
An update on neurobiological mechanisms involved in the ...
24
1 An update on neurobiological mechanisms involved in the development of chemotherapy- induced cognitive impairment (CICI) Laura Catalina Murillo Espitia Departamento de Biología, Facultad de Ciencias, Pontificia Universidad Javeriana [email protected] Abstract Chemotherapy is one of the most effective treatments against cancer. However, its toxic non- specificity affects both healthy and cancer cells. This toxicity generates side effects, being one of the most recognized the chemotherapy-induced cognitive impairment (CICI). Patients following chemotherapy report a decrease in cognitive abilities such as memory, learning, word finding and executive functions. Here I present a literature review on the main neurobiological mechanisms involved in CICI using a Boolean formula following the steps of the Preferred Reporting Items for Systematic Reviews and Meta-Analyses (Prisma) statement searching in several databases. It was found that main mechanisms described in the literature to explain CICI include neurotoxicity, decreased neurogenesis, loss of dendritic spines and dendritic complexity, altered neurotransmission, disruption of glial cells, loss of myelination and cytokine dysregulation leading to neuroinflammation. This review provides a general understanding of the neurobiological mechanisms underlying CICI and possible therapeutic targets to prevented. Further research is needed on alternative mechanisms related to the development and the severity of CICI and its prevention. Key words: chemobrain, cognitive impairment, chemotherapy drugs, CICI. 1. Introduction Cancer is one of the main causes of death worldwide (Ježek et al., 2021). In 2020, the World Health Organization reported 19’292.789 new cancer cases, being breast cancer the most prevalent type (World Health Organization, 2020). Hallmarks of these polygenic diseases are replicative immortality, angiogenesis induction, resisting cell death, sustaining proliferative signaling, evading growth suppressors, activating invasion and metastasis, cellular energetics dysregulation, genome instability and mutation, eluding immune destruction, and tumor-promoting inflammation (Hanahan & Weinberg, 2011). Due to cancer’s etiology complexity, each patient require different types of treatment or a combination of them, being chemotherapy and radiotherapy among the main treatments
Transcript of An update on neurobiological mechanisms involved in the ...

1
An update on neurobiological mechanisms involved in the development of chemotherapy-
induced cognitive impairment (CICI)
Laura Catalina Murillo Espitia
Departamento de Biología, Facultad de Ciencias, Pontificia Universidad Javeriana
Abstract
Chemotherapy is one of the most effective treatments against cancer. However, its toxic non-
specificity affects both healthy and cancer cells. This toxicity generates side effects, being one of the
most recognized the chemotherapy-induced cognitive impairment (CICI). Patients following
chemotherapy report a decrease in cognitive abilities such as memory, learning, word finding and
executive functions. Here I present a literature review on the main neurobiological mechanisms
involved in CICI using a Boolean formula following the steps of the Preferred Reporting Items for
Systematic Reviews and Meta-Analyses (Prisma) statement searching in several databases. It was
found that main mechanisms described in the literature to explain CICI include neurotoxicity,
decreased neurogenesis, loss of dendritic spines and dendritic complexity, altered neurotransmission,
disruption of glial cells, loss of myelination and cytokine dysregulation leading to neuroinflammation.
This review provides a general understanding of the neurobiological mechanisms underlying CICI
and possible therapeutic targets to prevented. Further research is needed on alternative mechanisms
related to the development and the severity of CICI and its prevention.
Key words: chemobrain, cognitive impairment, chemotherapy drugs, CICI.
1. Introduction
Cancer is one of the main causes of death worldwide (Ježek et al., 2021). In 2020, the World Health
Organization reported 19’292.789 new cancer cases, being breast cancer the most prevalent type
(World Health Organization, 2020). Hallmarks of these polygenic diseases are replicative
immortality, angiogenesis induction, resisting cell death, sustaining proliferative signaling, evading
growth suppressors, activating invasion and metastasis, cellular energetics dysregulation, genome
instability and mutation, eluding immune destruction, and tumor-promoting inflammation (Hanahan
& Weinberg, 2011). Due to cancer’s etiology complexity, each patient require different types of
treatment or a combination of them, being chemotherapy and radiotherapy among the main treatments

2
given their effectiveness in increasing patients survival with different types of cancer (Wen et al.,
2021).
Several chemotherapeutic drugs are used in cancer treatment, bring classified by the mechanism of
action. Alkylating agents (cyclophosphamide, cisplatin, carboplatin) and nitrosoureas like carmustine
are DNA crosslinkers (Cheung-Ong et al., 2013), anti-metabolites such as fluorouracil and
methotrexate inhibit DNA synthesis, plant alkaloids (vincristine, paclitaxel, docetaxel) induce
apoptosis through DNA damage (Habli et al., 2017), anti-tumor antibiotics as doxorubicin and
mitoxantrone inhibits topoisomerase II (Castel et al., 2017), or inhibits the synthesis and repair
mechanisms of DNA (Bhattacharya & Mukherjee, 2015), all of them reducing viability of rapid
dividing cells. Nevertheless, an important issue of chemotherapy drugs is their effects on healthy and
cancer cells, inducing multiple side effects.
One side effect of chemotherapy of non-central nervous system cancers is known as “chemobrain”,
“chemo fog”, or Chemotherapy-Induced Cognitive Impairment (CICI), which manifest during or after
treatment and includes learning, memory, attention, and executive function impairment in varying
degrees (Kaplan et al., 2016; Shi et al., 2019). Epidemiological studies in the United States record
that as of 2019 there were 16.9 million cancer survivors (American Cancer Society, 2020). Within
survivors, the prevalence of cognitive impairment after chemotherapy is estimated to be 15%-75%,
and up to 17%-35% of them suffer long-term effects (Moore et al., 2019; Myers et al., 2008).
Common symptoms reported by patients suffering CICI include inability to solve problems, lack of
concentration, difficulty finding the right words when speaking, memory loss, learning disabilities,
impaired executive functions and processing speed (Moore et al., 2019; Ren, Boriero, et al., 2019);
other neurobehavioral changes affecting the patient's quality of life after chemotherapy are anxiety
and depression (Liao et al., 2018). Even when it has been reported that just having cancer diagnosis
can affect cognitive function, evidence supports that chemotherapy further alters proper cognitive
functioning (Joly et al., 2015).
Considering epidemiological studies, it can be expected that an increasing number of new cases of
cancer and chemobrain will be reported. In this regard, current research has been focusing on
understanding the neurobiological mechanisms underlying CICI and searching for therapeutic targets
(Nguyen & Ehrlich, 2020; Stankovic et al., 2020). Although some mechanisms underlying CICI have
been described, knowledge about effects of different chemotherapeutics on cognition is constantly

3
growing. In this vein, this review aims to present a comprehensible update on the main
neurobiological mechanisms involved in CICI.
2. Methods
The literature review was developed following the steps of the Preferred Reporting Items for
Systematic Reviews and Meta-Analyses (Prisma) statement (Liberati et al., 2009). First, several
databases were selected from the electronic sources provided by the Pontificia Universidad Javeriana
(Academic Search Complete, Google Scholar, Nature, ProQuest, ScienceDirect, Scopus,
SpringerLink, Taylor & Francis and Web of Science). Then, the Boolean search was performed with
the following formula: chemobrain AND mechanism AND biochemical AND cellular. Inclusion
criteria were as defined as follows: original articles or reviews in English, peer-reviewed, published
in the period 2012-2021, with clinical, in vivo, and in vitro study models. Additional articles were
identified from the reference lists of previously selected articles and chosen for further review; in this
case, the date of publication was not considered.
All previously obtained articles were reviewed and duplicates were excluded from the list. Other
exclusion criteria were as follows: articles that corresponded to abstracts or conference reports,
articles on neurodegenerative diseases, computational approaches, neurobehavioral changes, or
articles in which the chemotherapy regimen was not specified. Prisma flowchart depicts the process
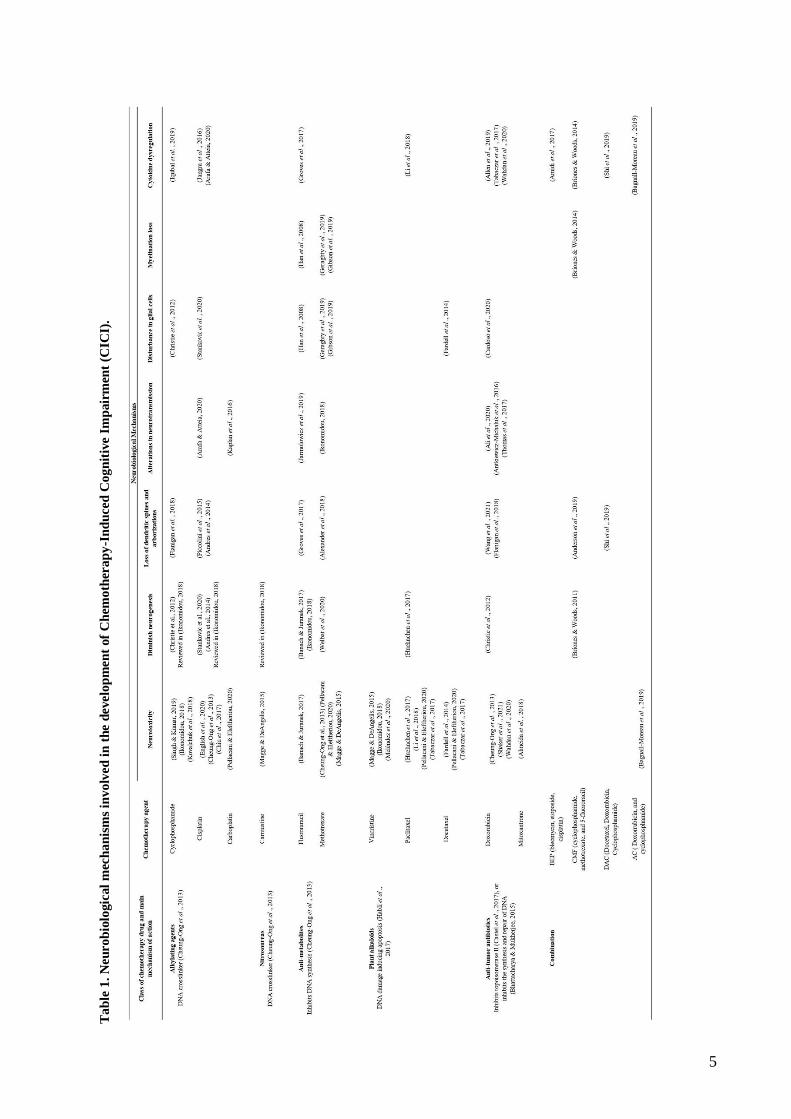
for articles and reviews selection (Fig. 1). Subsequently, the selected articles were read and analyzed,
and the information was summarized in Table 1. Finally, in the results section each of the
neurobiological mechanisms listed in the table was briefly described.

4
Figure 1. Preferred Reporting Items for Systematic Reviews and Meta-Analysis (PRISMA) flowchart for articles and
reviews search process. 408 documents were identified with the Boolean search in the following databases: Academic
Search Complete = 39, Google Scholar =157, Nature = 2, ProQuest = 50, ScienceDirect = 22, Scopus = 122, SpringerLink
= 12, Taylor & Francis, = 3 and Web of Science = 1. Additional documents from the reference lists = 11. Finally, 283
records were screened and finally 52 articles were selected for full-text review. Created with BioRender.com.
3. Results
In this section it will be briefly described the main neurobiological mechanisms underlying CICI
found in the literature search (Table 1) including neurotoxicity, decreased neurogenesis, loss of
dendritic spines and dendritic complexity, altered neurotransmission, disruption of glial cells, loss of
myelination and cytokine dysregulation leading to neuroinflammation (Fig. 2).
5
Ta
ble
1.
Neu
rob
iolo
gic
al
mec
han
ism
s in
volv
ed
in
th
e d
evelo
pm
en
t of
Ch
emo
ther
ap
y-I
nd
uced
Co
gn
itiv
e I
mp
air
men
t (C
ICI)
.

6
3.1 Neurobiological mechanisms
3.1.1 Neurotoxicity
Neurotoxicity is understood as the deleterious effect that a compound may have, or the indirect
metabolic alteration caused by that compound on the Central Nervous System (CNS) or peripheral
nervous system (PNS) (Pellacani & Eleftheriou, 2020). Notably, the CNS is integrated by two main
types of cells: neurons, and glial cells (astrocytes, oligodendrocytes, and microglia). The correct
functioning and development of CNS rely on the refined communication between these cells
(Pellacani & Eleftheriou, 2020), process that can be seriously impaired by chemotherapeutic agents.
Given that exogenous agents such as chemotherapeutic drugs have been reported to be toxic to the
nervous system and other organs such as, bone marrow and lungs, chemotherapeutics doses are
limited (Pellacani & Eleftheriou, 2020).
Specifically, it has been described in a murine model that intraperitoneal injections of
cyclophosphamide (CPA), cause histopathological damage in the cerebral cortex, characterized by
necrotic and apoptotic nuclei (Singh & Kumar, 2019). Additionally, it has been reported in a murine
model that exposure to CPA can change the expression of up to 200 genes in mice brains of their
progeny, suggesting that these changes in the transcriptomic profile may be affecting brain
development and function not only in the animal receiving the chemotherapy but also in its progeny
(Kovalchuk et al., 2018).
Similarly, intraperitoneal injections of paclitaxel in a murine model led an increase in apoptosis in
hippocampal neurons by increasing proinflammatory cytokines (Li et al., 2018). In addition, it has
also been reported that cisplatin induces mitochondrial dysfunction as evidenced by decreased
respiratory capacity and morphological alterations, events related to the poor performance of treated
animals in new object/place recognition (NOPRT) tests (Chiu et al., 2017). Furthermore, reduced
neuronal survival has also been reported using cisplatin in an in vitro model of cortical neurons
(English et al., 2020), in this particular case, astrocyte-mediated mitochondrial transfer induced a
recovery of cisplatin-treated neurons. Likewise, the cisplatin analog, carboplatin produces
mitochondrial damage and oxidative stress (Chiorazzi et al., 2015 reviewed in Pellacani &
Eleftheriou, 2020).
Also, the use of doxorubicin (DOX) induced histopathological damage showing nuclear pyknosis in
mice hippocampal neurons, particularly in the subiculum, fascia dentata and hilus, along with darker
cytoplasm and abnormal morphology (Shaker et al., 2021). Interestingly, in an in vitro assay

7
differentiated SH-SY5Y human neuronal cell model, the use of mitoxantrone was described to be
more toxic than DOX, supported by viability assays and the observation of apoptotic nuclei. Further
assays revealed that cytotoxicity was mediated by depolarization of the mitochondrial membrane
potential after 48 hours of exposure to chemotherapeutics (Almeida et al., 2018).
On the other hand, it has also been documented that intermittent treatment with docetaxel significantly
increased LC3-II formation (a marker of autophagy) in the hippocampus of treated mice (Fardell
et al., 2014). These autophagy processes are crucial in CNS homeostasis as they allow the degradation
and clearance of malfunctioning organelles, toxins, and pathogens (Pellacani & Costa, 2018).
Moreover, the systemic administration of fluorouracil reduced the expression of the Brain-Derived
Neurotrophic Factor (BDNF) levels in rat hippocampus, an important protein involved in
neurodevelopment and learning and memory (Mustafa et al., 2008 reviewed in Banach & Juranek,
2017). Also, it has been recently reported that another chemotherapeutic drug, vincristine, cause
destruction of hippocampal tissue when administered unilaterally or bilaterally to mouse (Meléndez
et al., 2020).
Taken this evidence together, it is likely that neurotoxicity is a key mechanism underlying CICI as it
directly impairs the homeostasis and viability of CNS cells, mainly neurons in important brain areas
involved in cognitive processes. Apparently, neurotoxic effects rely on oxidative stress,
mitochondrial dysfunction, and apoptosis.
3.1.2 Diminishing neurogenesis
Neurogenesis is a complex process in which a wide range of cellular and molecular events occur to
generate fully mature postmitotic neurons. In the adult mammal brain neurogenesis has been
documented in different regions including the olfactory bulb, neocortex, striatum, and hippocampus
(Snyder, 2019). Notably, special emphasis has been given to hippocampal neurogenesis due to its
relationship with higher cognitive functions, especially with learning and memory (Kempermann
et al., 2015).
In this vein it has been reported in a murine model that intraperitoneal injections of
cyclophosphamide and doxorubicin, caused a 47% and 53% decrease in the number of immature
doublecortin (DCX)-labeled neurons, respectively, when compared to saline treated controls; also,
ectopic migration of these immature neurons was observed (Christie et al., 2012). A decrease in
neurons labeled simultaneously with NeuN (marker for mature neurons) and BrdU (marker for
proliferative cells) was also observed after treatment with these chemotherapeutics, suggesting that

8
hippocampal neurogenesis was significantly impaired (Christie et al., 2012; Ikonomidou, 2018).
Similarly, the use of the CMF (cyclophosphamide, methotrexate and fluorouracil) regimen in an in
vivo model also resulted in a decrease of proliferative cells in the hippocampus, specifically in the
dentate gyrus (Briones & Woods, 2011).
Similar results have been observed after the systemic administration of fluorouracil, inducing a
decrease of DCX and BDNF positive cells in hippocampus (Mustafa et al., 2008 reviewed in Banach
& Juranek, 2017), and therefore indicating a disruption in hippocampal neurogenesis (Reviewed in
Ikonomidou, 2018). It should be noted that BDNF plays a critical role in the differentiation and
survival of new neurons, and reduction in its expression leads to memory deficits (Mustafa et al.,
2008).
Interestingly, it was reported that acute treatment with cisplatin causes an increase in apoptotic cells
in the CA1, CA3 and subgranular zone (SGZ) of the dentate gyrus in the hippocampus of treated male
rats (Andres et al., 2014). In the same study authors reported that cisplatin treatment resulted in the
depletion of neural stem cells (NSCs) cultured in vitro; however, NSCs were more resilient than
differentiated hippocampal neurons (Andres et al., 2014). Notably, antioxidant treatments can prevent
the deleterious effect of cisplatin and carmustine on hippocampal neurogenesis, suggesting that
oxidative stress is an important mechanism in this context (Reviewed in Ikonomidou, 2018; Reviewed
in Stankovic et al., 2020)..
Comparable whit these results, it was reported that intravenous injection of methotrexate in rats
significantly reduced DCX expression in the hippocampus. This effect was accompanied by a
decrease in BDNF and Nuclear factor erythroid-related factor (Nrf2) levels in both the hippocampus
and prefrontal cortex, both factors critical in the regulation of neurogenesis and subsequent
differentiation of NSCs (Kärkkäinen et al., 2014; Welbat et al., 2020). Decline in NSCs viability has
also been observed in paclitaxel treated cells in culture (Huehnchen et al., 2017).
As evidenced above, several chemotherapeutics diminish the expression of neurotrofic factors and
impaired neurogenesis in key areas for learning and memory, suggesting that impaired NSCs
production, migration and differentiation is a possible neurobiological mechanism underlying the
cognitive deterioration observed after the use of chemotherapeutics.
3.1.3 Loss of dendritic spines and arborizations

9
Neuronal connectivity and function are highly related to the number of dendritic spines and dendritic
tree complexity (Andres et al., 2014). Dendritic spines are specialized protrusions that are generally
located in the excitatory synapses (Chidambaram et al., 2019). These tiny structures have been
reported as critical sites in synaptic plasticity, where their morphology and density are essential for
memory and learning processes (Chidambaram et al., 2019). Four types of dendritic spines have been
proposed based in their morphology: mushroom spines, thin spines, stubby spines, and cup-shaped
spines, while mushroom spines are considered the most stable thin spines are considered the least
stable (Chidambaram et al., 2019). The potential importance of dendritic spines in memory and
learning has been described from the point of view of long-term potentiation (LTP). In this process,
the spines with the highest activity are those that will be maintained over time (Kasai et al., 2010).
Similarly, it has also been documented that the complexity of the dendritic tree has an important role
in memory formation and consolidation (Alexander et al., 2018).
Specifically, intravenous injection of cyclophosphamide or doxorubicin on ovariectomized mice
induce a reduction in the density of stubby dendritic spines in the dentate gyrus (Flanigan et al., 2018).
Similarly, intraperitoneal injection of doxorubicin in mice decrease the density of dendritic spines
and synaptic density in neurons of the CA1 region of the hippocampus (Wang et al., 2021). Also, in
vivo and in vitro model, it was documented that treatment with cisplatin caused a reduction in the
density of dendritic spines and dendritic branches in apical and basal dendrites in pyramidal neurons
of the CA1 and CA3 regions of the hippocampus (Andres et al., 2014). The synaptic damage induced
by cisplatin was not reversible, even after five days post-treatment (Andres et al., 2014).
Similar to this, it has been described in a juvenile murine model that the intrathecal application of
methotrexate considerably affects the dendritic architecture and reduces the density of mushroom
dendritic spines in CA1, CA3 and dentate gyrus of the hippocampus (Alexander et al., 2018).
Consistent with these results, it was reported in another murine model that the intraperitoneal
application of docetaxel, doxorubicin, and cyclophosphamide (DAC) significantly eliminate dendritic
spines of medial prefrontal cortex neurons (Shi et al., 2019).
More complex effects were reported in CMF-treated animals that exhibited a significant increase in
the number of stubby spines and a decrease in the number of mushroom-shaped spines in the DG,
while in CA1 and CA3 regions of the hippocampus there was a significant increase in the density of
dendritic spines (Anderson et al., 2019). However, decreases in the number of branch points, dendritic
ends, dendritic length, and dendritic complexity were observed in CMF-treated animals.

10
In addition, it has been described in an aged murine model that the use of intraperitoneal injections
of fluorouracil caused a decrease in the density of mushroom dendritic spines in the DG, while the
density of thin and stubby dendritic spines is increased. Accordingly, apical dendrites of pyramidal
neurons of the CA1 region showed decreased density of mushroom spines but increased density of
stubby spines; however, a decrease in thin spines was documented in the basal dendrites of pyramidal
neurons in CA3 (Groves et al., 2017). Also, it has been documented that fluorouracil negatively
affected dendritic complexity in DG neurons (Groves et al., 2017).
Taken together, these results indicate that chemotherapeutic treatment affects dendritic spine
dynamics and that there is also a tendency to decrease dendritic complexity in pyramidal neurons of
the hippocampus. One of the most marked trends was the decreased number of mushroom dendritic
spines and the increase in thin and stubby spines. Given that mushroom spines are the most stable
and the thin spines the least stable, changes in the density ratio of these spines could explain in part
the development of CICI.
3.1.4 Alterations in neurotransmission
Neuronal communication occurs via electrochemical gradient; the action potential travels through the
neuron cell body and generate neurotransmitter release to the synaptic cleft (Faber & Pereda, 2018).
It has been seen that the most important brain regions in cognition, such as the prefrontal cortex and
the hippocampus, are densely innervated with serotonergic, cholinergic and dopaminergic afferents.
Therefore, neurotransmitters such as serotonin, acetylcholine and dopamine (DA) are essential
modulators of memory and learning (Gonzalez-Burgos & Feriavelasco, 2008) (Levin, 2006). Also,
glutamate plays an important role in cognition, specifically in spatial memory, given that hippocampal
neurons are glutamatergic (Handra, 2019).
It has been described that intraperitoneal application of cisplatin-treatment induced a considerable
increase in brain acetylcholine levels, while generating a decrease in the relative expression levels
and activity of acetylcholinesterase (Arafa & Atteia, 2020). In contrast, in another in vivo model it
has been reported that the use of doxorubicin produces a reduction in the levels of acetylcholine in
both, hippocampus and prefrontal cortex (Ali et al., 2020). In this second study researchers found that
DOX treatment impaired learning and memory (Ali et al., 2020).
Also, decreased release of dopamine in the striatum was documented after intravenous application of
fluorouracil in a murine model, these findings were correlated with the decrease in attention shown
by these animals (Jarmolowicz et al., 2019). Similar results were obtained after treating animals with

11
carboplatin; DA and serotonin release were negatively affected while DA uptake was decreased in
the striatum, these changes that correlate to the impairment in spatial learning exhibited by the
carboplatin-treated animals (Kaplan et al., 2016).
Additionally, it has been found that intraperitoneal application of doxorubicin generates a decrease
of DA and its extra-neuronal metabolite 3-methoxytyramine (3-MT) in both the frontal cortex and
hippocampus of treated animals compared to the saline control (Antkiewicz-Michaluk et al., 2016).
In parallel, another research group documented that doxorubicin application produced a slower
glutamate uptake in the frontal cortex, while the dentate gyrus produced a slower clearance of
glutamate and consequently an overflow of glutamate in this region (Thomas et al., 2017). All this
together affected the swimming speed of the doxorubicin-treated group; however this effect lasted
only 24 hours after administration (Thomas et al., 2017).
Because of the above results, it stands out that the different chemotherapeutic agents are affecting
different neurotransmitter systems which in turn are affecting cognitive processes such as memory
and learning, common symptoms presented by patients with CICI. Therefore, neurotransmitter
balance can be considered as an essential mechanism in the development of the phenomenon.
3.1.5 Disturbance in glial cells
As mentioned before, glial cells are important for the CNS functioning. Particularly, astrocytes have
been reported as the physical and metabolic support of neurons (Kıray et al., 2016). However, it has
been described that astrocytes are involved in many other processes. In particular, astrocytes have
been shown to play a key role in cognitive processing by contributing to synaptic transmission and
structural plasticity, where a bidirectional communication between neurons and astrocytes is
established (Santello et al., 2019). Accordingly, it has also been documented that both astrocytes and
microglia play an important role in the formation and pruning of synapses. Also, even when the main
role of microglia is to phagocytize apoptotic neuronal bodies and some synapses (Vainchtein &
Molofsky, 2020), in recently years, it has been reported that microglia modulates the formation of
new neurons in the hippocampus, regulate dendritic and axonal growth, and stimulate the formation,
modulation and relocation of synapses, processes necessary for memory formation (Rodríguez-
Iglesias et al., 2019).
Effects of microglial activation after the intraperitoneal administration of cyclophosphamide have
been observed in the hippocampus of male rats that show increased in ED1-positive cells (a marker
of microglial activation) in response to treatment; also, this effect seem to be related to impaired

12
performance on novel place recognition (NORT) and contextual fear conditioning tests (Christie
et al., 2012).
In concurrence, in a male murine model it has been shown that intermittent intraperitoneal application
of docetaxel generates an increase in GFAP-positive astrocytes, indicating astrocyte activation. This
activation was seen at 48 hours after docetaxel administration and extended, at lower levels, up to 9
days after application. Furthermore, it was noted that docetaxel-treated animals performed worse in
the novel object recognition test (Fardell et al., 2014).
Also, oligodendrocyte survival result impaired in a dose-dependent after treatment of cells in culture
with cisplatin (Dietrich et al., 2006 reviewed in Stankovic et al., 2020). The same research group
reported similar results in an in vivo and in vitro models in which decreased oligodendrocyte survival
was evidenced by treatment with fluorouracil (Han et al., 2008).
Likewise, in a juvenile murine model it has been shown that peritoneal application of methotrexate
induces microglial activation, which in turn affects oligodendrocyte maturation dynamics and
increases astrocyte reactivity. Authors of these paper also reported that the reduction of microglia
restored tri-glial stability and cognitive function, assessed by the novel object recognition task
(Geraghty et al., 2019; Gibson et al., 2019). Importantly, other authors have also showed that
administration of doxorubicin induced astrocytic activation in the frontal cortex, striatum,
hippocampus, hypothalamus, and cerebellum. Finally, the authors suggested that these results
partially explained the cognitive impairment evidenced by a more unsatisfactory outcome in new
object recognition tests (Cardoso et al., 2020).
Taking together, these results reflect the enormous impact of chemotherapeutics on glial populations.
Particularly, the activation of astrocytes and microglia was evidenced. These is relevant since these
processes cause tissue damage due to the release of proinflammatory cytokines that in turn generate
neuroinflammation and ultimately neurodegeneration (Pekny et al., 2014).
3.1.6 Myelination loss
In the CNS, myelination is a complex and lifelong process in which oligodendrocytes wrap segments
of the axons of certain neurons. These segments become isolated and enable the saltatory action
potential, that is an evolutionary mechanism that had allowed faster conduction and processing of
information (Aggarwal et al., 2011; de Faria et al., 2019). Two types of myelination have been
described. The first is intrinsic myelination, which occurs independently of neuronal activity. The

13
second is adaptive myelination, which is regulated and shaped by neuronal activity and is associated
with neuroplasticity, critical for learning (Bechler et al., 2018).
In a murine model, methotrexate exposure has been documented to decrease the cortical expression
of BDNF, which together with TrkB, is required for adaptive myelination (Geraghty et al., 2019).
Interestingly, BDNF expression is normalized after microglia depletion, although the underlying
mechanisms are not yet known (Geraghty et al., 2019; Gibson et al., 2019). Also, toxic effects on
CNS progenitor cells and non-dividing oligodendrocytes has been reported with the use of
fluorouracil both in vivo and in vitro (Han et al., 2008). Administration of fluorouracil generates a
delayed damage in myelin and axons, which can be observed up to 56 days after the completion of
the treatment, resulting in a myelinopathy that did not correlate with chronic inflammation or vascular
damage caused by the chemotherapeutic (Han et al., 2008).
Similarly, it has been documented that the use of CMF regimen generates a decrease in myelin basic
protein (MBP) and O4 (a pro-oligodendrocyte marker), as well as a decrease in both, the area covered
by myelin and reduced thickness of the sheath in the corpus callosum of female rats (Briones &
Woods, 2014). This study, however, correlate neuroinflammation with loss of myelination and
subsequent cognitive impairment (Briones & Woods, 2014).
Overall, these results indicate that chemotherapeutics also can negatively affect the myelination
process. This damage appears to be persistent despite the cessation of chemotherapeutics. It would
partially explain why some patients report that chemobrain symptoms persist years after completing
cycles of chemotherapy.
3.1.7 Cytokine dysregulation
Cytokines are part of cellular communication, playing a fundamental role in the development and
physiology of the CNS; for these reasons CNS is especially vulnerable to dysregulated cytokine
network, and some changes in the microenvironment due to diseases or external factors.
Chemotherapy frequently lead to inflammation, and chronic inflammation can lead to tissue damage
and cell infiltration in the CNS. The mentioned phenomena intertwine, generating a vicious cycle of
inflammatory damage plus tissue damage, eventually leading to neurodegeneration (Becher et al.,
2017).
It has been documented that the use of cyclophosphamide in a murine model generates alterations
among cytokines. Within the proinflammatory cytokines interleukin-1 (IL-1), tumor necrosis

14
factor (TNF), and interleukin-6 (IL-6) were found to be significantly elevated in the hippocampus
and cortex, while the anti-inflammatory cytokine IL-10 was significantly decreased. In sum, this
imbalance induces a neuroinflammatory environment that contributes to cognitive impairment,
evidenced by increased immobility time in the forced swim test (FST) and decreased performance in
the latency retention transfer test observed in treated animals. These tests evaluate the extent of
behaviors associated with depression and memory acquisition and retention, respectively (Iqubal
et al., 2019).
In agreement, it has been reported that the application of cisplatin generates IL-1, and TNF,
elevation in the hippocampus of a murine model. Along with these results, it was also described that
treated animals performed worse in the NORT tests and in the motor coordination test (Jangra et al.,
2016). Similarly, other authors reported in another murine model that the application of cisplatin
caused a significant increase in the level of both TNF and IL-6 in the cerebral cortex of the treated
group (Arafa & Atteia, 2020). Likewise, it has been reported that intraperitoneal application of
fluorouracil in an aged murine model increased the levels of IL-1, IL-2, IL-3, IL-4, IL-5, and IL-17
in the hippocampus after one month of treatment, which was related to detrimental effects on mature
hippocampal neurons (Groves et al., 2017).
Furthermore, other researchers have described that chronic intraperitoneal application of paclitaxel
in a male murine model elevates levels of the proinflammatory cytokines TNF and IL-1, in the
hippocampus, and these results were related to the subsequent decrease in spatial memory of treated
animals by increasing escape latency times in the Morris Water Maze (MWM) test (Li et al., 2018).
Notably, treatment with the bleomycin, etoposide, and cisplatin (BEP) regimen in patients with
testicular cancer (TC) also produced a significant increase in TNF levels compared to the group that
also had TC but did not receive BEP treatment. In this study, elevated TNF levels were associated
with poorer cognitive performance (Amidi et al., 2017).
Moreover, administration of doxorubicin generates a dramatic increase in the levels of the
proinflammatory cytokines TNF and IL-17, while the level of the anti-inflammatory cytokine IL-
10 declines in the hippocampus of treated animals. Treated animals also showed a reduction in short-
term memory as measured by the spontaneous alternation test (SAP) (Wahdan et al., 2020).
Consistent with this, in another murine model in which DOX was applied intraperitoneally, an
overexpression of TNF levels was evidenced in the brain (Tabaczar et al., 2017). Similarly, in a
female OVX murine model, it was reported that the application of the regimen doxorubicin and

15
cyclophosphamide (AC) produced in the hippocampus a significant elevation of the cytokines IL-2,
IL-3, IL-4, IL-6, IL-10, IL-1ra, IL-17 and TNF (Bagnall-Moreau et al., 2019).
In addition, it has been reported that the use of the CMF regimen in an aged female murine model
generated increased levels of the proinflammatory cytokines TNF and IL-1b but reduced levels of
the anti-inflammatory cytokine IL-10 in the corpus callosum. Intriguingly, after one month of
cessation of treatment with the CMF regimen, the treated animals exhibited a worse performance in
memory and discrimination tests (Briones & Woods, 2014). Similar results were obtained with the
DAC regimen in which levels of the proinflammatory cytokines TNF and IL-6 were significantly
increased, but levels of the anti-inflammatory cytokine IL-4 and IL-10 were decreased in the whole
brain, especially in the prefrontal cortex and hippocampus of a female murine model (Shi et al., 2019).
Considering the evidence, chemotherapeutics promotes an imbalance between proinflammatory and
anti-inflammatory cytokines. Remarkable, it was documented that regardless of the chemotherapeutic
used, the cytokine frequently found to be elevated was TNF. This finding is important since it has
been described in the literature that the expression of this cytokine is related to increased oxidative
stress and mitochondrial dysfunction, which decreased in a TNF null model (Ren et al., 2019).
Additionally, it has also been documented that the elevation of reactive oxygen species (ROS) and
consequently of proinflammatory cytokines generate physical damage to the blood-brain barrier
(BBB), which subsequently contributes to the entry of small concentrations of chemotherapeutics that
normally do not have the capacity to pass the BBB, as is the case of Dox (Ren et al., 2019).
Finally, the described mechanisms show the different effects of chemotherapeutic agents that directly
or indirectly are affecting the communication or function of important brain regions involved in
cognition, such as the prefrontal cortex, hippocampus and striatum (Gonzalez-Burgos & Feriavelasco,
2008). For this reason, further research should continue to explore the short- and long-term effects of
chemotherapeutics, as this will also help in the development of therapeutic strategies.

16
Figure 2. Main neurobiological mechanisms involved in the development of Chemotherapy-Induced Cognitive
Impairment (CICI). The blue lines indicate mechanisms occurring in the prefrontal cortex, the green lines indicate
mechanisms occurring in the hippocampus, the orange line indicates processes occurring in the corpus callosum and the
red line indicates processes occurring in the cerebellum. Created with BioRender.com.
3.2 Future directions
Given that most of the original articles here presented reported results obtained in healthy murine
models, it is important to validate this results in cancer models, rarely contemplated, in order to dissect
cognitive impairment induced by chemotherapy from the effects induced by the disease itself.
In addition, during the review, it became evident that some murine models used only males or only
females or only ovariectomized females. In these cases, valuable information about the effect of
chemotherapeutics in the groups that are not under consideration could be missed. For this reason, it
would be enlightening if more controls could be considered within the experimental design in order
to have a more global view of the obtained results.

17
Conclusion
In conclusion, chemotherapy-induced cognitive impairment is a complex phenomenon mediated by
altered neurobiological mechanisms, among which neurotoxicity, decreased neurogenesis, loss of
spines and dendritic complexity, altered neurotransmission, disruption of glial cells, loss of
myelination and cytokine dysregulation leading to neuroinflammation were highlighted. It should be
made clear that it is still necessary to elucidate what other mechanisms are involved in the
development of chemobrain. Therefore, more research is needed, hopefully including cancer models
in their experimental design, to understand this problem globally.
Acknowledgements
I would especially like to thank my family, especially my parents for all the support and love they
have given me throughout my career and my life. I love you with all my heart.
To my director Sonia Luz Albarracin for all these years of dedication and for giving me the
opportunity to be part of such a special research group.
Acknowledgement to the Ecosistema Científico - Colombia Científica " Generación de alternativas
terapéuticas en cáncer a partir de plantas a partir de plantas a través de procesos de investigación y
desarrollo traslacional, articulados en sistemas de valor sostenibles ambiental y económicamente"
financed by Banco Mundial.
To my friends who have supported me so much in good and difficult moments, especially Andrés
Pascagaza, Jose Velasco, Viviana Vargas, and Juan Diego Tovar.
To incredible people like Ana Lucía Rangel, Laura Girón and Maria Alejandra Ariza for their great
support and affection during the last semesters.
To my friends since the first semester for so many good memories, Daniel Botiva y Klaus Lubinus.
To Jhon Sutachan and other members of the Neurobiochemistry laboratory from whom I have learned
so many things during all these years.
References
Aggarwal, S., Yurlova, L., & Simons, M. (2011). Central nervous system myelin: Structure, synthesis and
assembly. Trends in Cell Biology, 21(10), 585–593. https://doi.org/10.1016/j.tcb.2011.06.004

18
Alexander, T. C., Simecka, C. M., Kiffer, F., Groves, T., Anderson, J., Carr, H., Wang, J., Carter, G., &
Allen, A. R. (2018). Changes in cognition and dendritic complexity following intrathecal
methotrexate and cytarabine treatment in a juvenile murine model. Behavioural Brain Research, 346,
21–28. https://doi.org/10.1016/j.bbr.2017.12.008
Alhowail, A. H., Bloemer, J., Majrashi, M., Pinky, P. D., Bhattacharya, S., Yongli, Z., Bhattacharya, D.,
Eggert, M., Woodie, L., Buabeid, M. A., Johnson, N., Broadwater, A., Smith, B., Dhanasekaran, M.,
Arnold, R. D., & Suppiramaniam, V. (2019). Doxorubicin-induced neurotoxicity is associated with
acute alterations in synaptic plasticity, apoptosis, and lipid peroxidation. Toxicology Mechanisms and
Methods, 29(6), 457–466. https://doi.org/10.1080/15376516.2019.1600086
Ali, M. A., Menze, E. T., Tadros, M. G., & Tolba, M. F. (2020). Caffeic acid phenethyl ester counteracts
doxorubicin-induced chemobrain in Sprague-Dawley rats: Emphasis on the modulation of oxidative
stress and neuroinflammation. Neuropharmacology, 181, 108334.
https://doi.org/10.1016/j.neuropharm.2020.108334
Allen, B. D., Apodaca, L. A., Syage, A. R., Markarian, M., Baddour, A. A. D., Minasyan, H., Alikhani,
L., Lu, C., West, B. L., Giedzinski, E., Baulch, J. E., & Acharya, M. M. (2019). Attenuation of
neuroinflammation reverses Adriamycin-induced cognitive impairments. Acta Neuropathologica
Communications, 7(1), 186. https://doi.org/10.1186/s40478-019-0838-8
Almeida, D., Pinho, R., Correia, V., Soares, J., Bastos, M., Carvalho, F., Capela, J., & Costa, V. (2018).
Mitoxantrone is More Toxic than Doxorubicin in SH-SY5Y Human Cells: A ‘Chemobrain’ In Vitro
Study. Pharmaceuticals, 11(2), 41. https://doi.org/10.3390/ph11020041
American Cancer Society. (2020). Cancer Facts & Figures 2020 (p. 76). Atlanta: American Cancer
Society. https://www.cancer.org/content/dam/cancer-org/research/cancer-facts-and-statistics/annual-
cancer-facts-and-figures/2020/cancer-facts-and-figures-2020.pdf
Amidi, A., Agerbæk, M., Wu, L. M., Pedersen, A. D., Mehlsen, M., Clausen, C. R., Demontis, D.,
Børglum, A. D., Harbøll, A., & Zachariae, R. (2017). Changes in cognitive functions and cerebral
grey matter and their associations with inflammatory markers, endocrine markers, and APOE
genotypes in testicular cancer patients undergoing treatment. Brain Imaging and Behavior, 11(3),
769–783. https://doi.org/10.1007/s11682-016-9552-3
Anderson, J. E., Trujillo, M., McElroy, T., Groves, T., Alexander, T., Kiffer, F., & Allen, A. R. (2019).
Early Effects of Cyclophosphamide, Methotrexate, and 5-Fluorouracil on Neuronal Morphology and
Hippocampal-Dependent Behavior in a Murine Model. Toxicological Sciences, kfz213.
https://doi.org/10.1093/toxsci/kfz213
Andres, A. L., Gong, X., Di, K., & Bota, D. A. (2014). Low-doses of cisplatin injure hippocampal
synapses: A mechanism for ‘chemo’ brain? Experimental Neurology, 255, 137–144.
https://doi.org/10.1016/j.expneurol.2014.02.020
Antkiewicz-Michaluk, L., Krzemieniecki, K., Romanska, I., Michaluk, J., & Krygowska-Wajs, A. (2016).
Acute treatment with doxorubicin induced neurochemical impairment of the function of dopamine
system in rat brain structures. Pharmacological Reports, 68(3), 627–630.
https://doi.org/10.1016/j.pharep.2016.01.009
Arafa, M. H., & Atteia, H. H. (2020). Protective Role of Epigallocatechin Gallate in a Rat Model of
Cisplatin-Induced Cerebral Inflammation and Oxidative Damage: Impact of Modulating NF-κB and
Nrf2. Neurotoxicity Research, 37(2), 380–396. https://doi.org/10.1007/s12640-019-00095-x
Bagnall-Moreau, C., Chaudhry, S., Salas-Ramirez, K., Ahles, T., & Hubbard, K. (2019). Chemotherapy-
Induced Cognitive Impairment Is Associated with Increased Inflammation and Oxidative Damage in
the Hippocampus. Molecular Neurobiology, 56(10), 7159–7172. https://doi.org/10.1007/s12035-
019-1589-z

19
Banach, M., & Juranek, J. K. (2017). Chemobrain—A Troubling Side Effect of Chemotherapy. Journal
of Depression and Anxiety, 06(03). https://doi.org/10.4172/2167-1044.S11-002
Becher, B., Spath, S., & Goverman, J. (2017). Cytokine networks in neuroinflammation. Nature Reviews
Immunology, 17(1), 49–59. https://doi.org/10.1038/nri.2016.123
Bechler, M. E., Swire, M., & ffrench-Constant, C. (2018). Intrinsic and adaptive myelination-A sequential
mechanism for smart wiring in the brain: Intrinsic and Adaptive Myelination Mechanisms.
Developmental Neurobiology, 78(2), 68–79. https://doi.org/10.1002/dneu.22518
Bhattacharya, B., & Mukherjee, S. (2015). Cancer Therapy Using Antibiotics. Journal of Cancer Therapy,
06(10), 849–858. https://doi.org/10.4236/jct.2015.610093
Briones, T. L., & Woods, J. (2011). Chemotherapy-induced cognitive impairment is associated with
decreases in cell proliferation and histone modifications. BMC Neuroscience, 12(1), 124.
https://doi.org/10.1186/1471-2202-12-124
Briones, T. L., & Woods, J. (2014). Dysregulation in myelination mediated by persistent
neuroinflammation: Possible mechanisms in chemotherapy-related cognitive impairment. Brain,
Behavior, and Immunity, 35, 23–32. https://doi.org/10.1016/j.bbi.2013.07.175
Cardoso, C. V., de Barros, M. P., Bachi, A. L. L., Bernardi, M. M., Kirsten, T. B., de Fátima Monteiro
Martins, M., Rocha, P. R. D., da Silva Rodrigues, P., & Bondan, E. F. (2020). Chemobrain in rats:
Behavioral, morphological, oxidative and inflammatory effects of doxorubicin administration.
Behavioural Brain Research, 378, 112233. https://doi.org/10.1016/j.bbr.2019.112233
Castel, H., Denouel, A., Lange, M., Tonon, M.-C., Dubois, M., & Joly, F. (2017). Biomarkers Associated
with Cognitive Impairment in Treated Cancer Patients: Potential Predisposition and Risk Factors.
Frontiers in Pharmacology, 8. https://doi.org/10.3389/fphar.2017.00138
Cheung-Ong, K., Giaever, G., & Nislow, C. (2013). DNA-Damaging Agents in Cancer Chemotherapy:
Serendipity and Chemical Biology. Chemistry & Biology, 20(5), 648–659.
https://doi.org/10.1016/j.chembiol.2013.04.007
Chidambaram, S. B., Rathipriya, A. G., Bolla, S. R., Bhat, A., Ray, B., Mahalakshmi, A. M.,
Manivasagam, T., Thenmozhi, A. J., Essa, M. M., Guillemin, G. J., Chandra, R., & Sakharkar, M. K.
(2019). Dendritic spines: Revisiting the physiological role. Progress in Neuro-Psychopharmacology
and Biological Psychiatry, 92, 161–193. https://doi.org/10.1016/j.pnpbp.2019.01.005
Chiorazzi, A., Semperboni, S., & Marmiroli, P. (2015). Current View in Platinum Drug Mechanisms of
Peripheral Neurotoxicity. Toxics, 3(3), 304–321. https://doi.org/10.3390/toxics3030304
Chiu, G. S., Maj, M. A., Rizvi, S., Dantzer, R., Vichaya, E. G., Laumet, G., Kavelaars, A., & Heijnen, C.
J. (2017). Pifithrin-μ Prevents Cisplatin-Induced Chemobrain by Preserving Neuronal Mitochondrial
Function. Cancer Research, 77(3), 742–752. https://doi.org/10.1158/0008-5472.CAN-16-1817
Christie, L.-A., Acharya, M. M., Parihar, V. K., Nguyen, A., Martirosian, V., & Limoli, C. L. (2012).
Impaired Cognitive Function and Hippocampal Neurogenesis following Cancer Chemotherapy.
Clinical Cancer Research, 18(7), 1954–1965. https://doi.org/10.1158/1078-0432.CCR-11-2000
Dietrich, J., Han, R., Yang, Y., Mayer-Pröschel, M., & Noble, M. (2006). CNS progenitor cells and
oligodendrocytes are targets of chemotherapeutic agents in vitro and in vivo. Journal of Biology, 5(7),
22. https://doi.org/10.1186/jbiol50
English, K., Shepherd, A., Uzor, N.-E., Trinh, R., Kavelaars, A., & Heijnen, C. J. (2020). Astrocytes rescue
neuronal health after cisplatin treatment through mitochondrial transfer. Acta Neuropathologica
Communications, 8(1), 36. https://doi.org/10.1186/s40478-020-00897-7
Faber, D. S., & Pereda, A. E. (2018). Two Forms of Electrical Transmission Between Neurons. Frontiers
in Molecular Neuroscience, 11, 427. https://doi.org/10.3389/fnmol.2018.00427

20
Fardell, J. E., Zhang, J., De Souza, R., Vardy, J., Johnston, I., Allen, C., Henderson, J., & Piquette-Miller,
M. (2014). The impact of sustained and intermittent docetaxel chemotherapy regimens on cognition
and neural morphology in healthy mice. Psychopharmacology, 231(5), 841–852.
https://doi.org/10.1007/s00213-013-3301-8
Faria, O., Gonsalvez, D. G., Nicholson, M., & Xiao, J. (2019). Activity‐dependent central nervous system
myelination throughout life. Journal of Neurochemistry, 148(4), 447–461.
https://doi.org/10.1111/jnc.14592
Flanigan, T. J., Anderson, J. E., Elayan, I., Allen, A. R., & Ferguson, S. A. (2018). Effects of
Cyclophosphamide and/or Doxorubicin in a Murine Model of Postchemotherapy Cognitive
Impairment. Toxicological Sciences, 162(2), 462–474. https://doi.org/10.1093/toxsci/kfx267
Geraghty, A. C., Gibson, E. M., Ghanem, R. A., Greene, J. J., Ocampo, A., Goldstein, A. K., Ni, L., Yang,
T., Marton, R. M., Paşca, S. P., Greenberg, M. E., Longo, F. M., & Monje, M. (2019). Loss of
Adaptive Myelination Contributes to Methotrexate Chemotherapy-Related Cognitive Impairment.
Neuron, 103(2), 250-265.e8. https://doi.org/10.1016/j.neuron.2019.04.032
Gibson, E. M., Nagaraja, S., Ocampo, A., Tam, L. T., Wood, L. S., Pallegar, P. N., Greene, J. J., Geraghty,
A. C., Goldstein, A. K., Ni, L., Woo, P. J., Barres, B. A., Liddelow, S., Vogel, H., & Monje, M.
(2019). Methotrexate Chemotherapy Induces Persistent Tri-glial Dysregulation that Underlies
Chemotherapy-Related Cognitive Impairment. Cell, 176(1–2), 43-55.e13.
https://doi.org/10.1016/j.cell.2018.10.049
Gonzalez-Burgos, I., & Feriavelasco, A. (2008). Serotonin/dopamine interaction in memory formation. En
Progress in Brain Research (Vol. 172, pp. 603–623). Elsevier. https://doi.org/10.1016/S0079-
6123(08)00928-X
Groves, T. R., Farris, R., Anderson, J. E., Alexander, T. C., Kiffer, F., Carter, G., Wang, J., Boerma, M.,
& Allen, A. R. (2017). 5-Fluorouracil chemotherapy upregulates cytokines and alters hippocampal
dendritic complexity in aged mice. Behavioural Brain Research, 316, 215–224.
https://doi.org/10.1016/j.bbr.2016.08.039
Habli, Z., Toumieh, G., Fatfat, M., Rahal, O., & Gali-Muhtasib, H. (2017). Emerging Cytotoxic Alkaloids
in the Battle against Cancer: Overview of Molecular Mechanisms. Molecules, 22(2), 250.
https://doi.org/10.3390/molecules22020250
Han, R., Yang, Y. M., Dietrich, J., Luebke, A., Mayer-Pröschel, M., & Noble, M. (2008). Systemic 5-
fluorouracil treatment causes a syndrome of delayed myelin destruction in the central nervous system.
Journal of Biology, 7(4), 12. https://doi.org/10.1186/jbiol69
Hanahan, D., & Weinberg, R. A. (2011). Hallmarks of Cancer: The Next Generation. Cell, 144(5), 646–
674. https://doi.org/10.1016/j.cell.2011.02.013
Handra, C. (2019). THE CONNECTION BETWEEN DIFFERENT NEUROTRANSMITTERS
INVOLVED IN COGNITIVE PROCESSES. FARMACIA, 67(2), 193–201.
https://doi.org/10.31925/farmacia.2019.2.1
Huehnchen, P., Boehmerle, W., Springer, A., Freyer, D., & Endres, M. (2017). A novel preventive therapy
for paclitaxel-induced cognitive deficits: Preclinical evidence from C57BL/6 mice. Translational
Psychiatry, 7(8), e1185–e1185. https://doi.org/10.1038/tp.2017.149
Ikonomidou, C. (2018). Chemotherapy and the pediatric brain. Molecular and Cellular Pediatrics, 5(1),
8. https://doi.org/10.1186/s40348-018-0087-0
Iqubal, A., Sharma, S., Najmi, A. K., Syed, M. A., Ali, J., Alam, M. M., & Haque, S. E. (2019). Nerolidol
ameliorates cyclophosphamide-induced oxidative stress, neuroinflammation and cognitive
dysfunction: Plausible role of Nrf2 and NF- κB. Life Sciences, 236, 116867.
https://doi.org/10.1016/j.lfs.2019.116867

21
Jangra, A., Kwatra, M., Singh, T., Pant, R., Kushwah, P., Ahmed, S., Dwivedi, D., Saroha, B., & Lahkar,
M. (2016). Edaravone alleviates cisplatin-induced neurobehavioral deficits via modulation of
oxidative stress and inflammatory mediators in the rat hippocampus. European Journal of
Pharmacology, 791, 51–61. https://doi.org/10.1016/j.ejphar.2016.08.003
Jarmolowicz, D. P., Gehringer, R., Lemley, S. M., Sofis, M. J., Kaplan, S., & Johnson, M. A. (2019). 5-
Fluorouracil impairs attention and dopamine release in rats. Behavioural Brain Research, 362, 319–
322. https://doi.org/10.1016/j.bbr.2019.01.007
Ježek, J., Cooper, K. F., & Strich, R. (2021). The Impact of Mitochondrial Fission-Stimulated ROS
Production on Pro-Apoptotic Chemotherapy. Biology, 10(1), 33.
https://doi.org/10.3390/biology10010033
Joly, F., Giffard, B., Rigal, O., De Ruiter, M. B., Small, B. J., Dubois, M., LeFel, J., Schagen, S. B., Ahles,
T. A., Wefel, J. S., Vardy, J. L., Pancré, V., Lange, M., & Castel, H. (2015). Impact of Cancer and Its
Treatments on Cognitive Function: Advances in Research From the Paris International Cognition and
Cancer Task Force Symposium and Update Since 2012. Journal of Pain and Symptom Management,
50(6), 830–841. https://doi.org/10.1016/j.jpainsymman.2015.06.019
Kaplan, S. V., Limbocker, R. A., Gehringer, R. C., Divis, J. L., Osterhaus, G. L., Newby, M. D., Sofis, M.
J., Jarmolowicz, D. P., Newman, B. D., Mathews, T. A., & Johnson, M. A. (2016). Impaired Brain
Dopamine and Serotonin Release and Uptake in Wistar Rats Following Treatment with Carboplatin.
ACS Chemical Neuroscience, 7(6), 689–699. https://doi.org/10.1021/acschemneuro.5b00029
Kärkkäinen, V., Pomeshchik, Y., Savchenko, E., Dhungana, H., Kurronen, A., Lehtonen, S., Naumenko,
N., Tavi, P., Levonen, A.-L., Yamamoto, M., Malm, T., Magga, J., Kanninen, K. M., & Koistinaho,
J. (2014). Nrf2 Regulates Neurogenesis and Protects Neural Progenitor Cells Against Aβ Toxicity:
Regulation and Protection of NPCs by Nrf2. STEM CELLS, 32(7), 1904–1916.
https://doi.org/10.1002/stem.1666
Kasai, H., Fukuda, M., Watanabe, S., Hayashi-Takagi, A., & Noguchi, J. (2010). Structural dynamics of
dendritic spines in memory and cognition. Trends in Neurosciences, 33(3), 121–129.
https://doi.org/10.1016/j.tins.2010.01.001
Kempermann, G., Song, H., & Gage, F. H. (2015). Neurogenesis in the Adult Hippocampus. Cold Spring
Harbor Perspectives in Biology, 7(9), a018812. https://doi.org/10.1101/cshperspect.a018812
Kıray, H., Lindsay, S. L., Hosseinzadeh, S., & Barnett, S. C. (2016). The multifaceted role of astrocytes
in regulating myelination. Experimental Neurology, 283, 541–549.
https://doi.org/10.1016/j.expneurol.2016.03.009
Koppelmans, V., Breteler, M. M. B., Boogerd, W., Seynaeve, C., Gundy, C., & Schagen, S. B. (2012).
Neuropsychological Performance in Survivors of Breast Cancer More Than 20 Years After Adjuvant
Chemotherapy. Journal of Clinical Oncology, 30(10), 1080–1086.
https://doi.org/10.1200/JCO.2011.37.0189
Kovalchuk, A., Ilnytskyy, Y., Woycicki, R., Rodriguez-Juarez, R., Metz, G. A. S., & Kovalchuk, O.
(2018). Adverse effects of paternal chemotherapy exposure on the progeny brain: Intergenerational
chemobrain. Oncotarget, 9(11), 10069–10082. https://doi.org/10.18632/oncotarget.24311
Kwatra, M., Jangra, A., Mishra, M., Sharma, Y., Ahmed, S., Ghosh, P., Kumar, V., Vohora, D., & Khanam,
R. (2016). Naringin and Sertraline Ameliorate Doxorubicin-Induced Behavioral Deficits Through
Modulation of Serotonin Level and Mitochondrial Complexes Protection Pathway in Rat
Hippocampus. Neurochemical Research, 41(9), 2352–2366. https://doi.org/10.1007/s11064-016-
1949-2
Levin, E. D. (Ed.). (2006). Neurotransmitter interactions and cognitive function. Birkhäuser Verlag.

22
Li, Z., Zhao, S., Zhang, H.-L., Liu, P., Liu, F.-F., Guo, Y.-X., & Wang, X.-L. (2018). Proinflammatory
Factors Mediate Paclitaxel-Induced Impairment of Learning and Memory. Mediators of
Inflammation, 2018, 1–9. https://doi.org/10.1155/2018/3941840
Liao, D., Xiang, D., Dang, R., Xu, P., Wang, J., Han, W., Fu, Y., Yao, D., Cao, L., & Jiang, P. (2018).
Neuroprotective Effects of dl-3-n-Butylphthalide against Doxorubicin-Induced Neuroinflammation,
Oxidative Stress, Endoplasmic Reticulum Stress, and Behavioral Changes. Oxidative Medicine and
Cellular Longevity, 2018, 1–13. https://doi.org/10.1155/2018/9125601
Liberati, A., Altman, D. G., Tetzlaff, J., Mulrow, C., Gøtzsche, P. C., Ioannidis, J. P. A., Clarke, M.,
Devereaux, P. J., Kleijnen, J., & Moher, D. (2009). The PRISMA statement for reporting systematic
reviews and meta-analyses of studies that evaluate health care interventions: Explanation and
elaboration. Journal of Clinical Epidemiology, 62(10), e1–e34.
https://doi.org/10.1016/j.jclinepi.2009.06.006
Magge, R. S., & DeAngelis, L. M. (2015). The double-edged sword: Neurotoxicity of chemotherapy.
Blood Reviews, 29(2), 93–100. https://doi.org/10.1016/j.blre.2014.09.012
Meléndez, D. M., Nordquist, R. E., Vanderschuren, L. J. M. J., & van der Staay, F.-J. (2020). Spatial
memory deficits after vincristine-induced lesions to the dorsal hippocampus. PLOS ONE, 15(4),
e0231941. https://doi.org/10.1371/journal.pone.0231941
Moore, K., Stutzman, S., Priddy, L., & Olson, D. (2019). Chemobrain: A Pilot Study Exploring the
Severity and Onset of Chemotherapy-Related Cognitive Impairment. Clinical Journal of Oncology
Nursing, 23(4), 411–416. https://doi.org/10.1188/19.CJON.411-416
Mustafa, S., Walker, A., Bennett, G., & Wigmore, P. M. (2008). 5-Fluorouracil chemotherapy affects
spatial working memory and newborn neurons in the adult rat hippocampus. European Journal of
Neuroscience, 28(2), 323–330. https://doi.org/10.1111/j.1460-9568.2008.06325.x
Myers, J. S., Pierce, J., & Pazdernik, T. (2008). Neurotoxicology of Chemotherapy in Relation to Cytokine
Release, the Blood-Brain Barrier, and Cognitive Impairment. Oncology Nursing Forum, 35(6), 916–
920. https://doi.org/10.1188/08.ONF.916-920
Nguyen, L. D., & Ehrlich, B. E. (2020a). Cellular mechanisms and treatments for chemobrain: Insight
from aging and neurodegenerative diseases. EMBO Molecular Medicine, 12(6).
https://doi.org/10.15252/emmm.202012075
Nguyen, L. D., & Ehrlich, B. E. (2020b). Cellular mechanisms and treatments for chemobrain: Insight
from aging and neurodegenerative diseases. EMBO Molecular Medicine, 12(6).
https://doi.org/10.15252/emmm.202012075
Pekny, M., Wilhelmsson, U., & Pekna, M. (2014). The dual role of astrocyte activation and reactive gliosis.
Neuroscience Letters, 565, 30–38. https://doi.org/10.1016/j.neulet.2013.12.071
Pellacani, C., & Costa, L. G. (2018). Role of autophagy in environmental neurotoxicity. Environmental
Pollution, 235, 791–805. https://doi.org/10.1016/j.envpol.2017.12.102
Pellacani, C., & Eleftheriou, G. (2020). Neurotoxicity of antineoplastic drugs: Mechanisms, susceptibility,
and neuroprotective strategies. Advances in Medical Sciences, 65(2), 265–285.
https://doi.org/10.1016/j.advms.2020.04.001
Piccolini, V. M., Esposito, A., Dal Bo, V., Insolia, V., Bottone, M. G., De Pascali, S. A., Fanizzi, F. P., &
Bernocchi, G. (2015). Cerebellum neurotransmission during postnatal development: [Pt( O , O ′‐
acac)(γ‐acac)(DMS)] vs cisplatin and neurotoxicity. International Journal of Developmental
Neuroscience, 40(1), 24–34. https://doi.org/10.1016/j.ijdevneu.2014.10.006
Ren, X., Boriero, D., Chaiswing, L., Bondada, S., St. Clair, D. K., & Butterfield, D. A. (2019). Plausible
biochemical mechanisms of chemotherapy-induced cognitive impairment (“chemobrain”), a
condition that significantly impairs the quality of life of many cancer survivors. Biochimica et

23
Biophysica Acta (BBA) - Molecular Basis of Disease, 1865(6), 1088–1097.
https://doi.org/10.1016/j.bbadis.2019.02.007
Ren, X., Keeney, J. T. R., Miriyala, S., Noel, T., Powell, D. K., Chaiswing, L., Bondada, S., St. Clair, D.
K., & Butterfield, D. A. (2019). The triangle of death of neurons: Oxidative damage, mitochondrial
dysfunction, and loss of choline-containing biomolecules in brains of mice treated with doxorubicin.
Advanced insights into mechanisms of chemotherapy induced cognitive impairment (“chemobrain”)
involving TNF-α. Free Radical Biology and Medicine, 134, 1–8.
https://doi.org/10.1016/j.freeradbiomed.2018.12.029
Rodríguez-Iglesias, N., Sierra, A., & Valero, J. (2019). Rewiring of Memory Circuits: Connecting Adult
Newborn Neurons With the Help of Microglia. Frontiers in Cell and Developmental Biology, 7, 24.
https://doi.org/10.3389/fcell.2019.00024
Santello, M., Toni, N., & Volterra, A. (2019). Astrocyte function from information processing to cognition
and cognitive impairment. Nature Neuroscience, 22(2), 154–166. https://doi.org/10.1038/s41593-
018-0325-8
Shaker, F. H., El-Derany, M. O., Wahdan, S. A., El-Demerdash, E., & El-Mesallamy, H. O. (2021).
Berberine ameliorates doxorubicin-induced cognitive impairment (chemobrain) in rats. Life Sciences,
269, 119078. https://doi.org/10.1016/j.lfs.2021.119078
Shi, D.-D., Huang, Y.-H., Lai, C. S. W., Dong, C. M., Ho, L. C., Wu, E. X., Li, Q., Wang, X.-M., Chung,
S. K., Sham, P. C., & Zhang, Z.-J. (2019). Chemotherapy-Induced Cognitive Impairment Is
Associated with Cytokine Dysregulation and Disruptions in Neuroplasticity. Molecular
Neurobiology, 56(3), 2234–2243. https://doi.org/10.1007/s12035-018-1224-4
Singh, S., & Kumar, A. (2019). Protective Effect of Edaravone on Cyclophosphamide Induced Oxidative
Stress and Neurotoxicity in Rats. Current Drug Safety, 14(3), 209–216.
https://doi.org/10.2174/1574886314666190506100717
Snyder, J. S. (2019). Recalibrating the Relevance of Adult Neurogenesis. Trends in Neurosciences, 42(3),
164–178. https://doi.org/10.1016/j.tins.2018.12.001
Stankovic, J. S. K., Selakovic, D., Mihailovic, V., & Rosic, G. (2020). Antioxidant Supplementation in
the Treatment of Neurotoxicity Induced by Platinum-Based Chemotherapeutics—A Review.
International Journal of Molecular Sciences, 21(20), 7753. https://doi.org/10.3390/ijms21207753
Tabaczar, S., Czepas, J., Koceva, A., Kilanczyk, E., Piasecka-Zelga, J., & Gwozdzinski, K. (2017). The
effect of the nitroxide pirolin on oxidative stress induced by doxorubicin and taxanes in the rat brain.
Journal of Physiology and Pharmacology, 68, 295–308.
Tacar, O., Sriamornsak, P., & Dass, C. R. (2012). Doxorubicin: An update on anticancer molecular action,
toxicity and novel drug delivery systems. Journal of Pharmacy and Pharmacology, 65(2), 157–170.
https://doi.org/10.1111/j.2042-7158.2012.01567.x
Tau, G. Z., & Peterson, B. S. (2010). Normal Development of Brain Circuits. Neuropsychopharmacology,
35(1), 147–168. https://doi.org/10.1038/npp.2009.115
Thomas, T. C., Beitchman, J. A., Pomerleau, F., Noel, T., Jungsuwadee, P., Butterfield, D. A., Clair, D.
K. St., Vore, M., & Gerhardt, G. A. (2017). Acute treatment with doxorubicin affects glutamate
neurotransmission in the mouse frontal cortex and hippocampus. Brain Research, 1672, 10–17.
https://doi.org/10.1016/j.brainres.2017.07.003
Vainchtein, I. D., & Molofsky, A. V. (2020). Astrocytes and Microglia: In Sickness and in Health. Trends
in Neurosciences, 43(3), 144–154. https://doi.org/10.1016/j.tins.2020.01.003
Wahdan, S. A., El-Derany, M. O., Abdel-Maged, A. E., & Azab, S. S. (2020). Abrogating doxorubicin-
induced chemobrain by immunomodulators IFN-beta 1a or infliximab: Insights to neuroimmune

24
mechanistic hallmarks. Neurochemistry International, 138, 104777.
https://doi.org/10.1016/j.neuint.2020.104777
Wang, C., Zhao, Y., Wang, L., Pan, S., Liu, Y., Li, S., & Wang, D. (2021). C-phycocyanin Mitigates
Cognitive Impairment in Doxorubicin-Induced Chemobrain: Impact on Neuroinflammation,
Oxidative Stress, and Brain Mitochondrial and Synaptic Alterations. Neurochemical Research, 46(2),
149–158. https://doi.org/10.1007/s11064-020-03164-2
Welbat, J. U., Naewla, S., Pannangrong, W., Sirichoat, A., Aranarochana, A., & Wigmore, P. (2020).
Neuroprotective effects of hesperidin against methotrexate-induced changes in neurogenesis and
oxidative stress in the adult rat. Biochemical Pharmacology, 178, 114083.
https://doi.org/10.1016/j.bcp.2020.114083
World Health Organization. (2020). All cancers. International Agency for Research on Cancer.


















