recording time per session.
Analysis of calls.—The predator screech scold, or alarm call, ranges between 1-8
kHz (Francis et al. 1989; Elowson and Hailman 1991). Francis et al. (1989) further
described the alarm call as a harsh vocalization often given consecutively in a long series
but sometimes singly. For further description of the predator alarm call, see Barbour
(1977) and Elowson and Hailman (1991).
Calls were recorded using a shotgun microphone (Audio-technica U.S.,
line+gradient condenser microphone, frequency response 20-20,000Hz, Audio-technica
U.S., Inc., Stow, Ohio), which was mounted on a 54 cm parabolic dish. The microphone
was integrated with an audio interface (Tascam US-122 USB, 24-bit depth, 44 kHz rate)
to digitally convert the calls for analysis into a COMPAQ laptop running the sound
analysis software Raven 1.2.1 (Cornell Lab of Ornithology) (Fig. 2). The first step was
to run calls through a 1,000 Hz High Pass filter to reduce background noise.
Spectrograms were then created in Raven with the following parameters: 128 point frame
length (1398.80 Hz bandwidth), 256 point Fourier transform size, Hamming, 512 samples.
Four random adult FLSJ calls from five randomly selected family groups in each
subpopulation for each of the three predators, a total of 180 calls, were measured for two
acoustical variables using Raven. The maximum frequency “Max Frequency” function,
as demonstrated by Miller, Vanderwerf, and McPherson (2003), and the duration, or
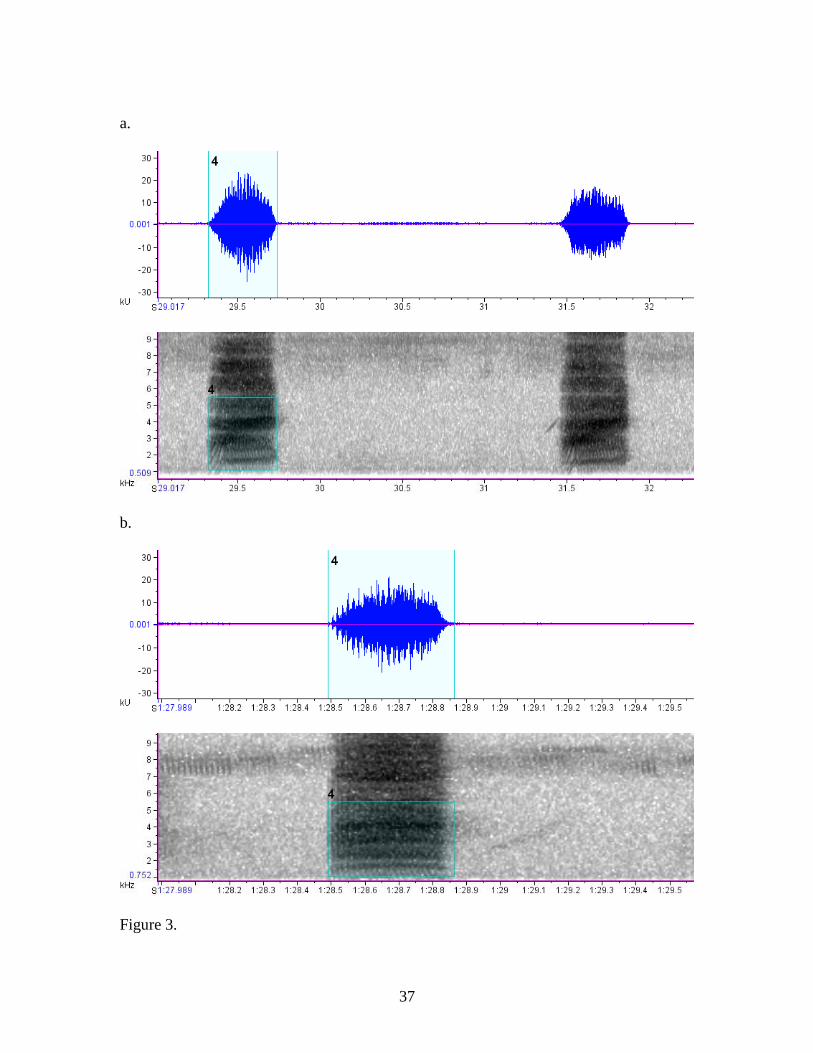
lengths, of the calls were analyzed (Fig. 3).

13
Statistical analyses.—For this study, a one-way analysis of variance (ANOVA)
was used to test differences between subpopulations (i.e. cat treatment between the three
subpopulations) and treatments within a subpopulation (i.e. between the cat, artificial and
live snake at LY). When statistical significance was determined, a Tukey Test (mean
separation test) was conducted.
Two datasets—maximum frequency for the artificial snake recordings between
subpopulations and call duration for the cat recordings between subpopulations—did not
follow normal distributions; thus, non-parametric analyses were utilized. Therefore, a
Kruskal-Wallis test, which acts similarly to a one-way ANOVA and is not dependent on
normal distribution, was used. This was followed by a non-parametric mean separation
test, the Mann-Whitney test, to determine specifically which treatment means were
significant.
RESULTS
Twenty calls per stimulus for a total of sixty calls per subpopulation (180 calls in
all) were analyzed for the two acoustic variables—max frequency and call duration. Data
were analyzed by treatments and subpopulations.
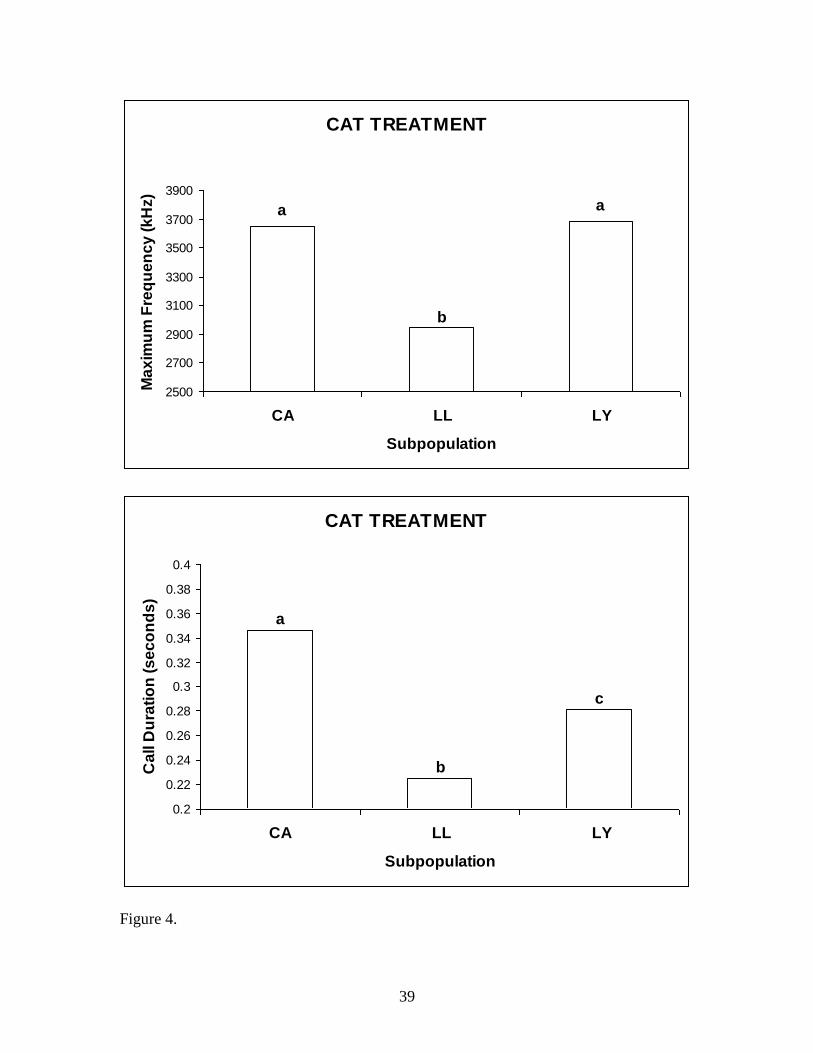
Cat treatment between subpopulations.—A one-way ANOVA for the average
max frequency, with normal distribution, was significant (P < 0.001). LL’s average max
frequency was significantly less (Tukey test, P < 0.001) than CA and LY. However, LY
and CA were not significantly different from each other. A Kruskal-Wallis test for the
average call duration, with non-normal distribution, revealed significance between
subpopulations (P < 0.001). All subpopulations were found to be significantly different
(Mann-Whitney test, P < 0.001) from each other (Fig 4).

14
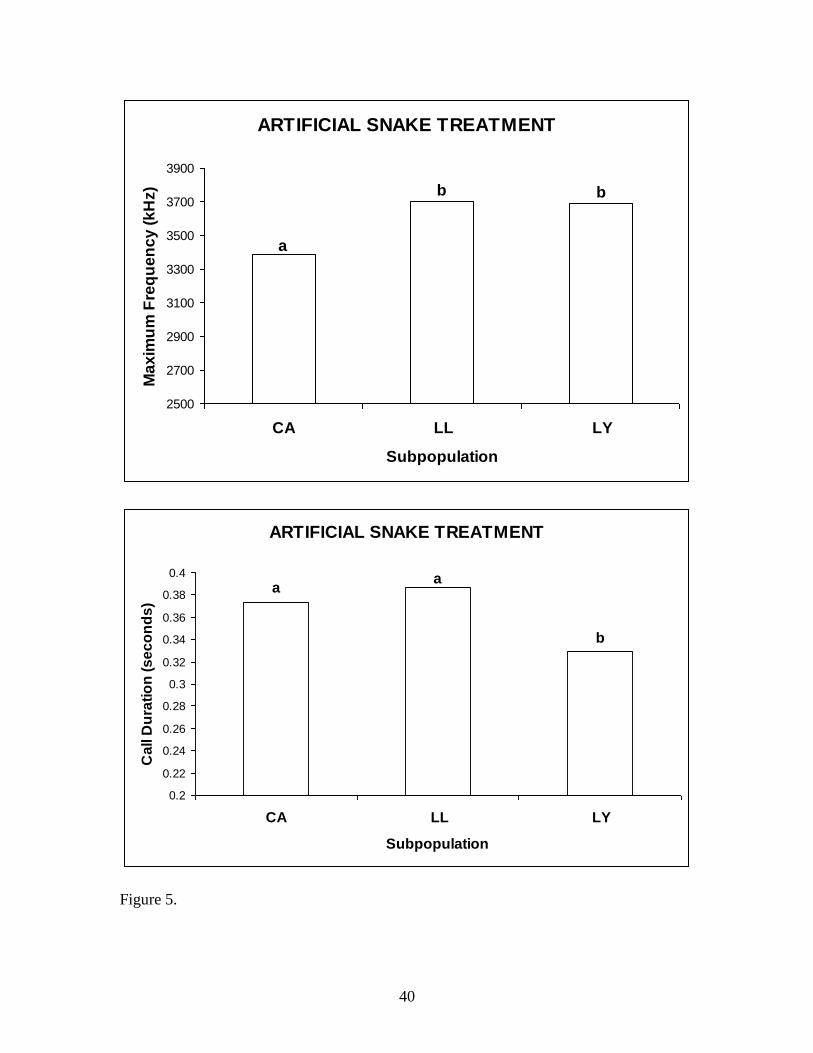
Artificial snake treatment between subpopulations.—A Kruskal-Wallis test for the
average max frequency, with non-normal distribution, revealed significance between
subpopulations (P = 0.01). CA was significantly less than LL and LY (Mann-Whitney
test, P < 0.01), but LL and LY were not significantly different from each other. A one-
way ANOVA for call durations, with normal distribution, between subpopulations was
significant (P = 0.005). The average call duration of LY was significantly less (Tukey
test, P = 0.005) than CA and LL, but there was no significant difference between CA and
LL (Fig 5).
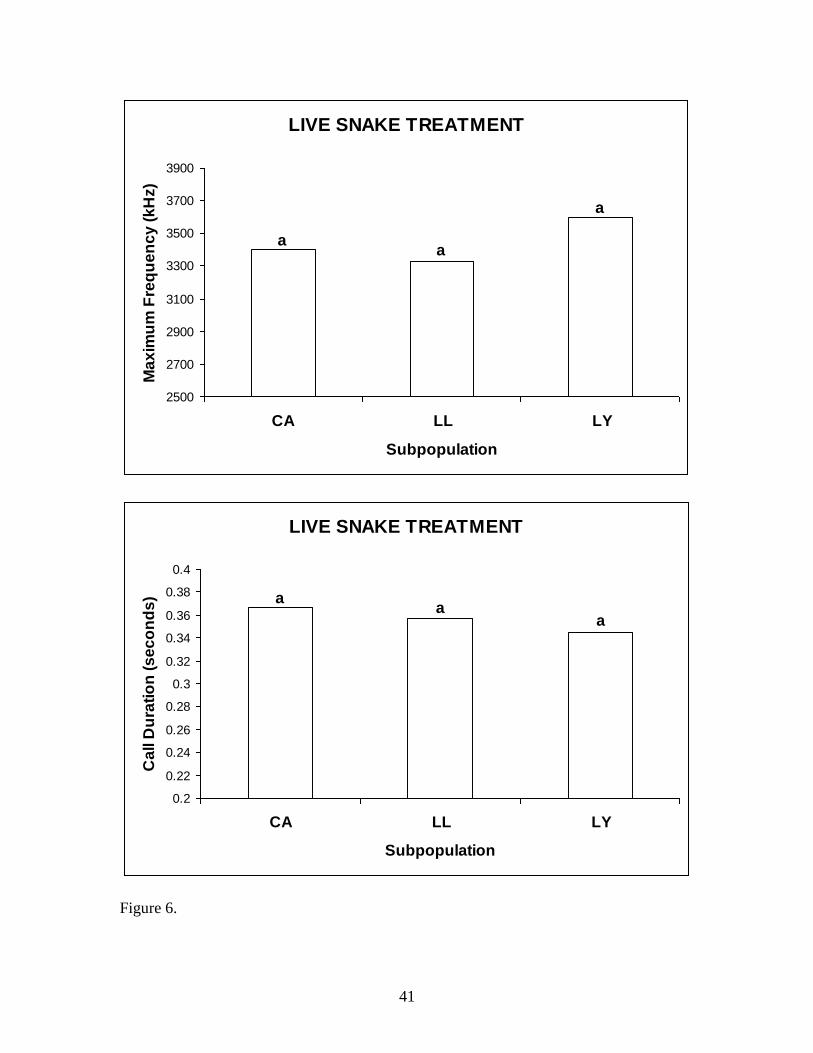
Live snake treatment between subpopulations.—A one-way ANOVA was not
significant for FLSJ responses to the live snake treatment at any of the three
subpopulations for the average max frequency (P = 0.236) or call duration (P = 0.572)
(Fig 6).
All treatments within CANA/MINWR.—A one-way ANOVA found no significant
differences for the average max frequency (P = 0.109) and average call duration (P =
0.229) between any treatment (Fig 7).
All treatments within Leisure Lakes.—A one-way ANOVA for the average max
frequency was significant (P < 0.001). The cat, artificial snake, and live snake treatments
were significantly different from each other (Tukey Test, P < 0.001). A one-way
ANOVA for the average call duration was significant (P < 0.001). The average call
duration for the cat treatment was significantly less (Tukey Test, P<0.001) than both
snake treatments, but the snake treatments were not significantly different from each
other (Fig 8).

15
All treatments within Lyonia Preserve.—In a one-way ANOVA analysis, there
were no significant differences found for the average max frequency between any
treatment (P = 0.766). A one-way ANOVA for the average call duration was significant
(P < 0.001). Similar to LL, the average call duration for the cat treatment was
significantly less (Tukey Test, P < 0.001) than both snake treatments, but the snake
treatments were not significantly different from each other (Fig 9).
DISCUSSION
Although the FLSJ has been studied extensively given its unique natural history,
its acoustic repertoire has typically been quantified and classified with minimal scientific
inquiry until recently. This study addressed dialectic variation between FLSJ
subpopulations with one particular acoustic entity, the predator alarm call. Dialectic
variation between FLSJ subpopulations has only been recently examined; using the
hiccup call, Russel (unpubl. data 2005) determined that there is indeed dialectic variation.
The current study confirms this conclusion, affirming dialectic variation within this bird
species.
Dialectic variation.—Dialect formation is widespread in many bird species (Harris and
Lemon 1971) and, therefore, it is not surprising to find dialectic variation between all
three FLSJ subpopulations. Results from this study for both maximum frequency and
call duration revealed that significant differences do exist between subpopulations in
response to the cat treatment and artificial snake treatment, but not between
subpopulations in regards to the live snake treatment (Figs. 4-6). Hence, dialectic
variation occurs between all three subpopulations when responding to two different
predator stimuli, the cat and artificial snake, but not to the live snake. Genetic isolation

16
and song or call learning between generations are potential causes for this dialectic
difference.
In general, dialectic variation has been extensively studied given its adaptive
significance for cultural-learning of songs and potential genetic isolation of populations
(Lachlan and Servedio 2004). Lemon (1966) suggested that dialects are influenced by
three factors: isolation of populations, generation to generation transmission of a dialect,
and a source of variation for the dialectic divergence. Since the FLSJ is highly sedentary
and disperses minimally away from its natal territory, isolation of the larger sub-regions
and even smaller subpopulations is becoming an influencing factor in this dialectic
divergence. Isolation, given time, often leads to genetic differentiation between
populations.
Song-learning.—Transmission of a dialect would be readily possible given the FLSJ
family structure, in which helpers often remain with the breeder pair for a year or more
(Woolfenden and Fitzpatrick 1996), thereby learning that particular dialect from the
family unit. This cultural-learning phenomenon is evident in many bird species, such as
Saddlebacks (Jenkins 1977), Indigo Buntings (Payne et al. 1981), and Chaffinches
(Lynch and Baker 1993), for example. In cultural transmission, or song learning, a song
or call is passed from one individual to another (Slater 1986). This type of song and call-
learning has not been extensively studied in the FLSJ. Questions Baptista (1975) posed
regarding song dialects among another non-migratory species, like the Nuttall’s White-
crowned Sparrow, may be appropriate to apply to the FLSJ given their similar natural
histories. Baptista addressed the following: What impacts do songs have on the gene
flow between neighboring populations? What is the state of song adaptation or

17
morphology on the borders of those populations with differing dialects? And, what
environmental “barriers” exist between populations?
The two initial questions Baptista (1975) posed would be interesting to apply to
FLSJ populations within a sub-regional area, for example, the Atlantic coast. A
comparison of a FLSJ call, studied or not previously studied acoustically, of families
within this region (CANA/MINWR to Kennedy Space Center) could yield interesting
comparisons. Baptista’s third question about potential environmental barriers is
addressed here.
Habitat fragmentation.—Another source for this dialectic variation may be due to the
highly fragmented nature of FLSJ habitat and disjointed landscape. Urbanization and
other anthropogenic sources (i.e. roads) have contributed greatly to the habitat loss that
has occurred across Florida (USFWS 1999). A disjointed landscape was evident in the
most urban study site (LY), where the 360 acre preserve of restored scrub sits within a
residential community (Fig. 10). The semi-rural site (LL) was broken up by roads and
interspersed with residential houses but it has an adjacent managed scrub habitat (Fig. 10).
The most rural site (CA) has had minimal human impact and has progressive
management practices for the present scrub habitat (Fig. 10).
With persistent pressures from habitat fragmentation and urbanization, FLSJ
survivability continues to significantly decrease, thereby increasing their risk for
extinction (Woolfenden and Fiztpatrick 1991; Breininger 1999, Breininger and Carter
2003). Several studies have examined the effects of habitat fragmentation and FLSJ
demographics within populations (see Breininger et al. 1996; Breininger 1999;
Breininger et al. 1999, Bowman and Woolfenden 2001). Breininger (1999) reported that

18
almost 30% of existing FLSJ populations inhabit fragments of habitats surrounded by
residential development. Breininger (1999) stated dispersal distances were longer in
urban areas than in rural or more natural areas. Also, demographic success (i.e.
population characteristics such as mortality, nesting, or fledgling success) in the FLSJ
was reduced in fragmented urban populations.
Since the FLSJ requires specific habitat conditions, habitat suitable for population
expansion is becoming increasingly harder to find, hence the longer dispersal distances in
urban settings. Thaxton and Hingtgen (1996) found that the FLSJ disperse from
fragmented habitat into larger land tracts but not from larger contiguous habitats to
smaller patchy habitats. These results, coupled with the limited average FLSJ dispersal
distance (1.6 km, Fitzpatrick 1999), may indicate a severe population interaction
limitation, even within the core populations found along the Atlantic and Gulf coasts, and
Lake Wales Ridge.
In addition to the factors mentioned previously, what else may be contributing to
this dialectic variation? This question may be answered by the second objective of this
study, which was to compare the response calls to three diverse ground predators within
each of the subpopulations.
When populations are separated from one another, thereby not communicating
with one another, one source of variation for dialectic divergence could be due to
different environmental stimuli (Nottebohm 1969). For instance, Thorington and
Bowman (2003) proposed that predator communities in urban and natural habitats follow
species-specific predation patterns. In suburban habitats, predators such as birds, rodents,
snakes, and terrestrial mammals may increase due to food availability. This seems most

19
likely, especially with cats. Different predator pressures may be influencing the dialectic
variation in the predator alarm calls within at least two study subpopulations.
Results from this part of the study for both max frequency and call duration
indicated significant differences in FLSJ responses to ground predators within LL and LY,
but not to ground predators within the CA subpopulation (Figs. 7-9). It is evident that
these subpopulations are exposed to different predator threats that, at this time, are
generating different acoustical responses.
Live snake.—There were no significant differences in response to the live snake
treatment between any of the subpopulations (Fig. 6). It is plausible that all of these
subpopulations have evolved with snakes and several snake species still exist in the
habitats of each subpopulation. Their response to the snake may have been influenced
mainly by the size of the snake stimulus (at least 58 in) but also perhaps due to its species
(Elaphe guttata). Elaphe guttata is a common snake found across Florida and is found
not only in rural areas but urban sites as well. Snakes account for the majority of FLSJ
deaths, especially young juveniles or nestlings (McGowan and Woolfenden 1989; Schaub
et al. 1992). Thorington and Bowman (2003) suggested that snake predation may
increase with human density because urban regions provide an area where nest predators
could flourish. However, there are few data that exist on snake communities and the
effects of urbanization on their populations and abundance or lack thereof in a suburban
setting (Bowman and Woolfenden 2001).
Artificial snake.—Some variation was evident in how the subpopulations responded to
the artificial snake. CA max frequency response to the artificial snake was significantly
different from LL and LY, whereas LL and LY were not significantly different from each

20
other (Fig. 5). Additionally, LY was significantly different from both LL and CA
subpopulations for the average call duration; however, there was not a difference in call
duration between CA and LL (Fig. 5).
The artificial snake mimicked an Eastern Indigo, which is a common snake
predator on the FLSJ (McGowan and Woolfenden 1989). However, due to the natural
history of this snake, it is more likely to frequent habitat-specific rural areas (USFWS
1999). This could explain why the max frequency of response to the artificial snake was
significantly different between CA and the other two sites. CA is the most natural, with
little human interference to significantly impact this snake’s habitat (Fig. 10). Duration
data indicated a significant difference between the two rural subpopulations (CA and LL)
with the more urban subpopulation (LY), but not between the rural subpopulations
themselves. Even low human density and development can impact snake populations
like the Eastern Indigo (Moler 1992; Bowman and Woolfenden 2001), thereby, again,
reinforcing perhaps D. couperi preference for the most rural areas. The two rural
subpopulations (CA and LL) have more exposure and awareness of the threat Eastern
Indigos pose compared to the urban subpopulation (LY) having little or no exposure to
these snakes. This could account for the different acoustic response to the artificial snake
treatment.
Domesticated cat.—An interesting aspect of this study observed cats as ground predators.
It is estimated that 60-100 million free-ranging cats exist within the United States (Mott
2004). Some experts estimate these cats kill millions of birds each year. Bobcats are a
natural predator of FLSJ, however, lower density and territoriality limit their impact on
the FLSJ. Domestic cats exist in high densities and are not territorial. This results in a

21
much more significant impact on local bird populations (Florida Fish and Wildlife
Conservation Commission 2001).
Results from the cat treatment were variable. For max frequency, LL was
significantly different from both CA and LY, while CA and LY were not different from
each other (Fig. 4). Interestingly, for call duration, all populations were significantly
different from each other. Lack of a clear pattern in the response of the FLSJ to domestic
cats may indicate that these birds could still be developing a response to this new
predation pressure. The more urban subpopulation (LY) has likely had the most
experience with cats as predators as opposed to the more rural sites (CA and LL) (Fig.
10). The FLSJ at CA responded to the cat similarly as to the other two predators,
therefore, there was no differentiation in threat levels presented by either the snakes or
cat. At LL, the more semi-rural subpopulation, data indicate perhaps a learning process
occurring within FLSJ families and their relationship with this emerging predator. A
dialectic divergence may become more distinct as the birds at LL are increasingly
exposed to cats. Their behavioral and acoustical response would continue to adapt to the
type of threat cats pose. Cats could easily be more apt to take fledglings and adults than
any other predator. In fact, cats could be one of the most burgeoning predator threats to
adult FLSJ. This could have significant ramifications for FLSJ subpopulations if they do
not learn and adapt their responses quickly to deal with this “new” predator.
Cats are often present in larger numbers near urban sites, even forming colonies,
because of the available food source provided by humans (Florida Fish and Wildlife
Conservation Commission 2001). If, by virtue of their relationship to urban areas, these

22
subpopulations continue to experience significantly different levels of predation by cats,
eventual dialectic variation or even extirpation of those subpopulations could occur.
Conclusions and Conservation Implications.—In the past, the FLSJ has evolved with and
adapted to predator threats with consistent responses as evidenced in this study with their
responses to the live snake. Historically, the FLSJ subpopulations were more connected,
but, today, the ever-growing urbanization has disrupted much of the optimal and
continual FLSJ habitat. This threat, along with increased exposure to cats, may occur in
random and isolated areas affecting the FLSJ sporadically. Therefore, subpopulations
would not have equal exposure to these new threats. This concomitant exposure of the
subpopulations to varying stimuli, including diverse ground predators, could encourage
greater dialectic variation.
Extinction of FLSJ subpopulations is the end result of habitat fragmentation (i.e.
population isolation). If further fragmentation of FLSJ habitat occurs, speciation is also
likely, given time and rate of divergence; however, the FLSJ faces the greater threat of
going extinct before full speciation could occur.
Further Recommendations.—Because of the threats facing FLSJ subpopulations
today, suggestions for future study are mentioned. One recommendation is to conduct
research on FLSJ calls in conjunction with behavior. In addressing both behavior and
calls, for instance, re-analyzing the predator alarm call to diverse predators (especially to
a cat), could potentially provide insightful understanding of dialectic divergence.
A second recommendation is to repeat this study with other FLSJ subpopulations
across Florida or even within a sub-region (Atlantic coast). As mentioned previously,
two initial questions posed by Baptista (1975) addressed song adaptation and learning,

23
dialect formation, and gene flow among neighboring populations. This study could be
repeated with the focus of investigating these questions.
Thirdly, other FLSJ calls could be studied to determine dialectic occurrences
between and/or within subpopulations. Some call examples for future analyses include
the aerial predator alarm weep or the rapid multiple weep.
Lastly, conduct further studies of free-ranging cats as predators of FLSJ to
determine their impact on this threatened species. By determining the impact and level of
threat that free-ranging cats have upon the FLSJ, greater conservation initiatives could be
employed for the FLSJ. Ultimately, the research from this study and future studies
concerning dialectic variation could provide understanding on the rate of divergence,
prospect for speciation, and the potential of extinction of this threatened species.

24
ACKNOWLEDGEMENTS
In Florida, I would like to warmly thank John Stiner and Candace Carter of
CANA/MINWR, Bonnie Cary and Sarah Cleveland of Volusia Co. Environmental
Management, and Reed Bowman of Archbold Biological Station for guidance, help, and
permission of study site use. I would like to thank Becky Bolt, at The Dynamac
Corporation of Kennedy Space Center, and volunteers who lent time and/or animals—
Jenn Stiner, Marcia Rickey, and students involved in Park Youth Workers at
CANA/MINWR. In Indiana, a huge thanks to my advisor Jan Reber for all her time and
patience regarding this project, Ken Constantine and Rob Reber for their wonderful help
on statistics, Bob Davis for his expertise in acoustics, Dick Squiers and Leland Boren for
the opportunity to participate in a great environmental science program, and Kory Russel
for the pioneer work of acoustical analyses in FLSJ subpopulations across Florida.

25
LITERATURE CITED
Ackers, S. H. and C. N. Slobodchikoff 1999. Communication of stimulus size and shape
in alarm calls of Gunnison’s prairie dogs, Cynomys gunnisoni. Ethology 105(2):149-162.
Apel, K. M. and M. S. Ficken. 1981. Predator alarm calls of young Black-capped
Chickadees. The Auk 98: 624.
Baker, M. C. and A. M. Becker. 2002. Mobbing calls of Black-capped Chickadees:
effects on urgency on call production. Wilson Bulletin 114(4): 510-516.
Baker, M. and D. Logue. 2003. Population differentiation in a complex bird sound: A
comparison of three bioacoustical analysis procedures. Ethology 109: 223-242.
Baker, M. C. and M. A. Cunningham. 1985. The biology of bird-song dialects. The
Behavioral and Brain Sciences 8: 85-133.
Barbour, D. B. 1977. Vocal communication in the Florida scrub jay. Master’s thesis,
University of South Florida, Tampa.
Bowman R. and G. E. Woolfenden. 2001. Nest success and the timing of nest failure of
Florida scrub-jays in suburban and wildland habitats. In: Marzluff, J. M., Bowman, R.
and Donnelly, R. (eds), Avian Ecology and Conservation in an Urbanizing World.
Kluwer Academic Publishing, pp. 383-402.

26
Bowman, R. and G. E. Woolfenden. 2002. Nest site selection by Florida Scrub-Jays in
natural and human-modified habitats. Wilson Bulletin 114(1): 128-135.
Breininger, D. R., V. L. Larson, D. M. Oddy, R. B. Smith, and M. J. Barkaszi. 1996.
Florida Scrub-Jay demography in different landscapes. The Auk 113(3): 617-625.
Breininger, D. R., M. A. Burgman, and B. M. Stith. 1999. Influence of habitat quality,
catastrophes, and population size on extinction risk of the Florida scrub-jay. Wildlife
Society Bulletin 27(3): 810-822.
Breininger, D. R. and G. M. Carter. 2003. Territory quality transitions and source-sink
dynamics in a Florida Scrub-Jay population. Ecological Applications 13(2): 516-529.
Brown M. M., Kreiter N. A., Maple J. T., Sinnott J. M. 1992. Silhouettes elicit alarm
calls from captive vervet monkeys (Cercopithecus aethiops). Journal of Comparative
Psychology 106(4): 350-359.
Cheney, D. and R. Seyfarth. 1990. How monkeys see the world. Chicago: University of
Chicago Press.
Cunningham, M. A. and M. C. Baker, T. J. Boardman. 1987. Microgeographic song
variation in the Nuttall’s White-crowned Sparrow. The Condor 89: 261-275.

27
Elowson, A. M. and J. P. Hailman. 1991. Analysis of complex variation: dichotomous
sorting of predator-elicited calls of the Florida scrub jay. Bioacoustics: The International
Journal of Animal Sound and Its Recording 3: 295-320.
Fischer J., K. Hammerschmidt, D. L. Cheney, and R. M. Seyfarth. 2001. Acoustic
features of female chacma baboon barks. Ethology 107(1): 33-54.
Fitzpatrick, J. W., G. E. Woolfenden, and M. T. Kopeny. 1991. Ecology and
development-related habitat requirements of the Florida Scrub Jay (Aphelocoma
coerulescens coerulescens). Florida Game and Fresh Water Fish Commission, Nongame
Wildlife Program Technical Report NO. 8. pp. 49.
Fitzpatrick, J. W., R. Bowman, D. Breininger, M. O’Connell, B. Stith, J. Thaxton, B.
Toland, and G. Woolfenden. 1994. Habitat conservation plans for the Florida Scrub
Jay: a biological framework. Unpublished draft report. On file at U.S. Fish and Wildlife
Service, South Florida Ecosystem Office; Vero Beach, Florida.
Fitzpatrick, J. W., G. E. Woolfenden, and R. Bowman. 1999. Dispersal distances and its
demographic consequences in the Florida Scrub-Jay. In; Proc. 22nd International
Ornithological Congress, Adams, N.J. and R. H. Slotow (Eds.). Birdlife South Africa. pp.
2465-2479.
Florida Fish and Wildlife Conservation Commission. 2001. Impacts of feral and free-

28
ranging domestic cats on wildlife in Florida. Florida Fish and Wildlife Conservation
Commission.
Francis, A. M., J. P. Hailman, and G. E. Woolfenden. 1989. Mobbing by Florida scrub
jays: behaviour, sexual asymmetry, role of helpers and ontogeny. Animal Behavior 38:
795-816.
Harris, M. A. and R. E. Lemon. 1971. Songs of song sparrows (Melospiza melodia):
individual variation and dialects. Canadian Journal of Zoology 50: 301-309.
Jenkins, P. 1977. Cultural transmission of song patterns and dialect development in a
free-living bird population. Animal Behavior 25: 50-78.
Kroodsma, D. E. 1974. Song learning, dialects, and dispersal in the Bewick’s wren. Z.
Tierpsychology 35: 352-380.
Lachlan, R. F. and M. R. Servedio. 2004. Song learning accelerates allopatric speciation.
Evolution 58(9): 2049-2063.
Lemon, R.E. 1966. Geographic variation in the song of Cardinals. Canadian Journal of
Zoology 44: 413-428.
Laiolo, P., A. Rolando, A. Delestrade, and A. De Sanctis. 2001. Geographic

29
diversification in the call repertoire of the genus Pyrrhocorax (Aves, Corvidae).
Canadian Journal of Zoology 79: 1568-1576.
Lynch, A. and A. J. Baker. 1993. A population memetics approach to cultural evolution
in Chaffinch song: meme diversity within populations. The American Naturalist 141(4):
597-620.
Lynch, A. and A. J. Baker. 1994. A population memetrics approach to cultural evolution
in Chaffinch song: differentiation among populations. Evolution 48: 351-359.
MacDougall-Shackleton E. A. and S. A. MacDougall-Shackleton. 2001. Cultural and
genetic evolution in Mountain White-crowned Sparrows: song dialects are associated
with population structure. Evolution 55(12): 2568-2575.
McGowan, K. J. and G. E. Woolfenden. 1989. A sentinel system in the Florida scrub jay.
Animal Behavior 37: 1000-1007.
Menges, E. S. 1999. Ecology and conservation of Florida scrub. In Savannas, Barrens,
and rock outcrop plant communities of North America (R.C. Andersons, J.S. Fralish, and
J.M. Baskin, Eds.) Cambridge University Press.
Miller, E. H., E.Vanderwerf, and L. McPherson. 2003. Vocalizations of the Tuamoto
Sandpiper Prosobonia cancellata. Wilson Bulletin 115(4): 455-463.

30
Moler, P. E. 1992. Eastern indigo snake, p. 181-186. In P. E. Moler (ed.), Rare and
Endangered Biota of Florida. Vol. 3, Amphibians and Reptiles. University Press of
Florida. Gainesville, Florida.
Morton, E. S. and M. D. Shalter. 1977. Vocal response to predators in pair-bonded
Carolina Wrens. The Condor 79: 222-227.
Mott, M. 2004. U.S. faces growing feral cat problem.
http://news.nationalgeographic.com/news/2004/09/0907_040907_feralcats.
Nottebohm, F. 1969. The song of the Chingolo, Zonotrichia capensis, in Argentina:
description and evaluation of a system of dialects. Condor 71: 299-315.
Payne, R. B., W. L. Thompson, K. L. Fiala and L. L. Sweany. 1981. Local song traditions
in Indigo Buntings: cultural transmission of behavior patterns across generations.
Behavior 77: 199-221.
Placer, J. and C. N. Slobodchikoff. 2000. A fuzzy-neural system for identification of
species-specific alarm calls of Gunnison’s prairie dogs. Behavioral Processes 52: 1-9.
Russel, K. 2005. Call variations between sub-regional Florida Scrub-Jay (Aphelocoma
coerulescens) populations. Thesis. Unpublished data.

31
Schaub, R., R. L Mumme, and G. E. Woolfenden. 1992. Predation on the eggs and
nestlings of Florida Scrub Jays. The Auk 109(3): 585-593.
Slater, P. J. B. 1986. The cultural transmission of bird song. TREE 1(4): 94-97.
Stenzler, L. M. and J. W. Fitzpatrick. 2002. Isolation of microsatellite loci in the Florida
Scrub-Jay Aphelocoma coerulescens. Molecular Ecology Notes 2: 547-550.
Stith, B. M., J.W. Fitzpatrick, G. E. Woolfenden, and B. Pranty. 1996. Classification and
conservation of metapopulations: a case study of the Florida scrub-jay. In: D.R.
McCullough (ed), Metapopulations and Wildlife Conservation Island. Washington D.C.,
pp. 187-215.
Thaxton, J. E. and T. M. Hingtgen. 1996. Effects of suburbanization and habitat
fragmentation on Florida Scrub-Jay dispersal. Florida Field Naturalist 24: 25-37.
Thorington, K. and R. Bowman. 2003. Predation rate on artificial nests increases with
human housing density in suburban habitats. Ecography 26: 188-196.
U. S. Fish and Wildlife Service. 1999. South Florida multi-species recovery plan. Atlanta,
Georgia. 2172 pp.
Waite T. A. and T. C. Grubb, Jr. 1987. Dominance, foraging and predation risk in the

32
Tufted Titmouse. The Condor 89: 936-940.
Walters, J. 1990. Anti-predatory behavior of Lapwings: field evidence of
discriminative abilities. Wilson Bulletin 102(1): 49-70.
Woolfenden, G. E. and J. W. Fitzpatrick. 1984. The Florida Scrub Jay: demography of a
cooperative breeding bird. Princeton University Press, Princeton, New Jersey.
Woolfenden, G. E. and J. W. Fitzpatrick. 1990. The Florida Scrub Jay: a synopsis after 18
years of study. In: Cooperative breeding in birds: long-term studies in ecology and
behavior. Ed. P.B. Stacey and W.D. Koenig. Cambridge University Press.
Woolfenden, G. E. and J. W. Fitzpatrick. 1991. Florida Scrub Jay ecology and
conservation. Pages 542-565 in Bird population studies (C.M. Perrins, J.D. Lebreton, and
G.J.M. Hirons, Eds.) Oxford University Press, New York.
Woolfenden, G. E. and J. W. Fitzpatrick. 1996. Florida scrub jay. The Birds of North
America No. 228: 1-26.

33
FIGURE LEGENDS
Figure 1. Five major sub-regions of the Florida Scrub-Jay including the three “core
populations” pertinent to this study. They include the Atlantic Coast, Ocala, and Lake
Wales Ridge sub-regions. The white stars indicate the study subpopulation locations
within each sub-region. Adapted by K. Russel (unpubl. data 2005) from Fitzpatrick et al.
1994 cited in USFWS 1999.
Figure 2. Diagram of the sound recording equipment and setup. Model from K. Russel
(unpubl. data 2005).
Figure 3. Duration of the call was measured from the waveform while max frequency
was taken from the spectrogram. a. Waveform and spectrogram figure from a family
group at CA responding to the cat treatment. b. Waveform and spectrogram figure from
a family at LL responding to the live snake treatment. c. Waveform and spectrogram
figure from a family at LY responding to the artificial snake treatment.
Figure 4. Mean max frequency and call duration to the cat treatment between
subpopulations. The different letters (a,b,c) indicate significance between subpopulations.
Figure 5. Mean max frequency and call duration to the artificial snake treatment between
subpopulations. The different letters (a,b,c) indicate significance between subpopulations.

34
Figure 6. Mean max frequency and call duration to the live snake treatment between
populations. There were no significant differences between subpopulations; hence the
letter “a” was applied for all three sites.
Figure 7. Mean max frequency and call duration between stimuli within CANA/MINWR
subpopulation. There were no significant differences between treatments; hence the letter
“a” was applied for all three treatments.
Figure 8. Mean max frequency and call duration between stimuli within Leisure Lakes
subpopulation. The different letters (a,b,c) indicate significance between treatments.
Figure 9. Mean max frequency and call duration between stimuli within Lyonia Preserve
subpopulation. The different letters (a,b,c) indicate significance between treatments.
Figure 10. Aerial photographs revealing landscape patterns in the three study sites with
the FLSJ families studied outlined in black. CANA/MINWR is the most rural, and study
families are located on land managed by the Fish and Wildlife Service, National Park
Service, and Kennedy Space Center. Leisure Lakes is a semi-developed residential
community adjacent to a state park with managed scrub habitat, and is therefore ‘semi-
rural.’ Lyonia Preserve, a 360 acre restored scrub ecosystem and managed scrub habitat,
is located within the City of Deltona and is surrounded by residential communities. The
scale for the zoomed in aerial photo on the right is 1 in to 200 m.

35
FIGURES
Figure 1.

36
Figure 2.
37
a.
b.
Figure 3.

38
c.
Figure 3.
39
CAT TREATMENT
a
b
a
2500
2700
2900
3100
3300
3500
3700
3900
CA LL LY
Subpopulation
Ma
xim
um
Fre
qu
en
cy
(k
Hz)
CAT TREATMENT
c
b
a
0.2
0.22
0.24
0.26
0.28
0.3
0.32
0.34
0.36
0.38
0.4
CA LL LY
Subpopulation
Ca
ll D
ura
tio
n (
se
co
nd
s)
Figure 4.
40
ARTIFICIAL SNAKE TREATMENT
bb
a
2500
2700
2900
3100
3300
3500
3700
3900
CA LL LY
Subpopulation
Ma
xim
um
Fre
qu
en
cy
(k
Hz)
ARTIFICIAL SNAKE TREATMENT
b
aa
0.2
0.22
0.24
0.26
0.28
0.3
0.32
0.34
0.36
0.38
0.4
CA LL LY
Subpopulation
Ca
ll D
ura
tio
n (
se
co
nd
s)
Figure 5.
41
LIVE SNAKE TREATMENT
a
aa
2500
2700
2900
3100
3300
3500
3700
3900
CA LL LY
Subpopulation
Ma
xim
um
Fre
qu
en
cy
(k
Hz)
LIVE SNAKE TREATMENT
aa
a
0.2
0.22
0.24
0.26
0.28
0.3
0.32
0.34
0.36
0.38
0.4
CA LL LY
Subpopulation
Ca
ll D
ura
tio
n (
se
co
nd
s)
Figure 6.

42
TREATMENTS WITHIN CANA/MINWR
aa
a
2500
2700
2900
3100
3300
3500
3700
3900
Cat Artificial snake Live snake
Treatment
Ma
xim
um
Fre
qu
en
cy (k
Hz)
TREATMENTS WITHIN CANA/MINWR
aa
a
0.2
0.22
0.24
0.26
0.28
0.3
0.32
0.34
0.36
0.38
0.4
Cat Artificial snake Live snake
Treatment
Ca
ll D
ura
tio
n (s
eco
nd
s)
Figure 7.

43
TREATMENTS WITHIN LEISURE LAKES
c
b
a
2500
2700
2900
3100
3300
3500
3700
3900
Cat Artificial snake Live snake
Treatment
Ma
xim
um
Fre
qu
en
cy (k
Hz)
TREATMENTS WITHIN LEISURE LAKES
b
b
a
0.2
0.22
0.24
0.26
0.28
0.3
0.32
0.34
0.36
0.38
0.4
Cat Artificial snake Live snake
Treatment
Ca
ll D
ura
tio
n (s
eco
nd
s)
Figure 8.

44
TREATMENTS WITHIN LYONIA PRESERVE
aaa
2500
2700
2900
3100
3300
3500
3700
3900
Cat Artificial snake Live snake
Treatment
Ma
xim
um
Fre
qu
en
cy (k
Hz)
TREATMENTS WITHIN LYONIA PRESERVE
bb
a
0.2
0.22
0.24
0.26
0.28
0.3
0.32
0.34
0.36
0.38
0.4
Cat Artificial snake Live snake
Treatment
Ca
ll D
ura
tio
n (s
eco
nd
s)
Figure 9.

45
CANA/MINWR
LEISURE LAKES
Figure 10.

46
Figure 10.
LYONIA PRESERVE


















