
An exploratory osteological study of the muscovy duck Cairina...
15
An exploratory osteological study of the muscovy duck (Cairina moschata) (Aves: Anatidae) with implications for neotropical archaeology Peter W. Stahl * Department of Anthropology, Binghamton University, Binghamton, NY, 13902 USA Received 21 September 2004; received in revised form 6 January 2005 Abstract The muscovy duck (Cairina moschata) (Aves: Anatidae) is the most enigmatic of the native New World animal domesticates. Although large and conspicuous, this duck is nonetheless rarely recovered in archaeological context. When identified, its domesticated status remains uncertain for lack of distinguishing criteria used to separate male from female and domestic from wild specimens. This study offers provisional criteria for osteological discrimination based upon an analysis of 34 museum skeletons. It applies these data to archaeological specimens recovered from the western lowlands of Ecuador, where muscovy duck, along with other domesticated animals may appear in contexts associated with prehispanic trade. Ó 2005 Elsevier Ltd. All rights reserved. Keywords: Muscovy duck; Cairina moschata; Animal domestication; South America; Archaeology 1. Introduction Compared to other parts of the world, few animals were originally domesticated in South America. In- digenous domesticates include: two closely related camelids, the llama (Lama glama) and alpaca (Vicugna pacos); a medium-sized rodent, the guinea pig or cuy (Cavia porcellus) [17]; and, a large duck, the muscovy (Cairina moschata). Domesticated cuy and camelids continue to be important for contemporary indigenous populations, and we know more about their early domestication and later prehispanic distribution. How- ever, these aspects of the muscovy duck are virtually unknown. In addition to a host of indigenous names, it is presently referred to in Latin America as the pato real (royal), criollo (native), machaco´n (heavy), manso (tame), mudo (mute), casero (house), negro (black), almizclado/almizcarado (musky; from Latin moschatus), perulero (peruvian; also Nahuatl peruto´to´l), bravo (wild), and do mato (forest). Both its common English and generic Latin appellations underscore the enigma that is the muscovy duck. The duck’s ancestry is connected neither with Moscow (muscovy) nor with Cairo (Cairina) [13,20,28]; rather, it is indigenous to the lowland neotropics where it may have been domesticated originally. A very large forest duck, the muscovy is taxonom- ically included with the tropical to subtropical perching ducks (Tribe: Cairinini). The Cairinini are large anatids characterized by long tails, which they use as brakes for landing in trees where their sharp claws enable them to both roost and nest in hollows from 3 to 20 m above ground [21]. In the wild, the muscovy is widely distributed along both coasts of Mexico, from Sinaloa and Tamaulipas in the west and Nuevo Leo´ n in the east. * Tel.: C1 607 777 2557; fax: C1 607 777 2477. E-mail address: [email protected] 0305-4403/$ - see front matter Ó 2005 Elsevier Ltd. All rights reserved. doi:10.1016/j.jas.2005.01.009 Journal of Archaeological Science 32 (2005) 915e929 http://www.elsevier.com/locate/jas
Transcript of An exploratory osteological study of the muscovy duck Cairina...

Journal of Archaeological Science 32 (2005) 915e929
http://www.elsevier.com/locate/jas
An exploratory osteological study of the muscovy duck(Cairina moschata) (Aves: Anatidae) with implications
for neotropical archaeology
Peter W. Stahl*
Department of Anthropology, Binghamton University, Binghamton, NY, 13902 USA
Received 21 September 2004; received in revised form 6 January 2005
Abstract
The muscovy duck (Cairina moschata) (Aves: Anatidae) is the most enigmatic of the native New World animal domesticates.Although large and conspicuous, this duck is nonetheless rarely recovered in archaeological context. When identified, its
domesticated status remains uncertain for lack of distinguishing criteria used to separate male from female and domestic from wildspecimens. This study offers provisional criteria for osteological discrimination based upon an analysis of 34 museum skeletons. Itapplies these data to archaeological specimens recovered from the western lowlands of Ecuador, where muscovy duck, along with
other domesticated animals may appear in contexts associated with prehispanic trade.� 2005 Elsevier Ltd. All rights reserved.
Keywords: Muscovy duck; Cairina moschata; Animal domestication; South America; Archaeology
1. Introduction
Compared to other parts of the world, few animalswere originally domesticated in South America. In-digenous domesticates include: two closely relatedcamelids, the llama (Lama glama) and alpaca (Vicugnapacos); a medium-sized rodent, the guinea pig or cuy(Cavia porcellus) [17]; and, a large duck, the muscovy(Cairina moschata). Domesticated cuy and camelidscontinue to be important for contemporary indigenouspopulations, and we know more about their earlydomestication and later prehispanic distribution. How-ever, these aspects of the muscovy duck are virtuallyunknown.
In addition to a host of indigenous names, it ispresently referred to in Latin America as the pato real
* Tel.: C1 607 777 2557; fax: C1 607 777 2477.
E-mail address: [email protected]
0305-4403/$ - see front matter � 2005 Elsevier Ltd. All rights reserved.
doi:10.1016/j.jas.2005.01.009
(royal), criollo (native), machacon (heavy), manso(tame), mudo (mute), casero (house), negro (black),almizclado/almizcarado (musky; from Latin moschatus),perulero (peruvian; also Nahuatl perutotol), bravo(wild), and do mato (forest). Both its common Englishand generic Latin appellations underscore the enigmathat is the muscovy duck. The duck’s ancestry isconnected neither with Moscow (muscovy) nor withCairo (Cairina) [13,20,28]; rather, it is indigenous to thelowland neotropics where it may have been domesticatedoriginally.
A very large forest duck, the muscovy is taxonom-ically included with the tropical to subtropical perchingducks (Tribe: Cairinini). The Cairinini are large anatidscharacterized by long tails, which they use as brakes forlanding in trees where their sharp claws enable them toboth roost and nest in hollows from 3 to 20 m aboveground [21]. In the wild, the muscovy is widelydistributed along both coasts of Mexico, from Sinaloaand Tamaulipas in the west and Nuevo Leon in the east.

916 P.W. Stahl / Journal of Archaeological Science 32 (2005) 915e929
Its range continues southward through Central Americainto forested lowland areas of South America where themuscovy has been recorded as far south as Peru to thewest of the Andes, and northern Argentina andUruguay to the east. In the Caribbean, there is noevidence of wild populations north of Trinidad. Wildforms are grayish black ventrally and blackish withgreen and purple iridescence on the dorsal side, alongwith prominent white patches on their wing coverts.Female ducks have feathered faces, whereas drakesfeature bare faces with prominent occipital and nuchalcrests and small fleshy caruncles extending from aroundthe orbital region to the base of the bill. Most notably,males are much larger than, and often double the weightof females, with little overlap between the sexes.Although known to gather in groups of over 50 duringbreeding, muscovy ducks usually fly crepuscularly insmall groups or alone. Very territorial, they prefer banksalong streams and swamps, and forage in savannasduring the wet season, yet are never far from trees wherethey spend the night between palm fronds or in treeholes. A highly omnivorous and opportunistic eater, themuscovy is known to consume small fish and reptiles,invertebrates, crustaceans, insects and especially ter-mites, water plants and seeds, and agricultural crops[3,13,21,27,28,40,44].
Today domesticated and feral muscovies are distrib-uted worldwide. Shortly after Europeans arrived in theNew World, domesticated muscovies were brought backto Europe and subsequently introduced into Africa, Asia,and Australia. Domestic poultry production prizes themuscovy for its hardiness and lean meat which containslower fat content than its Asian domestic counterpart,the domestic mallard or Asian pekin duck (Anasplatyrhynchos). Known to copulate freely with otherducks, promiscuous muscovy drakes are principally usedto produce large hybrids, increase vigor, and improve eggproduction. Current interest in domesticated muscovyducks is centered in France where they are crossed withthe Asian pekin duck to produce sterile hybrid mule ormulard ducks for meat and pate de foie gras. Domesti-cation produces greater variation in plumage andsignificant size increase. Domesticated muscovies tendtowards albinism, or pied black and white coloration,and feature larger red caruncles. Most impressive isa notable increase in overall weight and size. Under idealconditions, drakes attain weights of up to 6 kg comparedto 4 kg for wild forms, whereas, female ducks can increasetheir maximum weight from 1.5 kg in the wild to over2.5 kg under domestication [5,13,21,28,29,42].
Certain behavioral and ecological characteristics ofthe muscovy duck would in theory favor its domestica-tion by humans. They can be territorial and gregarious,have no fixed breeding season, and tend toward prolificegg production with rapidly maturing precocial hatch-lings. Requiring relatively little care, they are susceptible
to less contagious diseases, grow to a large size, and arequite capable of protecting themselves from predators.Omnivorous wild muscovies molest crops, and domes-ticates are often used in household scavenging or forcleaning open drains and irrigation ditches [1,13,15,16,42]. Many early European accounts describedomesticated muscovy ducks around the neotropics(e.g., [1,7:13]; however, we know very little about theirprehispanic domestication. Muscovy duck specimens areonly rarely identified in zooarchaeological bone sam-ples, a deficiency that is exacerbated by even limitedskeletal description, including relevant data for discrim-inating males from females, and wild from domesticbirds. This paper describes an exploratory study thatprovides pertinent skeletal data for archaeologicalapplication and examines recently identified specimensfrom the neotropical lowlands of Ecuador as a pre-liminary test.
2. Materials and methods
I studied 34 vouchered skeletons currently housed inmajor research collections throughout the United States(Table 1). Unfortunately a few specimens are repre-sented by only partial skeletons, and some lack crucialsupporting data, i.e. sex and domesticated status. Manyspecimens are simply labeled ‘‘captive’’, a term that doesnot elucidate whether the bird was wild or bred incaptivity. Specimens obtained outside of the muscovy’swild range could be culled from feral populations. Inthe event that captive specimens are indeed feral, then thesample minimally includes only five wild specimens.The following osteometric study includes only adultspecimens; however, two juvenile skeletons were alsoexamined.
Each skeleton was measured for up to 64 standard-ized cranial and post cranial measurements described byvon den Driesch [37]. For sided elements, the left waschosen but substituted by its right counterpart in casesof missing or damaged specimens. In most instances,measurement was recorded with digital calipers to 0.00mm, while certain specimens approaching linear dimen-sions of 150 mm and more were measured with large dialcalipers. All measurements recorded in this paper areaveraged values of three separate measurements un-dertaken for each variable. Measurement error rangedbetween G0.05 mm. During measurement, each skeletalelement was also visually examined for qualitativeattributes that might have potential relevance forosteologically discriminating sex or domesticated status.All quantitative data were entered into SPSS ver. 11.5for statistical analysis.
Osteometric data for 32 adult specimens were firstexplored through principal components analysis (PCA),which can be used to identify the major variables that
917P.W. Stahl / Journal of Archaeological Science 32 (2005) 915e929
Table 1
Analyzed muscovy duck (Cairina moschata) specimens
Specimen Identification Sex Status Age Location
AMNH 1848 _ Captive Adult Central Park Zoo NY, USA
AMNH 3004 _ ‘‘Domesticated’’ Adult ?
AMNH 26184 ? Captive Adult Sea World, San Diego CA, USA
AMNH 1321 ? ? Adult Bronx Park, NY, USA
AMNH 26185 \ Captive Adult Sea World, San Diego CA, USA
AMNH 4251 ? Captive Adult Central Park Zoo NY, USA
AMNH 1850 ? Captive Adult Central Park Zoo NY, USA
USNM 622299 _ Wild Adult Wiwithau Mountain, Guyana
AMNH 675 _ ? Adult Central Park Zoo NY, USA
AMNH 11024 \ ? Adult Cider Mill Farm, Huntington NY, USA
CM 4991 ? ? Adult Level Green, Westmoreland County PA, USA
CM 13803 \ ? Adult Pinellas County FL, USA
CM 8265 ? ? Adult Between Pittsburgh and Uniontown PA, USA
FMNH 338938 _ Captive Adult St. Petersburg, Pinellas County FL, USA
FMNH 338937 _ ? Adult Bell, Gilchrist County FL, USA
FMNH 104719 \ ? Adult South America (cage)
FMNH 338936 \ Domesticated Adult Gainesville, Alachua County FL, USA
FMNH 338934 \ Captive Adult Tampa, Hillsborough County FL, USA
FLMNH 23357 _ Captive/Feral Adult Gainesville, FL USA
USNM 19263 ? Captive Adult Monserrat., held at National Zoo, Washington
D.C., USA
USNM 15777 ? Domesticated Adult Washington D.C., USA
CU 43137 ? ? Adult ?
FLMNH 23356 _ Captive/Feral Adult Gainesville FL, USA
FMNH 338935 \ Captive Adult The Wildfowl Trust, UK
FMNH 23354 _ Captive/Feral Juvenile Suncoast Seabird Sanctuary, St. Petersburg
FL, USA
USNM 224000 ? Captive Adult ?, zoo animal
UMMZ 133626 _ Wild Adult Guanacaste, Costa Rica
UMMZ 133627 \ Wild Adult Guanacaste, Costa Rica
KU 34661 \ Wild Adult El Astillero, Guatemala
MVZ 132911 \ Captive Adult San Diego Zoo CA, USA
MVZ 85522 ? Wild Adult El Salvador
LACM 35401 _ ? Juvenile Tarija, Capirenda Bolivia
AARF 1 _ Domestic Adult Ferndale NY, USA
AARF 2 \ Domestic Adult Ferndale NY, USA
AMNH, American Museum of Natural History; AARF, Archaeological Analytical Research Facility, Binghamton University; CM, Carnegie
Museum of Natural History; CU, Cornell University; FMNH, Field Museum of Natural History; FLMNH, Florida Museum of Natural History;
KU, University of Kansas Museum of Natural History; LACM, Los Angeles County Museum; MVZ, Museum of Vertebrate Zoology, University of
California, Berkeley; UMMZ, University of Michigan Museum of Zoology; USNM, Smithsonian Institution, National Museum of Natural History.
explain much of the variance within a sample. Sub-sequent follow-up examination used multiple (MAN-OVA) and univariate (ANOVA) analyses of variance,which test the null hypothesis that group means areequal in a population by comparing sample variancethat is estimated from mean values. In addition,bivariate scatter plots for 64 metric variables wereinspected, along with detailed visual examination ofeach skeletal element.
3. Results
For PCA, an unrotated solution was extracted usinglogarithmically transformed data and a covariancematrix. Together, the first two principal componentsaccount for almost 84% of total variance in the sample.The scores on unrotated loadings for principal compo-
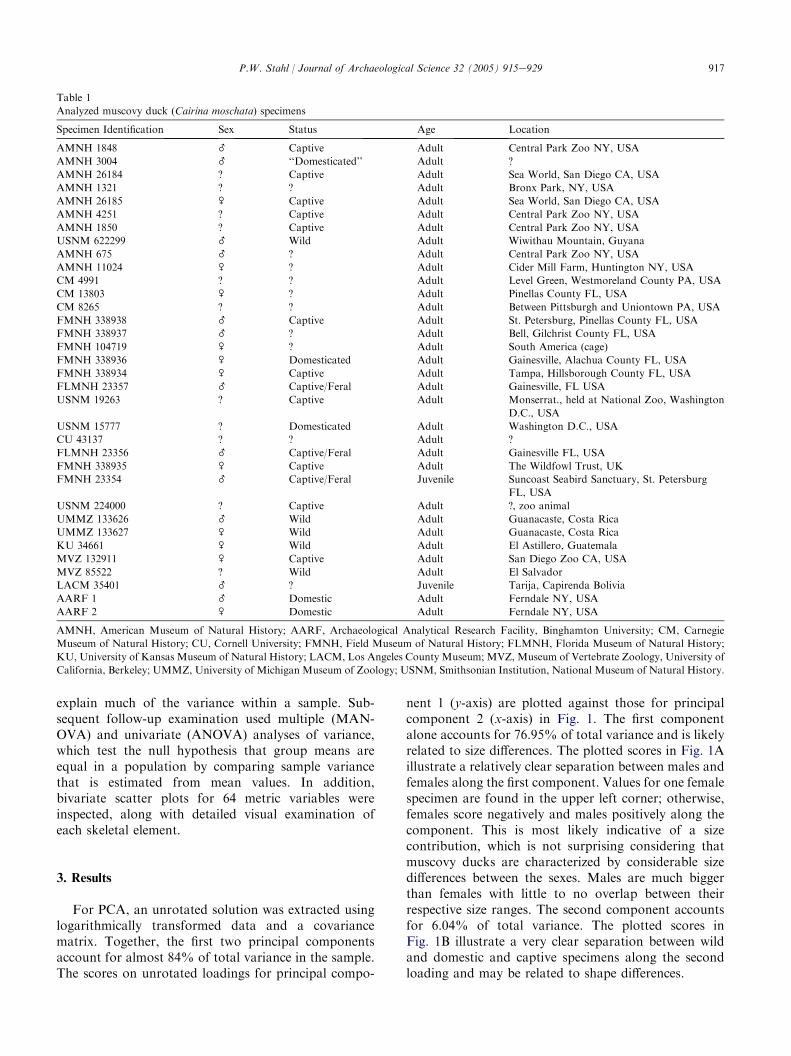
nent 1 (y-axis) are plotted against those for principalcomponent 2 (x-axis) in Fig. 1. The first componentalone accounts for 76.95% of total variance and is likelyrelated to size differences. The plotted scores in Fig. 1Aillustrate a relatively clear separation between males andfemales along the first component. Values for one femalespecimen are found in the upper left corner; otherwise,females score negatively and males positively along thecomponent. This is most likely indicative of a sizecontribution, which is not surprising considering thatmuscovy ducks are characterized by considerable sizedifferences between the sexes. Males are much biggerthan females with little to no overlap between theirrespective size ranges. The second component accountsfor 6.04% of total variance. The plotted scores inFig. 1B illustrate a very clear separation between wildand domestic and captive specimens along the secondloading and may be related to shape differences.
918 P.W. Stahl / Journal of Archaeological Science 32 (2005) 915e929
Loadings along the two principal components arelisted by variable in Appendix A. With one exception, allloadings along the first component are high andpositive. Highest positive values (O0.09) are dominatedby greatest length measurements, and are particularlyprevalent among limb variables. These results likelyrepresent size differences. Highest positive values (O0.3)along the second component include a cluster of cranialvariables and a few distal limb measurements. Somemay be indicative of shape differences. Size and shapeare examined in detail below, with emphasis on thosevariables that might be important for discriminatingmale from female, and/or wild from domestic specimens.
3.1. Osteometric discrimination of size
Statistical significance associated with trends sug-gested in the PCA loadings were further explored using
Fig. 1. (A) Scores on unrotated loadings for principal component 1 (y-
axis) and principal component 2 (x-axis), classified by sex. (B) Scores
on unrotated loadings for principal component 1 (y-axis) and principal
component 2 (x-axis), classified by wild/domestic/captive status.
analyses of variance. Initial use of MANOVA clearlyindicated statistically significant differences betweenmale and female specimens; however, follow-up analysesemployed univariate ANOVA in order to use as much ofthe data as possible. MANOVA rejects data for anentire specimen in cases where any individual value ismissing; therefore, by separately examining each vari-able, a maximum number of specimens were availablefor analysis. With sex defined as the fixed factor,separate measurements were individually compared asdependent variables. Each post hoc univariate ANOVAtest employed Tukey, Bonferoni, or R-E-G-W Q (Ryan-Einot-Gabriel-Welsch range) tests to evaluate pair-wisedifferences among means where equal variances wereassumed, and Dunnett’s C procedure where equalvariances were not assumed. A test of homogeneity ofvariance was used for choosing between assumptions.
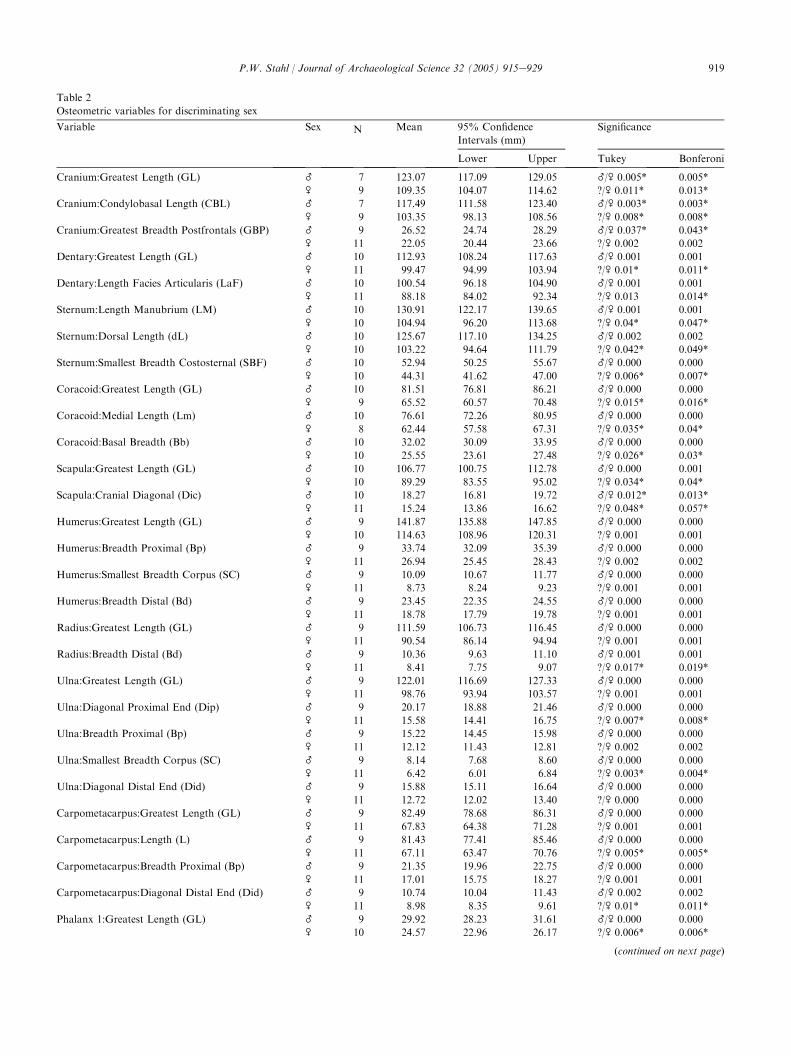
In order to be statistically conservative, only thosedependent variables that differed significantly ( p!0.05)from one another in both male/female and female/unknown comparisons are listed in Table 2. In no caseswere differences between male/unknown comparisonsstatistically significant. As some sex unknown specimensmight be female, considering only those variables withstatistically significant differences between both male/female and female/unknown categories avoids potentialoverlap. Table 2 lists 54 variables with significant valuesfor male/female and female/unknown comparisons,including sample size and significance scores (Tukeyand Bonferoni). Non-overlapping size criteria for malesand females are recorded, including their respectivemean values and lower and upper 95% confidenceboundaries in millimeters. p-values with an asterisk (*)exceed the critical p-value ( pR0.0023) suggested bya sequential Bonferoni correction which controls againstgroup-wide type 1 error [31].
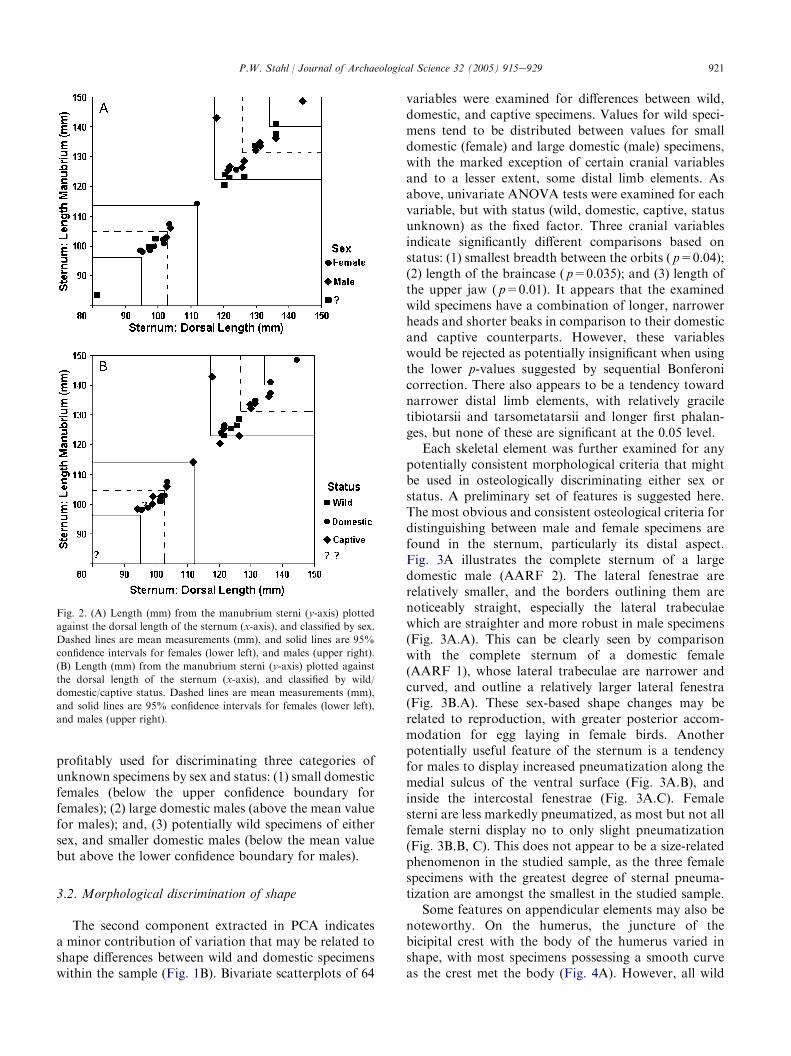
The non-overlapping ranges and their associatedmean values for each significant variable should be usefulfor discriminating males from females. Using two sternalmeasurements as an example, Fig. 2 plots manubriumlength (y-axis) against dorsal length (x-axis) for 31measured specimens, showing the lower and upper 95%confidence boundaries along with respective mean valuesfor female (lower left) and male specimens (upper right).Fig. 2A illustrates a clear separation of known male andfemale specimens, with the exception of one relativelysmall male, and one relatively large female. This clearseparation of male and female specimens is maintainedfor each of the 54 significant variables listed in Table 2.
Although the total sample size of measured wildspecimens is small, these data might also have someprovisional utility for discriminating wild from domesticspecimens osteometrically. Fig. 2B presents the samedata as Fig. 2A, but classified by wild/domestic/captivestatus. It is interesting to note that, with the exception ofone wild female specimen, all values below the upper
919P.W. Stahl / Journal of Archaeological Science 32 (2005) 915e929
Table 2
Osteometric variables for discriminating sex
Variable Sex N Mean 95% Confidence
Intervals (mm)
Significance
Lower Upper Tukey Bonferoni
Cranium:Greatest Length (GL) _ 7 123.07 117.09 129.05 _/\ 0.005* 0.005*
\ 9 109.35 104.07 114.62 ?/\ 0.011* 0.013*
Cranium:Condylobasal Length (CBL) _ 7 117.49 111.58 123.40 _/\ 0.003* 0.003*
\ 9 103.35 98.13 108.56 ?/\ 0.008* 0.008*
Cranium:Greatest Breadth Postfrontals (GBP) _ 9 26.52 24.74 28.29 _/\ 0.037* 0.043*
\ 11 22.05 20.44 23.66 ?/\ 0.002 0.002
Dentary:Greatest Length (GL) _ 10 112.93 108.24 117.63 _/\ 0.001 0.001
\ 11 99.47 94.99 103.94 ?/\ 0.01* 0.011*
Dentary:Length Facies Articularis (LaF) _ 10 100.54 96.18 104.90 _/\ 0.001 0.001
\ 11 88.18 84.02 92.34 ?/\ 0.013 0.014*
Sternum:Length Manubrium (LM) _ 10 130.91 122.17 139.65 _/\ 0.001 0.001
\ 10 104.94 96.20 113.68 ?/\ 0.04* 0.047*
Sternum:Dorsal Length (dL) _ 10 125.67 117.10 134.25 _/\ 0.002 0.002
\ 10 103.22 94.64 111.79 ?/\ 0.042* 0.049*
Sternum:Smallest Breadth Costosternal (SBF) _ 10 52.94 50.25 55.67 _/\ 0.000 0.000
\ 10 44.31 41.62 47.00 ?/\ 0.006* 0.007*
Coracoid:Greatest Length (GL) _ 10 81.51 76.81 86.21 _/\ 0.000 0.000
\ 9 65.52 60.57 70.48 ?/\ 0.015* 0.016*
Coracoid:Medial Length (Lm) _ 10 76.61 72.26 80.95 _/\ 0.000 0.000
\ 8 62.44 57.58 67.31 ?/\ 0.035* 0.04*
Coracoid:Basal Breadth (Bb) _ 10 32.02 30.09 33.95 _/\ 0.000 0.000
\ 10 25.55 23.61 27.48 ?/\ 0.026* 0.03*
Scapula:Greatest Length (GL) _ 10 106.77 100.75 112.78 _/\ 0.000 0.001
\ 10 89.29 83.55 95.02 ?/\ 0.034* 0.04*
Scapula:Cranial Diagonal (Dic) _ 10 18.27 16.81 19.72 _/\ 0.012* 0.013*
\ 11 15.24 13.86 16.62 ?/\ 0.048* 0.057*
Humerus:Greatest Length (GL) _ 9 141.87 135.88 147.85 _/\ 0.000 0.000
\ 10 114.63 108.96 120.31 ?/\ 0.001 0.001
Humerus:Breadth Proximal (Bp) _ 9 33.74 32.09 35.39 _/\ 0.000 0.000
\ 11 26.94 25.45 28.43 ?/\ 0.002 0.002
Humerus:Smallest Breadth Corpus (SC) _ 9 10.09 10.67 11.77 _/\ 0.000 0.000
\ 11 8.73 8.24 9.23 ?/\ 0.001 0.001
Humerus:Breadth Distal (Bd) _ 9 23.45 22.35 24.55 _/\ 0.000 0.000
\ 11 18.78 17.79 19.78 ?/\ 0.001 0.001
Radius:Greatest Length (GL) _ 9 111.59 106.73 116.45 _/\ 0.000 0.000
\ 11 90.54 86.14 94.94 ?/\ 0.001 0.001
Radius:Breadth Distal (Bd) _ 9 10.36 9.63 11.10 _/\ 0.001 0.001
\ 11 8.41 7.75 9.07 ?/\ 0.017* 0.019*
Ulna:Greatest Length (GL) _ 9 122.01 116.69 127.33 _/\ 0.000 0.000
\ 11 98.76 93.94 103.57 ?/\ 0.001 0.001
Ulna:Diagonal Proximal End (Dip) _ 9 20.17 18.88 21.46 _/\ 0.000 0.000
\ 11 15.58 14.41 16.75 ?/\ 0.007* 0.008*
Ulna:Breadth Proximal (Bp) _ 9 15.22 14.45 15.98 _/\ 0.000 0.000
\ 11 12.12 11.43 12.81 ?/\ 0.002 0.002
Ulna:Smallest Breadth Corpus (SC) _ 9 8.14 7.68 8.60 _/\ 0.000 0.000
\ 11 6.42 6.01 6.84 ?/\ 0.003* 0.004*
Ulna:Diagonal Distal End (Did) _ 9 15.88 15.11 16.64 _/\ 0.000 0.000
\ 11 12.72 12.02 13.40 ?/\ 0.000 0.000
Carpometacarpus:Greatest Length (GL) _ 9 82.49 78.68 86.31 _/\ 0.000 0.000
\ 11 67.83 64.38 71.28 ?/\ 0.001 0.001
Carpometacarpus:Length (L) _ 9 81.43 77.41 85.46 _/\ 0.000 0.000
\ 11 67.11 63.47 70.76 ?/\ 0.005* 0.005*
Carpometacarpus:Breadth Proximal (Bp) _ 9 21.35 19.96 22.75 _/\ 0.000 0.000
\ 11 17.01 15.75 18.27 ?/\ 0.001 0.001
Carpometacarpus:Diagonal Distal End (Did) _ 9 10.74 10.04 11.43 _/\ 0.002 0.002
\ 11 8.98 8.35 9.61 ?/\ 0.01* 0.011*
Phalanx 1:Greatest Length (GL) _ 9 29.92 28.23 31.61 _/\ 0.000 0.000
\ 10 24.57 22.96 26.17 ?/\ 0.006* 0.006*
(continued on next page)
920 P.W. Stahl / Journal of Archaeological Science 32 (2005) 915e929
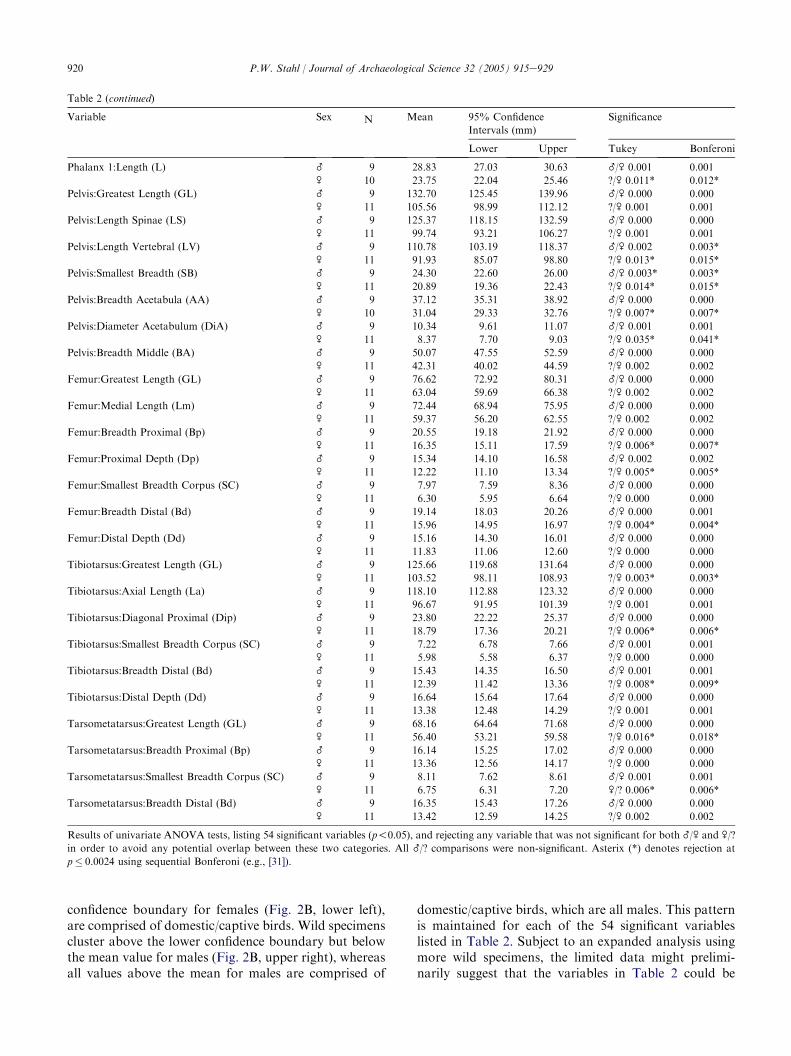
Table 2 (continued)
Variable Sex N Mean 95% Confidence
Intervals (mm)
Significance
Lower Upper Tukey Bonferoni
Phalanx 1:Length (L) _ 9 28.83 27.03 30.63 _/\ 0.001 0.001
\ 10 23.75 22.04 25.46 ?/\ 0.011* 0.012*
Pelvis:Greatest Length (GL) _ 9 132.70 125.45 139.96 _/\ 0.000 0.000
\ 11 105.56 98.99 112.12 ?/\ 0.001 0.001
Pelvis:Length Spinae (LS) _ 9 125.37 118.15 132.59 _/\ 0.000 0.000
\ 11 99.74 93.21 106.27 ?/\ 0.001 0.001
Pelvis:Length Vertebral (LV) _ 9 110.78 103.19 118.37 _/\ 0.002 0.003*
\ 11 91.93 85.07 98.80 ?/\ 0.013* 0.015*
Pelvis:Smallest Breadth (SB) _ 9 24.30 22.60 26.00 _/\ 0.003* 0.003*
\ 11 20.89 19.36 22.43 ?/\ 0.014* 0.015*
Pelvis:Breadth Acetabula (AA) _ 9 37.12 35.31 38.92 _/\ 0.000 0.000
\ 10 31.04 29.33 32.76 ?/\ 0.007* 0.007*
Pelvis:Diameter Acetabulum (DiA) _ 9 10.34 9.61 11.07 _/\ 0.001 0.001
\ 11 8.37 7.70 9.03 ?/\ 0.035* 0.041*
Pelvis:Breadth Middle (BA) _ 9 50.07 47.55 52.59 _/\ 0.000 0.000
\ 11 42.31 40.02 44.59 ?/\ 0.002 0.002
Femur:Greatest Length (GL) _ 9 76.62 72.92 80.31 _/\ 0.000 0.000
\ 11 63.04 59.69 66.38 ?/\ 0.002 0.002
Femur:Medial Length (Lm) _ 9 72.44 68.94 75.95 _/\ 0.000 0.000
\ 11 59.37 56.20 62.55 ?/\ 0.002 0.002
Femur:Breadth Proximal (Bp) _ 9 20.55 19.18 21.92 _/\ 0.000 0.000
\ 11 16.35 15.11 17.59 ?/\ 0.006* 0.007*
Femur:Proximal Depth (Dp) _ 9 15.34 14.10 16.58 _/\ 0.002 0.002
\ 11 12.22 11.10 13.34 ?/\ 0.005* 0.005*
Femur:Smallest Breadth Corpus (SC) _ 9 7.97 7.59 8.36 _/\ 0.000 0.000
\ 11 6.30 5.95 6.64 ?/\ 0.000 0.000
Femur:Breadth Distal (Bd) _ 9 19.14 18.03 20.26 _/\ 0.000 0.001
\ 11 15.96 14.95 16.97 ?/\ 0.004* 0.004*
Femur:Distal Depth (Dd) _ 9 15.16 14.30 16.01 _/\ 0.000 0.000
\ 11 11.83 11.06 12.60 ?/\ 0.000 0.000
Tibiotarsus:Greatest Length (GL) _ 9 125.66 119.68 131.64 _/\ 0.000 0.000
\ 11 103.52 98.11 108.93 ?/\ 0.003* 0.003*
Tibiotarsus:Axial Length (La) _ 9 118.10 112.88 123.32 _/\ 0.000 0.000
\ 11 96.67 91.95 101.39 ?/\ 0.001 0.001
Tibiotarsus:Diagonal Proximal (Dip) _ 9 23.80 22.22 25.37 _/\ 0.000 0.000
\ 11 18.79 17.36 20.21 ?/\ 0.006* 0.006*
Tibiotarsus:Smallest Breadth Corpus (SC) _ 9 7.22 6.78 7.66 _/\ 0.001 0.001
\ 11 5.98 5.58 6.37 ?/\ 0.000 0.000
Tibiotarsus:Breadth Distal (Bd) _ 9 15.43 14.35 16.50 _/\ 0.001 0.001
\ 11 12.39 11.42 13.36 ?/\ 0.008* 0.009*
Tibiotarsus:Distal Depth (Dd) _ 9 16.64 15.64 17.64 _/\ 0.000 0.000
\ 11 13.38 12.48 14.29 ?/\ 0.001 0.001
Tarsometatarsus:Greatest Length (GL) _ 9 68.16 64.64 71.68 _/\ 0.000 0.000
\ 11 56.40 53.21 59.58 ?/\ 0.016* 0.018*
Tarsometatarsus:Breadth Proximal (Bp) _ 9 16.14 15.25 17.02 _/\ 0.000 0.000
\ 11 13.36 12.56 14.17 ?/\ 0.000 0.000
Tarsometatarsus:Smallest Breadth Corpus (SC) _ 9 8.11 7.62 8.61 _/\ 0.001 0.001
\ 11 6.75 6.31 7.20 \/? 0.006* 0.006*
Tarsometatarsus:Breadth Distal (Bd) _ 9 16.35 15.43 17.26 _/\ 0.000 0.000
\ 11 13.42 12.59 14.25 ?/\ 0.002 0.002
Results of univariate ANOVA tests, listing 54 significant variables (p!0.05), and rejecting any variable that was not significant for both _/\ and \/?
in order to avoid any potential overlap between these two categories. All _/? comparisons were non-significant. Asterix (*) denotes rejection at
p� 0.0024 using sequential Bonferoni (e.g., [31]).
confidence boundary for females (Fig. 2B, lower left),are comprised of domestic/captive birds. Wild specimenscluster above the lower confidence boundary but belowthe mean value for males (Fig. 2B, upper right), whereasall values above the mean for males are comprised of
domestic/captive birds, which are all males. This patternis maintained for each of the 54 significant variableslisted in Table 2. Subject to an expanded analysis usingmore wild specimens, the limited data might prelimi-narily suggest that the variables in Table 2 could be
921P.W. Stahl / Journal of Archaeological Science 32 (2005) 915e929
profitably used for discriminating three categories ofunknown specimens by sex and status: (1) small domesticfemales (below the upper confidence boundary forfemales); (2) large domestic males (above the mean valuefor males); and, (3) potentially wild specimens of eithersex, and smaller domestic males (below the mean valuebut above the lower confidence boundary for males).
3.2. Morphological discrimination of shape
The second component extracted in PCA indicatesa minor contribution of variation that may be related toshape differences between wild and domestic specimenswithin the sample (Fig. 1B). Bivariate scatterplots of 64
Fig. 2. (A) Length (mm) from the manubrium sterni (y-axis) plotted
against the dorsal length of the sternum (x-axis), and classified by sex.
Dashed lines are mean measurements (mm), and solid lines are 95%
confidence intervals for females (lower left), and males (upper right).
(B) Length (mm) from the manubrium sterni (y-axis) plotted against
the dorsal length of the sternum (x-axis), and classified by wild/
domestic/captive status. Dashed lines are mean measurements (mm),
and solid lines are 95% confidence intervals for females (lower left),
and males (upper right).
variables were examined for differences between wild,domestic, and captive specimens. Values for wild speci-mens tend to be distributed between values for smalldomestic (female) and large domestic (male) specimens,with the marked exception of certain cranial variablesand to a lesser extent, some distal limb elements. Asabove, univariate ANOVA tests were examined for eachvariable, but with status (wild, domestic, captive, statusunknown) as the fixed factor. Three cranial variablesindicate significantly different comparisons based onstatus: (1) smallest breadth between the orbits ( p=0.04);(2) length of the braincase ( p=0.035); and (3) length ofthe upper jaw ( p=0.01). It appears that the examinedwild specimens have a combination of longer, narrowerheads and shorter beaks in comparison to their domesticand captive counterparts. However, these variableswould be rejected as potentially insignificant when usingthe lower p-values suggested by sequential Bonferonicorrection. There also appears to be a tendency towardnarrower distal limb elements, with relatively graciletibiotarsii and tarsometatarsii and longer first phalan-ges, but none of these are significant at the 0.05 level.
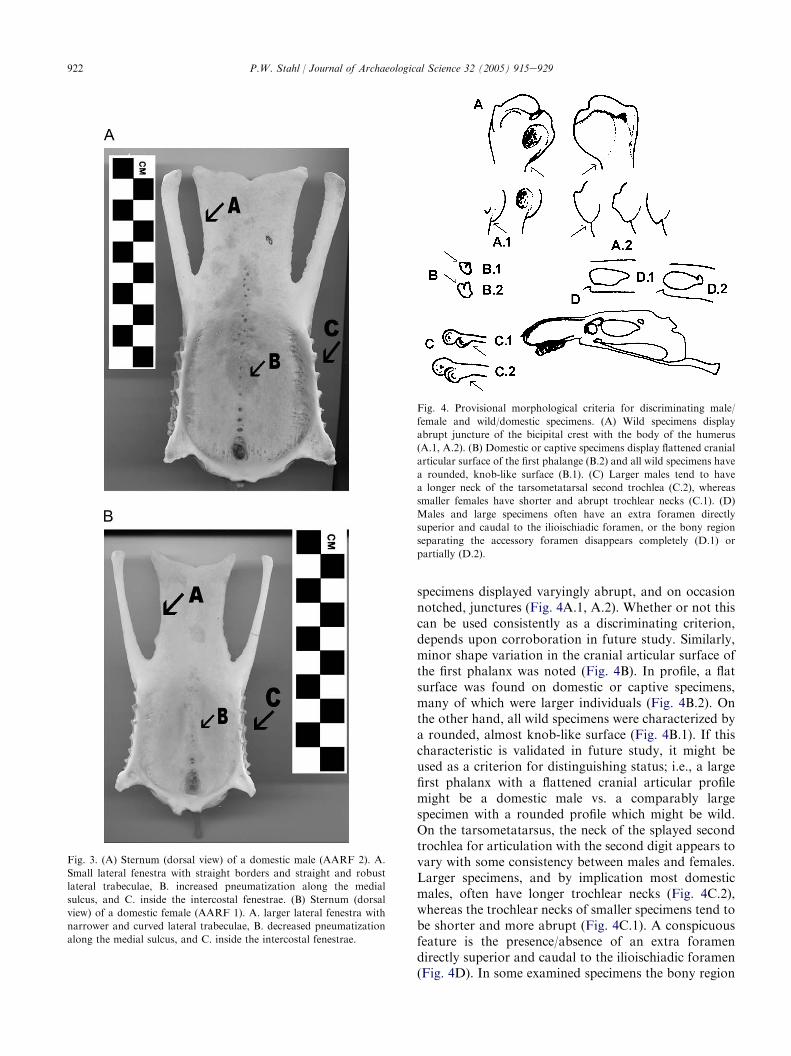
Each skeletal element was further examined for anypotentially consistent morphological criteria that mightbe used in osteologically discriminating either sex orstatus. A preliminary set of features is suggested here.The most obvious and consistent osteological criteria fordistinguishing between male and female specimens arefound in the sternum, particularly its distal aspect.Fig. 3A illustrates the complete sternum of a largedomestic male (AARF 2). The lateral fenestrae arerelatively smaller, and the borders outlining them arenoticeably straight, especially the lateral trabeculaewhich are straighter and more robust in male specimens(Fig. 3A.A). This can be clearly seen by comparisonwith the complete sternum of a domestic female(AARF 1), whose lateral trabeculae are narrower andcurved, and outline a relatively larger lateral fenestra(Fig. 3B.A). These sex-based shape changes may berelated to reproduction, with greater posterior accom-modation for egg laying in female birds. Anotherpotentially useful feature of the sternum is a tendencyfor males to display increased pneumatization along themedial sulcus of the ventral surface (Fig. 3A.B), andinside the intercostal fenestrae (Fig. 3A.C). Femalesterni are less markedly pneumatized, as most but not allfemale sterni display no to only slight pneumatization(Fig. 3B.B, C). This does not appear to be a size-relatedphenomenon in the studied sample, as the three femalespecimens with the greatest degree of sternal pneuma-tization are amongst the smallest in the studied sample.
Some features on appendicular elements may also benoteworthy. On the humerus, the juncture of thebicipital crest with the body of the humerus varied inshape, with most specimens possessing a smooth curveas the crest met the body (Fig. 4A). However, all wild
922 P.W. Stahl / Journal of Archaeological Science 32 (2005) 915e929
Fig. 3. (A) Sternum (dorsal view) of a domestic male (AARF 2). A.
Small lateral fenestra with straight borders and straight and robust
lateral trabeculae, B. increased pneumatization along the medial
sulcus, and C. inside the intercostal fenestrae. (B) Sternum (dorsal
view) of a domestic female (AARF 1). A. larger lateral fenestra with
narrower and curved lateral trabeculae, B. decreased pneumatization
along the medial sulcus, and C. inside the intercostal fenestrae.
specimens displayed varyingly abrupt, and on occasionnotched, junctures (Fig. 4A.1, A.2). Whether or not thiscan be used consistently as a discriminating criterion,depends upon corroboration in future study. Similarly,minor shape variation in the cranial articular surface ofthe first phalanx was noted (Fig. 4B). In profile, a flatsurface was found on domestic or captive specimens,many of which were larger individuals (Fig. 4B.2). Onthe other hand, all wild specimens were characterized bya rounded, almost knob-like surface (Fig. 4B.1). If thischaracteristic is validated in future study, it might beused as a criterion for distinguishing status; i.e., a largefirst phalanx with a flattened cranial articular profilemight be a domestic male vs. a comparably largespecimen with a rounded profile which might be wild.On the tarsometatarsus, the neck of the splayed secondtrochlea for articulation with the second digit appears tovary with some consistency between males and females.Larger specimens, and by implication most domesticmales, often have longer trochlear necks (Fig. 4C.2),whereas the trochlear necks of smaller specimens tend tobe shorter and more abrupt (Fig. 4C.1). A conspicuousfeature is the presence/absence of an extra foramendirectly superior and caudal to the ilioischiadic foramen(Fig. 4D). In some examined specimens the bony region
Fig. 4. Provisional morphological criteria for discriminating male/
female and wild/domestic specimens. (A) Wild specimens display
abrupt juncture of the bicipital crest with the body of the humerus
(A.1, A.2). (B) Domestic or captive specimens display flattened cranial
articular surface of the first phalange (B.2) and all wild specimens have
a rounded, knob-like surface (B.1). (C) Larger males tend to have
a longer neck of the tarsometatarsal second trochlea (C.2), whereas
smaller females have shorter and abrupt trochlear necks (C.1). (D)
Males and large specimens often have an extra foramen directly
superior and caudal to the ilioischiadic foramen, or the bony region
separating the accessory foramen disappears completely (D.1) or
partially (D.2).

923P.W. Stahl / Journal of Archaeological Science 32 (2005) 915e929
separating the accessory foramen disappears almostcompletely (Fig. 4D.1, D.2). Although there is onlya tendency for males to display this feature, it might bea size-related characteristic. The foramen tends toappear on the larger male synsacrum, and is oftenabsent on smaller female specimens, however withexceptions.
4. Discussion and archaeological application
Early European accounts document a wide geo-graphical distribution of domesticated muscovy ducksfrom Mexico in the north, southward throughoutCentral America and some of the Caribbean islands,into South America as far as Chile and Argentina (e.g.,[1:25,7:49, Table 5,13,41]. However, the prehistoricdistribution of domesticated muscovy is poorly known.Muscovy ducks are convincingly depicted on clay vesselsmade by the Moche and Chimu cultures of north coastalPeru (e.g., [2:84,14,23:58,26], and are possibly portrayedin Aztec codices and art from Mexico [41:4]. The use ofmuscovy ducks in Chimu featherwork has also beenestablished [26]. Nevertheless, osteological evidence isscarce, and when recovered and identified, requireseducated guesswork for inferring possible domesticatedor wild status.
Osteological evidence for muscovy ducks comes froma few, widely scattered localities (Fig. 5). This specieswas recorded at three localities on the central Pacificwatershed of Panama: (1) a purportedly upper Pleisto-cene fossil bed at El Hatillo in which extinct fauna wasalso recovered [39]; (2) Sitio Conte, a site whosecemetery was used mostly between AD 750 and AD950 [9] but whose refuse lenses, in which muscovy duckbones were reported in pioneering excavations byLothrop [24:16], include earlier and later deposits; and(3) Sitio Sierra where 16 specimens identified as Cairina,representing at least two individuals and some ducklingbones referable to this taxon on the basis of size, wererecovered in deposits that date to between AD 1 and AD500 [6:253,7:41]. Wetmore [38:329] briefly mentionsmuscovy duck in pre-columbian midden deposits atthe Hacienda Tocoron near Lake Valencia, Venezuela.Muscovy duck is also briefly mentioned in a collectionrecovered from three sites in Trinidad whose occupa-tions span the millennia after Christ [43:59].
Further south, published identifications of muscovyduck include specimens from the coastal lowlands ofEcuador. Numerous fragments of various skeletalelements were recovered at Ayalan in burial contextsassociated with the two final prehistoric occupationphases after the time of Christ [18]. From Salango, twospecimens were recovered in association with a pre-historic occupation phase dating to as early as AD 80[36]. In northern Peru, muscovy duck is probably
present in Mochica occupations of the Upper Piurariver after AD 500 [22:414], and appears in associationwith later Incaic structures and plazas in northwesternArgentina [33:222]. A large sample of 125 muscovyspecimens was recovered from two sites in the lowlandsof eastern Bolivia: Pailon 5 dating between AD 600 and1000; and, the majority from Pailon 6 dating to betweenAD 1100 and 1300 [19,30]. The domesticated status ofall these archaeologically recovered specimens remainsuncertain for lack of published diagnostic criteria. In thefollowing section, recently identified muscovy duckspecimens from two archaeological contexts in south-western Ecuador are discussed and examined in light ofthe aforementioned data for size and shape discrimina-tion of sex and wild/domesticated status.
4.1. Prehistoric muscovy duck insouthwestern Ecuador
Prehistoric muscovy duck has recently been identifiedat two archaeological sites in the lower Guayas Basin ofEcuador (Fig. 6). Roughly contemporaneous in age,both contexts are associated with intensive raised fieldagricultural production during the centuries preceding
Fig. 5. Areas with published accounts of archaeologically recovered
and identified muscovy duck.
924 P.W. Stahl / Journal of Archaeological Science 32 (2005) 915e929
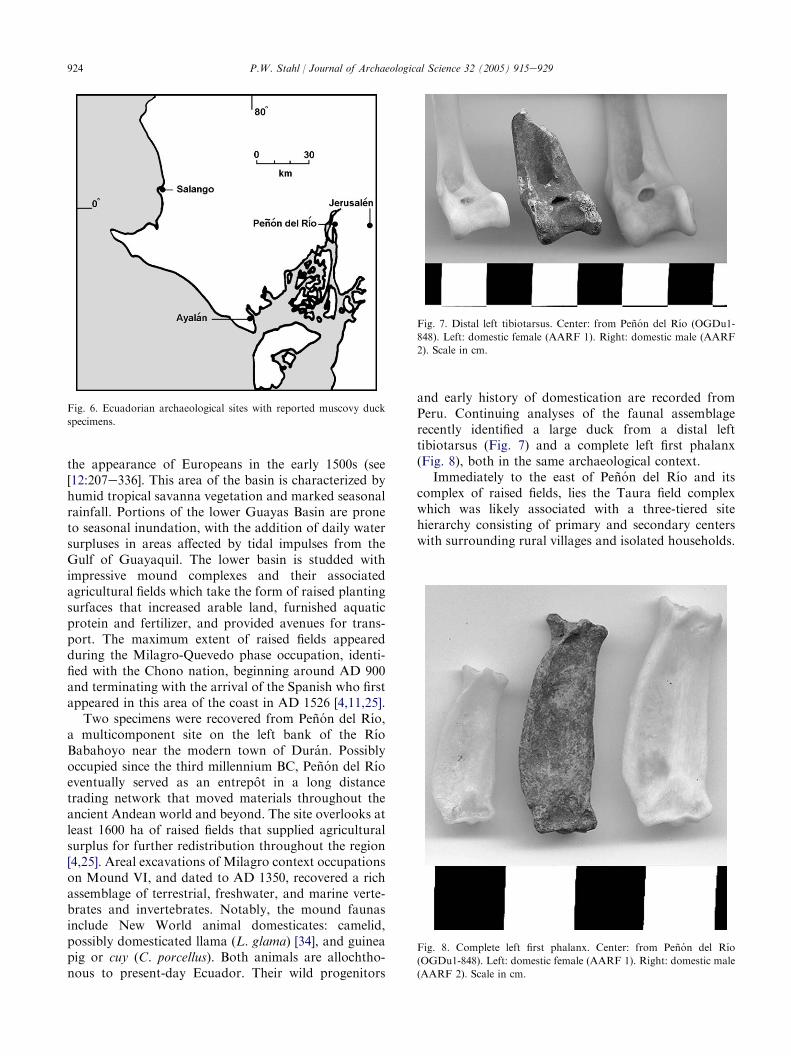
the appearance of Europeans in the early 1500s (see[12:207e336]. This area of the basin is characterized byhumid tropical savanna vegetation and marked seasonalrainfall. Portions of the lower Guayas Basin are proneto seasonal inundation, with the addition of daily watersurpluses in areas affected by tidal impulses from theGulf of Guayaquil. The lower basin is studded withimpressive mound complexes and their associatedagricultural fields which take the form of raised plantingsurfaces that increased arable land, furnished aquaticprotein and fertilizer, and provided avenues for trans-port. The maximum extent of raised fields appearedduring the Milagro-Quevedo phase occupation, identi-fied with the Chono nation, beginning around AD 900and terminating with the arrival of the Spanish who firstappeared in this area of the coast in AD 1526 [4,11,25].
Two specimens were recovered from Penon del Rıo,a multicomponent site on the left bank of the RıoBabahoyo near the modern town of Duran. Possiblyoccupied since the third millennium BC, Penon del Rıoeventually served as an entrepot in a long distancetrading network that moved materials throughout theancient Andean world and beyond. The site overlooks atleast 1600 ha of raised fields that supplied agriculturalsurplus for further redistribution throughout the region[4,25]. Areal excavations of Milagro context occupationson Mound VI, and dated to AD 1350, recovered a richassemblage of terrestrial, freshwater, and marine verte-brates and invertebrates. Notably, the mound faunasinclude New World animal domesticates: camelid,possibly domesticated llama (L. glama) [34], and guineapig or cuy (C. porcellus). Both animals are allochtho-nous to present-day Ecuador. Their wild progenitors
Fig. 6. Ecuadorian archaeological sites with reported muscovy duck
specimens.
and early history of domestication are recorded fromPeru. Continuing analyses of the faunal assemblagerecently identified a large duck from a distal lefttibiotarsus (Fig. 7) and a complete left first phalanx(Fig. 8), both in the same archaeological context.
Immediately to the east of Penon del Rıo and itscomplex of raised fields, lies the Taura field complexwhich was likely associated with a three-tiered sitehierarchy consisting of primary and secondary centerswith surrounding rural villages and isolated households.
Fig. 7. Distal left tibiotarsus. Center: from Penon del Rıo (OGDu1-
848). Left: domestic female (AARF 1). Right: domestic male (AARF
2). Scale in cm.
Fig. 8. Complete left first phalanx. Center: from Penon del Rıo
(OGDu1-848). Left: domestic female (AARF 1). Right: domestic male
(AARF 2). Scale in cm.
925P.W. Stahl / Journal of Archaeological Science 32 (2005) 915e929
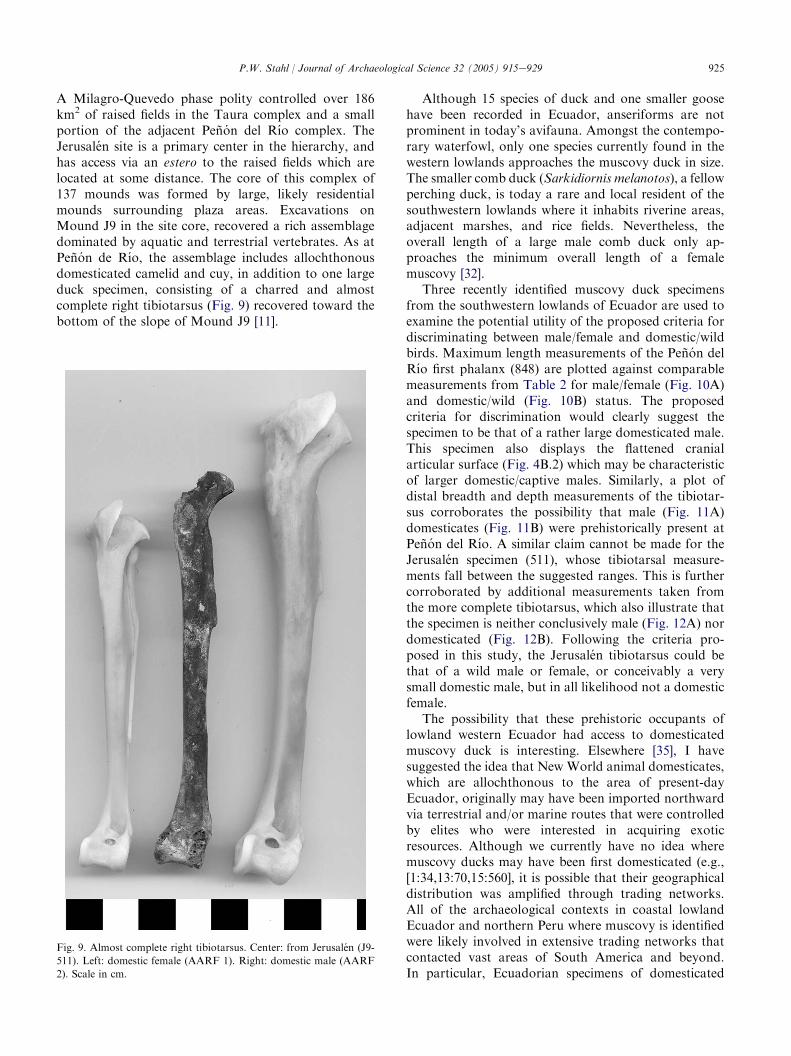
A Milagro-Quevedo phase polity controlled over 186km2 of raised fields in the Taura complex and a smallportion of the adjacent Penon del Rıo complex. TheJerusalen site is a primary center in the hierarchy, andhas access via an estero to the raised fields which arelocated at some distance. The core of this complex of137 mounds was formed by large, likely residentialmounds surrounding plaza areas. Excavations onMound J9 in the site core, recovered a rich assemblagedominated by aquatic and terrestrial vertebrates. As atPenon de Rıo, the assemblage includes allochthonousdomesticated camelid and cuy, in addition to one largeduck specimen, consisting of a charred and almostcomplete right tibiotarsus (Fig. 9) recovered toward thebottom of the slope of Mound J9 [11].
Fig. 9. Almost complete right tibiotarsus. Center: from Jerusalen (J9-
511). Left: domestic female (AARF 1). Right: domestic male (AARF
2). Scale in cm.
Although 15 species of duck and one smaller goosehave been recorded in Ecuador, anseriforms are notprominent in today’s avifauna. Amongst the contempo-rary waterfowl, only one species currently found in thewestern lowlands approaches the muscovy duck in size.The smaller comb duck (Sarkidiornis melanotos), a fellowperching duck, is today a rare and local resident of thesouthwestern lowlands where it inhabits riverine areas,adjacent marshes, and rice fields. Nevertheless, theoverall length of a large male comb duck only ap-proaches the minimum overall length of a femalemuscovy [32].
Three recently identified muscovy duck specimensfrom the southwestern lowlands of Ecuador are used toexamine the potential utility of the proposed criteria fordiscriminating between male/female and domestic/wildbirds. Maximum length measurements of the Penon delRıo first phalanx (848) are plotted against comparablemeasurements from Table 2 for male/female (Fig. 10A)and domestic/wild (Fig. 10B) status. The proposedcriteria for discrimination would clearly suggest thespecimen to be that of a rather large domesticated male.This specimen also displays the flattened cranialarticular surface (Fig. 4B.2) which may be characteristicof larger domestic/captive males. Similarly, a plot ofdistal breadth and depth measurements of the tibiotar-sus corroborates the possibility that male (Fig. 11A)domesticates (Fig. 11B) were prehistorically present atPenon del Rıo. A similar claim cannot be made for theJerusalen specimen (511), whose tibiotarsal measure-ments fall between the suggested ranges. This is furthercorroborated by additional measurements taken fromthe more complete tibiotarsus, which also illustrate thatthe specimen is neither conclusively male (Fig. 12A) nordomesticated (Fig. 12B). Following the criteria pro-posed in this study, the Jerusalen tibiotarsus could bethat of a wild male or female, or conceivably a verysmall domestic male, but in all likelihood not a domesticfemale.
The possibility that these prehistoric occupants oflowland western Ecuador had access to domesticatedmuscovy duck is interesting. Elsewhere [35], I havesuggested the idea that New World animal domesticates,which are allochthonous to the area of present-dayEcuador, originally may have been imported northwardvia terrestrial and/or marine routes that were controlledby elites who were interested in acquiring exoticresources. Although we currently have no idea wheremuscovy ducks may have been first domesticated (e.g.,[1:34,13:70,15:560], it is possible that their geographicaldistribution was amplified through trading networks.All of the archaeological contexts in coastal lowlandEcuador and northern Peru where muscovy is identifiedwere likely involved in extensive trading networks thatcontacted vast areas of South America and beyond.In particular, Ecuadorian specimens of domesticated
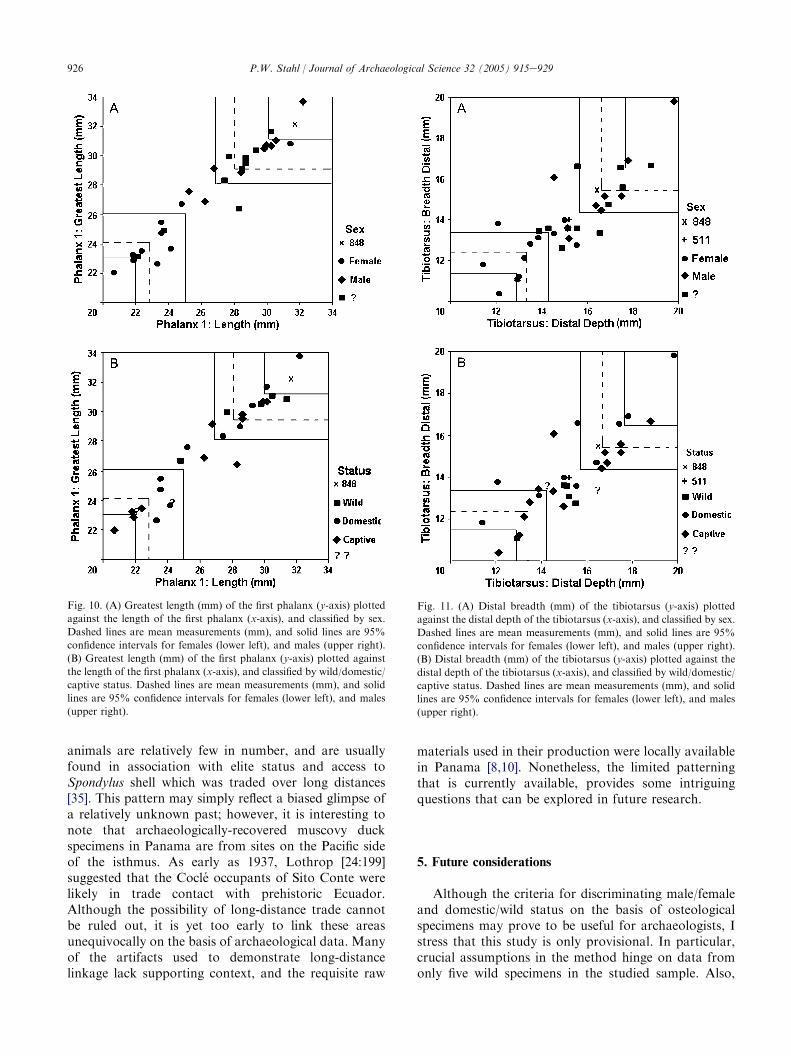
926 P.W. Stahl / Journal of Archaeological Science 32 (2005) 915e929
animals are relatively few in number, and are usuallyfound in association with elite status and access toSpondylus shell which was traded over long distances[35]. This pattern may simply reflect a biased glimpse ofa relatively unknown past; however, it is interesting tonote that archaeologically-recovered muscovy duckspecimens in Panama are from sites on the Pacific sideof the isthmus. As early as 1937, Lothrop [24:199]suggested that the Cocle occupants of Sito Conte werelikely in trade contact with prehistoric Ecuador.Although the possibility of long-distance trade cannotbe ruled out, it is yet too early to link these areasunequivocally on the basis of archaeological data. Manyof the artifacts used to demonstrate long-distancelinkage lack supporting context, and the requisite raw
Fig. 10. (A) Greatest length (mm) of the first phalanx (y-axis) plotted
against the length of the first phalanx (x-axis), and classified by sex.
Dashed lines are mean measurements (mm), and solid lines are 95%
confidence intervals for females (lower left), and males (upper right).
(B) Greatest length (mm) of the first phalanx (y-axis) plotted against
the length of the first phalanx (x-axis), and classified by wild/domestic/
captive status. Dashed lines are mean measurements (mm), and solid
lines are 95% confidence intervals for females (lower left), and males
(upper right).
materials used in their production were locally availablein Panama [8,10]. Nonetheless, the limited patterningthat is currently available, provides some intriguingquestions that can be explored in future research.
5. Future considerations
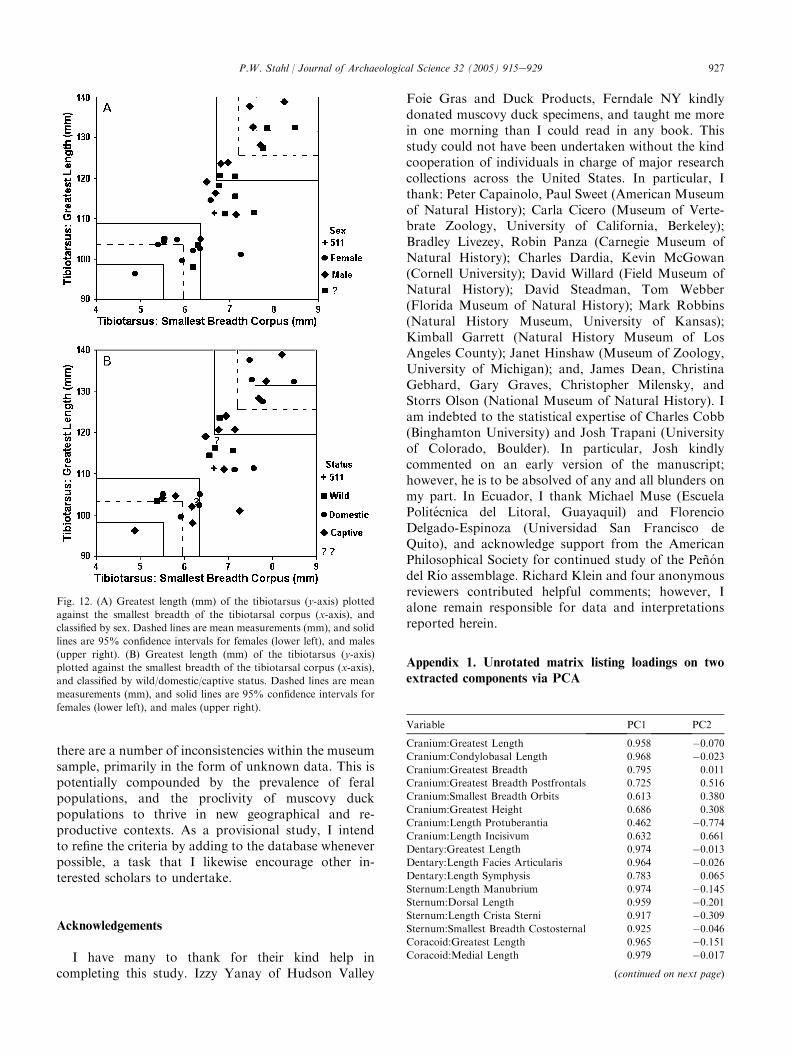
Although the criteria for discriminating male/femaleand domestic/wild status on the basis of osteologicalspecimens may prove to be useful for archaeologists, Istress that this study is only provisional. In particular,crucial assumptions in the method hinge on data fromonly five wild specimens in the studied sample. Also,
Fig. 11. (A) Distal breadth (mm) of the tibiotarsus (y-axis) plotted
against the distal depth of the tibiotarsus (x-axis), and classified by sex.
Dashed lines are mean measurements (mm), and solid lines are 95%
confidence intervals for females (lower left), and males (upper right).
(B) Distal breadth (mm) of the tibiotarsus (y-axis) plotted against the
distal depth of the tibiotarsus (x-axis), and classified by wild/domestic/
captive status. Dashed lines are mean measurements (mm), and solid
lines are 95% confidence intervals for females (lower left), and males
(upper right).
927P.W. Stahl / Journal of Archaeological Science 32 (2005) 915e929
there are a number of inconsistencies within the museumsample, primarily in the form of unknown data. This ispotentially compounded by the prevalence of feralpopulations, and the proclivity of muscovy duckpopulations to thrive in new geographical and re-productive contexts. As a provisional study, I intendto refine the criteria by adding to the database wheneverpossible, a task that I likewise encourage other in-terested scholars to undertake.
Acknowledgements
I have many to thank for their kind help incompleting this study. Izzy Yanay of Hudson Valley
Fig. 12. (A) Greatest length (mm) of the tibiotarsus (y-axis) plotted
against the smallest breadth of the tibiotarsal corpus (x-axis), and
classified by sex. Dashed lines are mean measurements (mm), and solid
lines are 95% confidence intervals for females (lower left), and males
(upper right). (B) Greatest length (mm) of the tibiotarsus (y-axis)
plotted against the smallest breadth of the tibiotarsal corpus (x-axis),
and classified by wild/domestic/captive status. Dashed lines are mean
measurements (mm), and solid lines are 95% confidence intervals for
females (lower left), and males (upper right).
Foie Gras and Duck Products, Ferndale NY kindlydonated muscovy duck specimens, and taught me morein one morning than I could read in any book. Thisstudy could not have been undertaken without the kindcooperation of individuals in charge of major researchcollections across the United States. In particular, Ithank: Peter Capainolo, Paul Sweet (American Museumof Natural History); Carla Cicero (Museum of Verte-brate Zoology, University of California, Berkeley);Bradley Livezey, Robin Panza (Carnegie Museum ofNatural History); Charles Dardia, Kevin McGowan(Cornell University); David Willard (Field Museum ofNatural History); David Steadman, Tom Webber(Florida Museum of Natural History); Mark Robbins(Natural History Museum, University of Kansas);Kimball Garrett (Natural History Museum of LosAngeles County); Janet Hinshaw (Museum of Zoology,University of Michigan); and, James Dean, ChristinaGebhard, Gary Graves, Christopher Milensky, andStorrs Olson (National Museum of Natural History). Iam indebted to the statistical expertise of Charles Cobb(Binghamton University) and Josh Trapani (Universityof Colorado, Boulder). In particular, Josh kindlycommented on an early version of the manuscript;however, he is to be absolved of any and all blunders onmy part. In Ecuador, I thank Michael Muse (EscuelaPolitecnica del Litoral, Guayaquil) and FlorencioDelgado-Espinoza (Universidad San Francisco deQuito), and acknowledge support from the AmericanPhilosophical Society for continued study of the Penondel Rıo assemblage. Richard Klein and four anonymousreviewers contributed helpful comments; however, Ialone remain responsible for data and interpretationsreported herein.
Appendix 1. Unrotated matrix listing loadings on two
extracted components via PCA
Variable PC1 PC2
Cranium:Greatest Length 0.958 �0.070
Cranium:Condylobasal Length 0.968 �0.023
Cranium:Greatest Breadth 0.795 0.011
Cranium:Greatest Breadth Postfrontals 0.725 0.516
Cranium:Smallest Breadth Orbits 0.613 0.380
Cranium:Greatest Height 0.686 0.308
Cranium:Length Protuberantia 0.462 �0.774
Cranium:Length Incisivum 0.632 0.661
Dentary:Greatest Length 0.974 �0.013
Dentary:Length Facies Articularis 0.964 �0.026
Dentary:Length Symphysis 0.783 0.065
Sternum:Length Manubrium 0.974 �0.145
Sternum:Dorsal Length 0.959 �0.201
Sternum:Length Crista Sterni 0.917 �0.309
Sternum:Smallest Breadth Costosternal 0.925 �0.046
Coracoid:Greatest Length 0.965 �0.151
Coracoid:Medial Length 0.979 �0.017
(continued on next page)

928 P.W. Stahl / Journal of Archaeological Science 32 (2005) 915e929
References
[1] E.G. Angulo, Interpretacion biologica acerca de la domesticacion
del Pato Criollo (Cairina moschata), Bulletin de L’Institut
Francais d’Etudes Andines 27 (1998) 17e40.
[2] W.C. Bennett, Archaeology of the North Coast of Peru. An
Account of Exploration and Excavation in Viru and Lambayeque
Valleys, in: American Museum of Natural History Anthropolog-
ical Papers, vol. 37, No. 1, New York, 1939.
[3] E.R. Blake, Manual of neotropical birds, vol. 1, University of
Chicago Press, Chicago, 1977.
[4] J.E. Buys, M. Muse, Arqueologıa de Asentamientos Asociados
a los Campos Elevados de Penon del Rıo, Guayas, Ecuador,
Appendix 1 (continued)
Variable PC1 PC2
Coracoid:Basal Breadth 0.962 �0.079
Coracoid:Breadth Facies Articularis 0.863 0.002
Scapula:Greatest Length 0.985 �0.015
Scapula:Cranial Diagonal 0.901 0.263
Humerus:Greatest Length 0.970 �0.144
Humerus:Breadth Proximal 0.965 �0.022
Humerus:Smallest Breadth Corpus 0.908 �0.097
Humerus:Breadth Distal 0.981 �0.101
Radius:Greatest Length 0.955 �0.184
Radius:Smallest Breadth Corpus 0.677 �0.303
Radius:Breadth Distal 0.871 0.117
Ulna:Greatest Length 0.967 �0.201
Ulna:Diagonal Proximal End 0.984 0.043
Ulna:Breadth Proximal 0.968 �0.130
Ulna:Smallest Breadth Corpus 0.849 �0.322
Ulna:Diagonal Distal End 0.959 �0.163
Carpometacarpus:Greatest Length 0.946 �0.268
Carpometacarpus:Length 0.944 �0.260
Carpometacarpus:Breadth Proximal 0.915 0.066
Carpometacarpus:Diagonal Distal End 0.835 0.153
Phalanx 1:Greatest Length 0.826 �0.471
Phalanx 1:Length 0.843 �0.525
Pelvis:Greatest Length 0.971 �0.031
Pelvis:Length Spina 0.964 �0.014
Pelvis:Length Vertebral 0.832 0.027
Pelvis:Cranial Breadth 0.777 0.070
Pelvis:Smallest Breadth 0.729 0.093
Pelvis:Breadth Acetabula 0.926 �0.060
Pelvis:Diameter Acetabulum 0.828 0.168
Pelvis:Breadth Middle 0.888 �0.125
Femur:Greatest Length 0.975 0.055
Femur:Medial Length 0.977 0.010
Femur:Breadth Proximal 0.885 0.275
Femur:Proximal Depth 0.839 �0.029
Femur:Smallest Breadth Corpus 0.925 0.112
Femur:Breadth Distal 0.918 0.274
Femur:Distal Depth 0.899 0.103
Tibiotarsus:Greatest Length 0.970 �0.005
Tibiotarsus:Axial Length 0.970 �0.101
Tibiotarsus:Diagonal Proximal 0.922 0.292
Tibiotarsus:Smallest Breadth Corpus 0.882 0.096
Tibiotarsus:Breadth Distal 0.889 0.354
Tibiotarsus:Distal Depth 0.889 0.112
Tarsometatarsus:Greatest Length 0.953 0.001
Tarsometatarsus:Breadth Proximal 0.916 0.300
Tarsometatarsus:Smallest Breadth Corpus 0.881 0.389
Tarsometatarsus:Breadth Distal 0.904 0.280
in: W.M. Denevan, K. Mathewson, G. Knapp (Eds.), Prehispanic
Agricultural Fields in the Andean Region, British Archaeological
Reports International Series 359 (ii), Oxford, 1987, pp. 225e248.
[5] G.A. Clayton, Muscovy duck, in: I.L. Mason (Ed.), Evolution of
Domesticated Animals, Longman, London, 1984, pp. 340e344.
[6] R.A. Cooke, Birds and men in prehistoric Central Panama, in:
F.W. Lange (Ed.), Recent Developments in Isthmian Archaeol-
ogy: Advances in the Prehistory of Lower Central America,
British Archaeological Reports International Series 212, Oxford,
1984, pp. 243e281.
[7] R.A. Cooke, A.J. Ranere, Precolumbian influences on the
zoogeography of Panama: an update based on archaeofaunal
and documentary data, Tulane Studies in Zoology and Botany,
Supplementary Publication 1 (1992) 21e58.
[8] R.A. Cooke, L.A. Sanchez, El papel del mar de las costas en el
Panama pre-Hispanico y del perıodo del contacto: redes locales y
relaciones externas, Revistas de Historia 43 (2001) 15e60.
[9] R.A. Cooke, L.A. Sanchez Herrera, K. Udagawa, Contextualized
goldwork from ‘‘Gran Cocle’’, Panama: an update based on
recent excavations and new radiocarbon dates for associated
pottery styles, in: C. McEwan (Ed.), Precolumbian Gold,
Technology, Style and Iconography, British Museum Press,
London, 2000, pp. 154e176.[10] R.A. Cooke, I. Isaza, J. Griggs, B. Desjardins, L.A. Sanchez, Who
crafted, exchanged, and displayed gold in pre-Columbian Pana-
ma? in: J. Quilter, J.W. Hoopes (Eds.), Gold and Power in Ancient
Costa Rica, Panama, and Colombia, Dumbarton Oaks Trustees
for Harvard University, Washington, DC, 2003, pp. 91e158.
[11] F. Delgado-Espinoza, Intensive agricultural and political eco-
nomy of the Yaguachi Chiefdom of Guayas Basin, Coastal
Ecuador, PhD Dissertation, University of Pittsburgh, Pittsburgh,
2002.
[12] W.M. Denevan, K. Mathewson, G. Knapp (Eds.), Prehispanic
Agricultural Fields in the Andean Region, British Archaeological
Reports International Series 359 (ii), Oxford, 1987.
[13] R.A. Donkin, The muscovy duck, Cairina moschata domestica.
Origins, dispersal, and associated aspects of the geography of
domestication, A.A. Balkema, Rotterdam, 1989.
[14] C.B. Donnan, Moche art of Peru, Pre-Columbian symbolic
communication, University of California Museum of Cultural
History, Los Angeles, 1978.
[15] D.W. Gade, Muscovy ducks, in: K.F. Kiple, K.C. Ornela (Eds.),
The Cambridge world history of food, vol. 1, Cambridge
University Press, Cambridge, 2000, pp. 559e561.
[16] U. Gille, F.-V. Salomon, Growth of duck bills, The Condor 101
(1999) 710e713.
[17] A. Gentry, J. Clutton-Brock, C.P. Groves, The naming of wild
species and their domestic derivatives, Journal of Archaeological
Science 31 (2004) 645e651.[18] B. Hesse, Archaeological evidence for muscovy duck in Ecuador,
Current Anthropology 21 (1980) 139e140.
[19] R. Hutterer, Archaeozoological remains (Vertebrata, Gastro-
poda) from prehispanic sites at Pailon, Bolivia, Beitrage zur
Allgemeinen und Vergleichenden Archaologie 17 (1997) 325e341.
[20] J.A. Jobling, A dictionary of scientific bird names, Oxford
University Press, Oxford, 1991.
[21] P.A. Johnsgard, Waterfowl of North America, Indiana University
Press, Bloomington, 1975.
[22] P. Kaulicke, El perıodo intermedio temprano en el Alto Piura:
avances del proyecto arqueologico ‘‘Alto Piura’’ (1987e1990),Bulletin de L’Institut Francais d’Etudes Andines 20 (1991) 381e
422.
[23] D. Lavallee, Les representations animales dans la ceramique
Mochica, Universite de Paris, Memoires de L’Institut D’Ethno-
logie IV, Paris, 1970.
[24] S.K. Lothrop, Cocle, AnArchaeological Study of Central Panama,
Part 1, Historical Background, Excavations at the Sitio Conte,

929P.W. Stahl / Journal of Archaeological Science 32 (2005) 915e929
Artifacts and Ornaments, Memoirs of the Peabody Museum
of Archaeology and Ethnology, vol. 7, Harvard University,
Cambridge, MA, 1937.
[25] M. Muse, Products and politics of a milagro entrepot: Penon del
Rıo Guayas Basin, Ecuador, Research in Economic Anthropol-
ogy 13 (1991) 269e323.
[26] J.P. O’Neill, Introduction: feather identification, in: A.P. Rowe
(Ed.), Costumes and Featherwork of the Lords of Chimor, The
Textile Museum, Washington, DC, 1984, pp. 145e150.
[27] R.S. Palmer (Ed.), Handbook of North American birds, vol. 3,
Yale University Press, New Haven, 1976.
[28] J.C. Phillips, A natural history of the ducks, vol. 1, Dover
Publications, New York, 1986.
[29] H. Pingel, K. Trettner, Mast- und Schlachtleistung von Moschu-
senten (Cairina moschata) und deren Kreuzungen mit Pekingenten
(Anas plathyrinchos), Archiv fur Tierzucht 30 (1987) 523e527.[30] H. Prumers, Excavaciones arquelologicas en Pailon (Depto. De
Sta. Cruz, Bolivia), Beitrage zur Allgemeinen und Vergleichenden
Archaologie 22 (2002) 95e213.[31] W.R. Rice, Analyzing tables of statistical tests, Evolution 43
(1989) 223e225.
[32] R.S. Ridgely, P.J. Greenfield, The birds of Ecuador, vol. 2, Field
guide, Cornell University Press, Ithaca.
[33] C. Rodriguez Loredo, Estudio zooarqueologico del Sitio Inca
Potrero-Chaquiago Barrios La Solana y Retambay Andalgala
Pcia. de Catamarca (Argentina), Relaciones de la Sociedad
Argentina de Antropologıa 22e23 (1997e1998) 203e245.
[34] P.W. Stahl, Prehistoric Camelids in the lowlands of
Western Ecuador, Journal of Archaeological Science 15 (1988)
355e365.
[35] P.W. Stahl, Pre-Columbian Andean animal domesticates at the
edge of empire, World Archaeology 34 (2003) 470e483.[36] P.W. Stahl, P. Norton, Precolumbian animal domesticates from
Salango, Ecuador, American Antiquity 52 (1987) 382e391.
[37] A. von den Driesch, A guide to the measurement of animal bones
from archaeological sites, Peabody Museum of Archaeology and
Ethnology Bulletin 1, Cambridge, MA, 1976.
[38] A. Wetmore, Pre-Columbian bird remains from Venezuela, Auk
52 (1935) 328e329.[39] A. Wetmore, The muscovy duck in the Pleistocene of Panama,
Wilson Bulletin 68 (1956) 327.
[40] A. Wetmore, The Birds of the Republic of Panama, Part 1, e
Tinamidae (Tinamous) to Rhyncopidae (Skimmers), Smithsonian
Miscellaneous Collections, vol. 150, Washington, DC.
[41] G.R. Whitley, The muscovy duck in Mexico, Anthropological
Journal of Canada 11 (2) (1973) 2e8.[42] G. Williamson, W.J.A. Payne, An introduction to animal
husbandry in the tropics, Longman, London, 1999.
[43] E.S. Wing, Factors influencing exploitation of marine resources,
in: E.P. Benson (Ed.), The Sea in the Pre-Columbian World,
Trustees for Harvard University, Washington, DC, 1977,
pp. 47e64.
[44] E.R. Woodyard, E.G. Bolen, Ecological studies of muscovy ducks
in Mexico, Southwestern Naturalist 29 (1984) 453e461.


















