
An Automated Device to Increase Screening Throughput of ...
90
e University of Maine DigitalCommons@UMaine Electronic eses and Dissertations Fogler Library Summer 8-1-2017 An Automated Device to Increase Screening roughput of Zebrafish Larvae Fuoad Saliou-Sulley University of Maine, [email protected] Follow this and additional works at: hp://digitalcommons.library.umaine.edu/etd Part of the Biological Engineering Commons , Biomedical Devices and Instrumentation Commons , and the Other Biomedical Engineering and Bioengineering Commons is Open-Access esis is brought to you for free and open access by DigitalCommons@UMaine. It has been accepted for inclusion in Electronic eses and Dissertations by an authorized administrator of DigitalCommons@UMaine. For more information, please contact [email protected]. Recommended Citation Saliou-Sulley, Fuoad, "An Automated Device to Increase Screening roughput of Zebrafish Larvae" (2017). Electronic eses and Dissertations. 2720. hp://digitalcommons.library.umaine.edu/etd/2720
Transcript of An Automated Device to Increase Screening Throughput of ...

The University of MaineDigitalCommons@UMaine
Electronic Theses and Dissertations Fogler Library
Summer 8-1-2017
An Automated Device to Increase ScreeningThroughput of Zebrafish LarvaeFuoad Saliou-SulleyUniversity of Maine, [email protected]
Follow this and additional works at: http://digitalcommons.library.umaine.edu/etd
Part of the Biological Engineering Commons, Biomedical Devices and InstrumentationCommons, and the Other Biomedical Engineering and Bioengineering Commons
This Open-Access Thesis is brought to you for free and open access by DigitalCommons@UMaine. It has been accepted for inclusion in ElectronicTheses and Dissertations by an authorized administrator of DigitalCommons@UMaine. For more information, please [email protected].
Recommended CitationSaliou-Sulley, Fuoad, "An Automated Device to Increase Screening Throughput of Zebrafish Larvae" (2017). Electronic Theses andDissertations. 2720.http://digitalcommons.library.umaine.edu/etd/2720

ANAUTOMATEDDEVICETOINCREASESCREENINGTHROUGHPUTOF
ZEBRAFISHLARVAE
By
FuoadSaliou-Sulley
B.S.UniversityofMaine,2014
ATHESIS
SubmittedinPartialFulfillmentofthe
RequirementsfortheDegreeof
MasterofScience
(inBiologicalEngineering)
TheGraduateSchool
TheUniversityofMaine
August2017
AdvisoryCommittee:
PaulJ.Millard,AssociateProfessorofChemicalandBiologicalEngineering,
Advisor
DouglasW.Bousfield,ProfessorofChemicalandBiologicalEngineering
SaraWalton,Lecturer,ChemicalandBiologicalEngineering

ii
©2017FuoadSaliou-Sulley
AllRightsReserved

ANAUTOMATEDDEVICETOINCREASESCREENINGTHROUGHPUTOF
ZEBRAFISHLARVAE
ByFuoadSaliou-Sulley
ThesisAdvisor:Dr.PaulMillard
AnAbstractoftheThesisPresented
inPartialFulfillmentofthe
Requirementsforthe
DegreeofMasterofScience
(inBiologicalEngineering)
August2017
Theuseofthezebrafishasananimalmodelalternativetomammalian
specieshasspawnedresearchadvancementsinseveralmedicalfields1.Sincethe
zebrafishsharesahighdegreeofsequenceandfunctionalhomologywithmammals,
studiesusingthisorganismcanprovidein-depthinsightintohostresponseto
diseaseandprovideaplatformfortestingarangeoftreatmentoptions.Theoptical
transparencyofzebrafishatearlystagesofdevelopmentpermitseasyassessmentof
theeffectsoftreatments,occurrenceoftumorsorotherabnormalgrowth,disease
progression,andimmuneresponse.Thesecharacteristicsmakeitidealforstudying
humandiseasessuchastheInfluenzaAVirus(IAV).Theconventionalmethodof
IAVtransmissionisbyaerosol,andsincethezebrafishdoesnotaccommodatethis
modeofentry,thevirusisinjectedintothespecimenunderstudy.

Thelarvaetypicallyrequiremanipulationduringpreparatoryprocedures
priortoassessment,aprocessthatcanbetimeconsumingandstressfulforthe
organism.
Inthisthesis,Idescribethedevelopmentofadevicedesignedtoeliminate
problemsassociatedwithmanipulatingzebrafishlarvaebyautomatically
conductingspecimenfromareservoirdirectlyintoanentrapmentdock,whereit
willbeimmobilizedforinjectionandrapidlyremovedpost-injection.Thiswillhelp
toreducethehandlingtimeoflargesamplesets,therebyincreasingthescreening
throughput.Zebrafishhavefastgrowthrates4hencepreparatoryproceduresfor
analysislikeinjectionshouldbeasquickandefficientaspossible.Thedeviceusesa
systemtoconduct48-72houroldzebrafishthroughaliquidmedium(eggwater)
usingasyringepump.Thecompletesystemconsistsofthreemainsubsystems,
namelythepump,opticaldetectionandentrapmentcomponents.A3Dprinted
housingenclosestheelectricalcomponentsoftheentiresystem.Thedeviceworks
byaspiratingindividualfishthroughatubeviaapressuregradientcreatedwitha
syringepump.Eachcycleofthedeviceinvolvesthefollowingsteps:(1)loading,(2)
sensing,(3)trapping,(4)injection,and(5)flushing.Duringloading,asinglelarvais
extractedfromthereservoirandconductedthroughatubepasttheoptical
detectionsubsystem.Atthesensingstage,theopticaldetectionsubsystem
composedofaphotodiodeandalaser,sensestransmittedlightfromthelaserand
discernstheentryoflarvafromairbubblesanddebriswithprecision.Uponlarva
recognition,thespecimenisthenconductedtotheentrapmentdock(step3)where
itwillbeimmobilizedforinjection(step4).Thefinalstep(5)involvesconducting

thelarvaoutoftheentrapmentdockandsubsequentlyoutofentiresystemfor
furtheranalysis.Thisdevicewillprimarilyservezebrafishresearcherswhointend
tointroducevaccines,pathogensandotherexperimentalmaterialsintomany
individualzebrafishlarvae.

iii
ACKNOWLEDGEMENTS
Iwouldliketousethisopportunitytothankmythesisadvisor,Dr.Paul
Millard,forwelcomingmeintohislabandforbeingmymentoroverthepastfew
years.Throughhissupport,mygraspforthesubjecthasimprovedvastlyandIam
verygratefulforhispatienceandguidance.IwouldalsoliketothankDr.Doug
BousfieldandDr.SaraWaltonforprovidingvaluablecontributionstothiswork.
AdditionallyIwouldliketothankSeanMichaelTaylorfromtheIMRCforproviding
criticaldesigncontributionsandforhelpingmetobecomefamiliarwiththe
prototypingequipment.IwouldalsoliketoacknowledgeMadelineMazjanisfor
helpingwiththecontrolsoftwareandforconductingdevicetests.
Withagreatdealofgratitude,Iwouldliketothankmyfamilyfortheir
continuoussupportinmyendeavors.Theyhavealwaysbelievedinmeand
encouragedmetobethebestversionofmyselfandtonevergiveuponmydreams,I
willforeverbegratefulfortheirsupport.

iv
TABLEOFCONTENTS
ACKNOWLEDGEMENTS………………………………………………………………………………………iii
LISTOFTABLES…………………………………………………………………………………………………vii
LISTOFFIGURES……………………………………………………….……………………………………...viii
CHAPTER1. LITERATUREREVIEW…………………………………………………………………….1
1.1 InfluenzaVirusOverview…………………………………………………………………………..1
1.2 InfluenzaVirusVariants…………………………………………………………………………….1
1.2.1 Epidemiology-IAV………………………………………………………………………….2
1.2.2 IAVTransmissionandViralPropagation………………………………………….3
1.2.3 ClinicalFeaturesofinfection…………………………………………………………...5
1.3 CurrentIAVResearch……………………………………………………………………….………..5
1.4 AnimalModels…………………………………………………………………………………………..6
1.4.1ZebrafishasAnInfluenzaAVirusModel………………………………………….7
1.4.2PathologicalPhenotypeObservedinIAVInfectedZebrafish……..……10
1.5 SystemicIAVInfection……………………………………………………………………………..11
1.5.1 α2,6-linkedSialicAcidsinZebrafish……………………………………………..14
1.6 ZebrafishAntiviralDefenses…………………………………………………………………….16
1.7 InjectionofZebrafishEmbryos………………………………………………………………...17
1.7.1 StandardTechniques…………………………………………………………………….21
1.7.2 RapidPrototypingandInfluenceofTechnologyonResearch………….21
1.7.3 NotableDevicesforZebrafishInjection………………………………………….22

v
CHAPTER2. METHODS…………………………………………………………………………………….27
2.1 Overview………………………………………………………………………………………………..27
2.2 PumpSubunitComponents……………………………………………………………………..29
2.3 OpticalDetectionSubunitComponents……………………...……………………...……..31
2.4 EntrapmentDock………………………………………………………...………………………….32
2.4.1 Biocompatibility…………………………………………………………………………...37
2.5 SpecificationsandParameters…………………………………………………………………38
2.6 Electronics……………………………………………………………………………………………...38
2.6.1 ControlMaps/SoftwareProgramming…………………………………………..42
CHAPTER3. EXPERIMENTALRESULTSANDDISCUSSION…………………………………46
3.1 InputReservoirEfficacyTest…………………………………………………………………...46
3.1.1 Procedure…………………………………………………………………………………….46
3.1.2 Results…...…………………………………………………………………………………….46
3.2 OpticaldetectionUnitEfficacyTest………………………………………………………….48
3.2.1 Procedure…………………………………………………………………………………….48
3.2.2 Results…………………………………………………………………………………………49
3.3 EntrapmentDockTests…………………………………………………………………………...50
3.3.1 Procedure…………………………………………………………………………………….50
3.4 AverageDurationofScreeningSteps(1Cycle)……………………….…………………51
3.4.1 Procedure…………………………………………………………………………………….51
3.5 QuantitativeAssessmentofAnimalHealth……………………………………................53
3.5.1 Procedure…………………………………………………………………………………….53
3.5.2 Results……………………..…………………………………………………………………..53

vi
CHAPTER4. SUMMARY,CONCLUSIONANDFUTUREWORK……………………………...56
4.1 Summary……………………..………………………………………………………………………….56
4.2 Conclusions……………………..………………………………………………………………………56
4.3 FutureWork……………………..……………………………………………………………………..57
BIBLIOGRAPHY………………………………………………………...………………………………………..60
APPENDIXA:ZEBRAFISHHUSBANDRY………………………………………………………………71
APPENDIXB:VALVE/SYRINGEPUMPCONTROLMAP…………………………………………73
APPENDIXC:MATERIALSLIST…………………………………………………………………………...75
BIOGRAPHYOFTHEAUTHOR…………………………………………………………………………….77

vii
LISTOFTABLES
Table1. Averagedurationofscreeningstepsfromthereservoirtothelight
detectoratdifferentflowrates.(0.08cminnerdiameterand88cm
length)………………………………………………………………………………………….52
Table2. Calculateddurationofscreeningstepsfromthephoto-detectortothe
entrapmentdockatdifferentflowrates.(0.08cminnerdiameterand
80cmlength)………………………………………………………………………………...52
Table3. Averagetimerequiredforinjection……………………………………………….52
Table4. Calculateddurationofscreeningstepsfromtheentrapmentdockto
theexit(oftheentiresystem)atdifferentflowrates.(0.08cminner
diameterand62cmlength)…………………………………………………………...52

viii
LISTOFFIGURES
Figure1. Processofinfluenzavirusinfection………………………………………………..13
Figure2. Atypicalmicromanipulatorformanualinjectionneedleoperationin
allthreeaxes(x,yandz)……………………………………………………………….20
Figure3. Deviceschematicanddesign…………………………………………………………28
Figure4. Animageoftheinputreservoir……………………………………………………..29
Figure5. AquaCulturesingleoutletaquariumairpump……………………………….30
Figure6. NewEraNE-1000xbidirectionalprogrammablesyringepump………31
Figure7. Animageoftheopticaldetectionunitcomponents………………………...32
Figure8. Imageoftheentrapmentdockwithhighlightedchannelsand
dimensionsinmillimeters……………………………………………………………..33
Figure9. Imagesoftheentrapmentdockanditssupportingcomponents
designedusingsolidedgecadsoftware………………………………………….36
Figure10. Electroniccomponents………………………………………………………………….39
Figure11. Cole-Parmertwo-waynormallyclosedsolenoidpinchvalve…………..40
Figure12. Imageofthefunctionalcircuitdiagramofthevoltagecomparator….42
Figure13. LabVIEWcontroldashboard………………………………………………………….43
Figure14. Valve/syringepumpcontrolmap…………………………………………………..44
Figure15. Abargraphdepictingthenumberofzebrafishlarvaecollectedafter
60secatdifferentflowrates…………………………………………………………48
Figure16. Awaveformchartofthephotodiodevoltagespikeswithrespectto
time……………………………………………………………………………………………...50

ix
Figure17. Imageoftheentrapmentdockillustratingtheimmobilizedzebrafish
larvaeinthetwopossibleorientations…………………..………………………51
Figure18. Comparisonofmorphologicalabnormalitiesofexperimental
zebrafish,atdifferentflowrates,withacontrolzebrafish……………....54
Figure19. Quantitativeassessmentofzebrafishhealthatdifferentflowrates
(n=100)………………………………………………………………………………………..55

1
CHAPTER1
LITERATUREREVIEW
1.1 InfluenzaVirusOverview
Thecontagious,acuterespiratoryillnessknownasthefluhasafflicted
humansforcenturies,yetitspreciseoriginremainsamystery.Theworking
hypothesisisthatwildaquaticbirdsweretheprimaryreservoirforbothbirdand
mammalianvirusspecies.Thereisextensiveevidenceforthedirectandindirect
transmissionofinfluenzafrombirdstootherspeciessuchaspigsandhorses5.
InfluenzavirusesbelongtotheOrthomyxoviridaefamilyofRNAviruseswhichaffect
theupperandlowerrespiratorytract.Theyaresphericalorfilamentousandrange
insizefrom80to120nmindiameter6,7.Generalsymptomsofinfluenzainclude
fever,malaise,headacheandcough8.Thediseaseaffectsallagegroupsbuthashigh
prevalenceinpeopleunder25yearsandhasmoredrasticandlifethreatening
effectsininfantsandtheelderly9,7,10,11.Fluoutbreaksappearsuddenlyandcan
persistforafewdaystomonths,ifuncontrolled.Theseverityofanoutbreakmay
varydependingonthevariantofinfluenzaviruscausingthedisease.
1.2 InfluenzaVirusVariants
TherearesixgenerawithintheOrthomyxoviridaevirusfamily,andthree,
InfluenzaA,InfluenzaBandInfluenzaC,containvirusesthatcancauseinfluenzain
humansandbirds,aswellasothermammals12,13.Thesevariantsarecharacterized
bysegmentednegativestrand-RNAgenomesthataretypicallyenclosedinalipid
envelope.Subtleclassificationsdistinguishthethreesubfamilies.Viralsurface

2
proteinsserveasuniqueidentificationmarkersforviruses,andtypicalsurface
proteinsinclude:ahemagglutinin(HA),whichagglutinateserythrocytesandplaysa
keyroleinviralpropagation,neuraminidase(NA),whichisanenzymeessentialfor
thereleaseofprogenyvirusparticlesfromthesurfaceofaninfectedcell,and
membrane(M)proteins,whichserveasionchannels.
1.2.1 Epidemiology-IAV
ThemostprevalentformofinfluenzaisinfluenzaA,whichprimarilyaffects
humans,othermammalsandbirds.InfluenzaAisadiseaseofimmensecomplexity.
Itsoccurrenceandoutcomedependoninteractionsbetweenthevirus,whichis
constantlychangingitsgeneticandantigeniccomposition,andtheimmunesystem
ofthehost,whichmaynotbeabletorespondadequatelywithinarestrictedtime
frame.Theprincipalfactordeterminingwhetherinfluenzaoutbreaksoccuristhe
degreeofcomplementarypairingbetweenthesurfaceantigens(HAandNA)ofthe
virusandtheantibodiesagainstthem,whichareinthecurrentpopulation7.
Thisclassofvirusesisthemostlethalofthethree(InfluenzaA,InfluenzaBand
InfluenzaC),withtheabilitytocauseseverediseasesonthepandemicscale.
PandemicsofinfluenzaAhaveoccurredaboutthreetimespercenturysince
1700andweremanifestedbyglobalspreadofthedisease,typicallywithhigh
morbidityandmortality14.Themostextremepandemicrecordedinthe20thcentury
wastheSpanishinfluenza,prevalentbetween1918and1919,withadeathtoll
estimatedtohavebeenbetween20and40millionpeopleacrosstheglobe7,14,.Other
recentnotablepandemicsincludethe2009avianflu(H1N1)pandemic; thisvirus

3
wasauniquecombinationofinfluenzavirusgenesneverpreviouslyidentifiedin
eitheranimalsorpeople,infectingmillionsandkillingthousandsworldwide15.
Forseveraldecades,theCentersforDiseaseControlandPrevention(CDC)
hasmadeannualestimatesofinfluenza-associateddeaths,whichhavebeenusedin
influenzaresearchtodevelopinfluenzacontrolandpreventionpolicy.Outofthe5-
20%ofpeoplethatcontractthefluintheUnitedStateseachyear,theCDCestimates
over20,000laboratory-confirmedhospitalizationsannually,duetoIAVandother
relatedcomplications16,17.Inaddition,theeconomyisburdenedbyabout87billion
USDduetoIAVrelatedissues,rangingfrommedicalexpenses,paidworkabsences,
lossesinproductivity,declineinretailspending,andeventourism18.
1.2.2 IAVTransmissionandViralPropagation
Thefluishighlycontagiousandinfectedpeoplecanspreaditviaaerosolto
othersuptoabout6feetaway.Thevirusspreadsmainlythroughtinyaerosol
dropletsproducedwheninfectedindividualscough,sneezeor,occasionally,just
talk.Theaerosoldropletscontainingthevirusenterthemouth,noseorrespiratory
tractofpeoplewithinproximity.Inaddition,theviruscanbecontractedwhen
peoplecomeintodirectcontactwithsurfacesandobjectscontaminatedwiththe
virusandthenlatertouchtheirmouth,nose,oreyes.Contaminatedsurfacesmay
includedoorhandles,tablesurfaces,banknotes,andelectronicinputdeviceslike
keyboards.ResearchhasproventhatIAVcansurviveonsuchsurfacesforupto
eighthours19,20,21.Theflucanbecontagiousevenbeforesymptomsbecome
apparent.Mostadultscaninfectothersbeginningonedaybeforetheirsymptomsof

4
thediseasearedisplayedandcanremaincontagiousforuptooneweekafter
becomingsick19.Youngchildrenwithweakenedimmunesystems,however,canbe
contagiouswellpastaweek.
Uponinfection,thevirusreplicatesintheupperandlowerrespiratorytract,
reachingitspeakat2-3daysafterinfection.Symptomsofthediseasestartto
becomeapparentatthisstage19.Thevirusinducespathologicchangesthroughout
therespiratorytract,withthemostsignificantpathologyoccurringinthelower
respiratorytract.Bronchoscopiesofpersonswiththeflushowadiffuse
involvementofthetrachea,larynxandthebronchi,withacutemucosal
inflammationandedema.Thevirushasanaffinityforcellsintheepithelialliningof
therespiratorymucosa.Forviralpropagationtoinitiate,infectionofthesecells
mustoccur.Infectionbeginsbyvirusbindingtothesurfaceofthehostcell,followed
bythefusionofviralandhostcellmembranes.Theattachmentandfusionis
mediatedbyvirushemagglutininglycoprotein(HA)interactingandbindingtosialic
acidcellreceptors.Bindingtosialicacidoccursviaashallowcavitynearthedistal
tipoftheHAglycoprotein.Subsequently,influenzavirusentersthecellbyreceptor-
mediatedendocytosis22.SincethevirushasnoneedforDNAcoding,transcription
occurswithinthenucleus,followedby:replicationofviralRNAandsecondary
transcription,translationofviralRNAtoproduceviralproteins,thenculminating
withtheassemblyofviralstructuralcomponentsandthereleaseofprogenyvirus
intotheextracellularmedium.

5
1.2.3 ClinicalFeaturesofInfection
Inhumans,influenzaischaracterizedbythesuddenonsetofanacute
respiratoryillnessaccompaniedbyheadaches,chillsandnonproductivecough.This
isusuallyfollowedbymuscleaches,highfever,generalizedweaknessandlossof
appetite.Thefeverusuallydeclinesbythethirddayandisgoneafteraboutaweek,
butthecoughingandsignsofweaknesscanpersistforabouttwomoreweeks10.Due
tothevirus’natureofcompromisingtheimmunesystem,thehostcanbeproneto
infectionbyotherpathogensthatcanleadtofurthercomplications.Complications
oftheflucanincludeearinfections,sinusinfections,dehydration,andworseningof
chronicmedicalconditionssuchascongestiveheartfailureordiabetes.Moresevere
illnessessuchasprimaryinfluenzapneumoniacandevelop,whichcanbelethal.
Secondarybacterialpneumonia,amorerareformofthedisease,canalsooccurif
theflugoesuntreated23.Peopleover65yearsofage,thosewithheartconditions,
asthma,emphysema,AIDSorpeoplereceivingchemotherapyareatanincreased
riskofcomplicationsfrominfluenza.
1.3 CurrentIAVResearch
InfluenzaAviruses,duetotheirgeneticmakeupandmodeofpropagation,
arecontinuouslychanginginseveralanimalhosts,includingbirds,pigs,horsesand
humans24.Asaresultoftherapidandconstantemergenceofnewviralstrains,IAV
ishighlylikelytocausehumanepidemics.Itis,however,necessarythatnaturally
occurringIAVshouldbemonitoredandcharacterizedtoimprovethecurrent
understandingofthehosttropismaswellasthevirulenceofIAV.Therehavebeen

6
numerousresearchadvancementsinIAVresearchinthepastfewdecades,someof
whichinclude,butarenotlimitedto:viral/humansurveillanceandcharacterization,
influenzagenomediversityofevolution,vaccinationtechnologies,tissuepathology
andinfluenzaanimalmodelmanipulationtechnologies24,25,26,27,28,29,30.TheNational
InstituteofAllergyandInfectiousDisease(NIAID)InfluenzaResearchProgram
supportsresearchtolearnmoreaboutthestructureandpathogenesisofinfluenza
viruses.Suchresearchhasproventobeinvaluabletothedevelopmentofnew
vaccines,therapeutics,anddiagnostics31.Currentresearchinvolvesunderstanding
viralreplication,theemergenceofnewviralstrainsandtheeffectthosehaveonthe
immunesystem.AmajorNIAIDactivityistheInfluenzaGenomeSequencingProject.
Thisisajointefforttoobtaincompletegeneticfootprintsofthousandsofhuman
andavianinfluenzastrains,withthegoalofprovidingvaluablegenomicknowledge
thatcanbeusedtocreateimprovedpublichealthcountermeasures.AsofMay2014,
morethan16,000humanandavianisolateshavebeencompletelysequencedand
madepubliclyavailable32.
NIAIDalsosupportstheresearchcommunitybydevelopingnewanimal
modelsforpreclinicalevaluationofvaccineandtherapeuticcandidates.Animal
modelsareusefultoolsthatserveasbiologicalresourcesformicroarrays,clones,
peptides,andreagents.
1.4 AnimalModels
Animalmodelshavebeenpivotalinresearchfordecades,leadingtomajor
drugdiscoveriesandmedicalbreakthroughs33,34.Suitablemodelsareusually

7
selectedbasedonphysiological,anatomicalandgeneticresemblancewithhumans.
Theunlimitedsupplyandeaseofmanipulationaresomeofthegeneraladvantages
ofanimalmodels.Scientificresearchcansometimesinvolvestatisticalanalysisof
testspecimens,mostlybymanipulatingonlyonevariablewhilekeepingothers
constant,andthenobservingtheconsequencesofthatchange.Anadequatenumber
oftestspecimensare,however,requiredtoaccomplishthistaskeffectively.The
relativeeaseofmanipulatinglargesamplesizesmakesanimalmodelsinvaluableto
medicalresearch.
Rodentsarethemostcommontypeofmammalusedinexperimentalstudies.
Extensiveresearchhasbeenconductedusingrats,mice,gerbils,guineapigs,and
hamsters35,36.Amongtheserodents,themajorityofgeneticstudies,especiallythose
involvingdisease,haveemployedmicebecausetheirgenomesaresimilartothatof
humansandmanyuniquestrainshavebeendeveloped.Othercommon
experimentalorganismsincludefruitflies,baker'syeastandzebrafish37.
1.4.1 ZebrafishasAnInfluenzaAVirusModel
Animalmodelsofinfluenzaareessentialtoresearchthatfocusesongaininga
betterunderstandingofthevirusanditseffectonhumans.Therearecurrently
variousresearchapplicationsofanimalmodelsrangingfromthepathogenesisand
hostresponsetoIAVtoantiviraldrugscreeningsandpre-clinicaltestingofantiviral
drugsandvaccines.IAVanimalmodelsarealsousedtoinvestigatethecompetency
ofvaccinesanddrugstocombatthevirus,preventfuturecomplicationsand
commonly,toreducethesymptomsofinfluenza.Inselectingthemodelsfor

8
research,acoupleoffactorshavetobeconsidered.Themostimportantcriterionis
thesusceptibilityoftheanimaltoinfluenzavirusinfectionwhilesupportingviral
replication,andexhibitinghumanhistopathologicalandmorphologicalsymptomsof
aninfluenzavirusinfection38.
Zebrafish(Daniorerio)modelsforinfectiousdiseasesarewellestablishedfor
characterizationofbacterial,fungalandviralinfections39,40,41.TheNational
InstitutesofHealth(NIH)alongwithotherresearchbodieshasapprovedthe
zebrafishanimalmodelforuseintheadvancementofbiomedicalstudies.Someof
itsinherentadvantages,suchasopticaltransparency,minimalhusbandry,large
clutchsizeandfastreproductivecyclemakethezebrafishanidealspecimenfor
high-throughputdrugtesting.Anotherimportantfactortoconsideristhatzebrafish
arepropagatedattheoptimalincubationtemperatureforvirusesduringinfection
studies.Zebrafishcanbegrowninatemperaturerangewithinthatofthehuman
respiratorytract(from25to33°C).
Asaresult,thezebrafishmightbewellsuitedtomodelinghumanrespiratoryviral
infections,giventhesimilaritiesbetweenthezebrafishswimbladderandthehuman
lung42,43,44,45.Therehavealsobeenrecentstudiesofzebrafishswimbladder
infectionswithhumanfungalpathogens.Furthermore,zebrafishattheearlystage
ofdevelopmentarehighlyamenabletogeneticmanipulationandtransgenesis2.
Amongallofthebenefitsofthezebrafishanimalmodelsystem,oneofthe
mostimportantcharacteristicsthatmakethissystemidealforIAVresearchisthe
zebrafish’simmunesystem.Duringtheonsetofdevelopment,zebrafishareknown
torelysolelyontheinnateimmuneresponseforthefirst4to6weeks46,47,48.

9
Hematopoiesisinzebrafishproduceslargelythesamedifferentiatedcelltypesas
observedinmammals.Monocytes,neutrophils,macrophagesandeosinophilshave
allbeendescribedwithinthezebrafishmyeloidlineage47.Inaddition,othercells
withmammaliancytotoxicproperties,suchasmastcellsandnaturalkillercells,
havealsobeenclassifiedwithinzebrafish49.Theinnateimmunefunctionsofthese
cellsprovideaneffectivedefensesystemduringtheearlystagesofdevelopment50.
Themajorityoftheresearchconductedusingthezebrafishmodelinvolves
fish-specificviralpathogensthathavetheabilitytoinfectandcausedisease;
however,fewhumanviruseshavebeendemonstratedtoinfectthezebrafish51.
Somerecentstudieshaveproventhezebrafishtobesusceptibletoviralinfectionby
mammalianviruses,ascharacterizedbyBurgosetal,wherethezebrafishwasused
asamodelofherpessimplexvirustype1(HSV-1)infectionofthenervoussystem52.
Gaboretal,attheUniversityofMainedescribedazebrafishmodelforhuman
influenzaAVirus(IAV)infectionandshowedthatzebrafishembryosaresusceptible
toinfectionwithbothinfluenzaAstrainsAPR8andX-31(Aichi).Influenza-infected
zebrafishshowedanincreaseinviralburdenandmortalityovertime.The
expressionofinnateantiviralgenes,thegrosspathologyandthehistopathologyin
infectedzebrafishrecapitulateclinicalsymptomsofinfluenzainfectionsinhumans2.
Thestudywasconductedbasedonthepremisethatanantiviralstatesimilartothat
inhumanswasinducedininfectedspecimens,leadingtothedeductionthat
zebrafishcanbeasuitablealternativetovertebrateanimalmodelsusedforIAV
studies.RecombinantinfluenzavirusescarryingaGFPreportergeneintheNS
segment(NS1-GFP)wereusedtocharacterizeinfectioninthezebrafish.Although

10
attenuatedcomparedtothewildtypevirus,itistypicalforthesemodifiedviral
strainstoreplicateefficiently53.Todeterminewhetherthemodifiedhumanisolates
ofIAVcouldcausediseaseinzebrafishembryos,differentstrainsofIAV[APR8
(H1N1)andX-31(H3N2)]wereinjectedintravenouslyinto48hpf(hourspost
fertilization)zebrafishthroughtheDuctofCuviertomimicviralinfectioninthe
humanbloodstream.Aninertphosphatebufferedsalinesolutionwasalsoinjected
intoanumberofzebrafishembryosinacontrolsetup2.
DatafromthestudydemonstratedthatIAVreplicateswithinzebrafishand
suggeststhatthedeathofIAV-infectedzebrafishislikelytobecausedbyIAV
infection.Toinvestigatetheeffectoftreatmentonthezebrafishimmunesystem,an
antiviralcompoundusedforthetreatmentofIAVinhumanswasintroduced,which
reducedthemortalityoftheinfectedfishaswellasthevirulenceofIAV.
1.4.2 PathologicalPhenotypeObservedinIAVInfectedZebrafish
Inanefforttocharacterizetheinnateimmuneresponseofzebrafishtothe
presenceofeitherstrainofIAV(APR8 or X-31)inthebloodstream,infected
embryosweremonitoredovertimeforbehavioralandphenotypicchanges2.A
controlledsetofembryoswereinjectedwithPBS(phosphate-bufferedsaline)and
monitoredunderidenticalconditions.Whilefishinthecontrolsetupdisplayedno
abnormalbehaviororsignsofviremia,theIAV-injectedzebrafishexhibited
symptomsoflethargy,withsignsofedemaobservedinthepericardiumandyolk
sac.Furthermore,theedemaworsenedovertimeandadditionalpathological
phenotypes,includingvaryingdegreesoflordoticcurvatureofthespine,were

11
observed.Otherobservationsincludedpigmentationdefects,cranialirregularities
andeyedeformities.Gaboretalextendedtheinvestigationtofurthercharacterizetheinnate
immuneresponseofzebrafishtoIAVinfectionbyconductingpathologicalanalysis
onthecellularlevel.Influenzavirustypicallyreplicatesintheepithelialcells
throughouttherespiratorytractofhumansand,theviruscanberecoveredfrom
boththeupperandlowerrespiratorytractofindividualswhohavebeennaturally
orexperimentallyinfected.IAVviremiainitsacutestagecanbecharacterizedby
multifocaldestructionanddesquamationofthepseudostratifiedcolumnar
epitheliumofthetracheaandbronchi29.Itisalsotypicalforthebasallayerofthe
epitheliumtobetheonlystructureremaining.Edemaandcongestionofthe
submucosacanalsobeobserved.
Inthecaseofthezebrafish,Gaboretal.preparedtissuesamplesfromIAVinfected
zebrafish48afterinjection.Analysisofthetissuesconfirmedtheclinicalsymptoms
ofinfluenza,includingnecrotictissuesandedema.
1.5 SystemicIAVInfection
Thevascularsystemallowstheflowofbloodfromthehearttoallpartsofthe
body,supplyingoxygentothemostvitalorganssuchasthekidneys,thebrainand
theliver.Sincebloodwithinthissystemisdistributedthroughoutthebodyfrom
theaorta,introductionofabloodbornepathogensuchasIAVintothevasculature
expeditesviralinfectionthroughoutthebody.IAVischaracterizedbyhemagglutinin
(HA),anantigenicglycoproteinusuallyfoundontheirsurfaces.Theseglycoproteins

12
areresponsibleforbindingthevirustothecellthatisbeinginfected54.Theviral
hemagglutininproteinsbindtosialicacidgroupsofcellularsurfaceglycoproteinsto
achieveviralattachmentandentry.ThespecificityoftheHA-Sialicbindingcomplex
isoneofthemajordeterminantsforcontrollingviraltropismandhostspecificity
Thisexplainswhysomeorganismsaresusceptibletocertaindisease-causing
pathogens,whileothersarenot.
Typically,humaninfluenzaviruseshaveabindingpreferenceforα2,6-linked
sialicacid.Avianinfluenzavirus,ontheotherhand,bindstotheα2,3-linkedsialic
acidofthehostbird56.Besidestheconventionalviralinfectionmechanism,thevirus
cansometimestraveltoacidicendosomesformembranefusionviaclathrinor
caveolinmediatedendocytosis57.Alternatepathwaysintothecellbyviruseshave
alsobeenstudied,demonstratingthatinfluenzaviruseshavetheabilitytoemploy
differentcellentrymechanismstoachieveviralinfection.Itis,however,notknown
whetherallinfluenzaviruseshavetheabilitytousetheothervariouspathwayswith
identicalpreferences56,58,59,60.FollowingtheinteractionbetweentheviralHAand
theα2,6-linkedsialicacid,endocytosisoftheviralparticleistriggered.Figure1
demonstratesatypicalprocessofcellentryoftwodifferentviruses:Influenzaand
HIV.

13
Schematic diagrams of HIV-1 and influenza virus fusion and entry processes. MajorconformationalchangesintheviralenvelopeglycoproteinsoccurforfusionofbothHIV-1andinfluenzavirus,buttherequirementsforfusiondiffer,asdotheviraltargettypes(notshown).ForHIV-1,gp120binds sequentially to its primary receptor CD4 and, after an initial conformational change, to co-receptors(chemokinereceptor)CCR5orCXCR4(notshown)(step1a).Co-receptorinteractiontriggersfusionoftheviralandcellularmembranes,initiatedbytheHIV-1fusionpeptidethatislocatedingp41(step2a).FusionandentryoftheHIV-1genomicRNAandaccompanyingviralproteinsintothetargetcelloccursatthecellsurfaceatneutralpH.Followingentry,theHIV-1RNAgenomesaretranscribedbyreverse transcriptase intoDNA(step3a)andtheHIV-1pre-integrationcomplex is transportedto thenucleusforintegrationintotargetcellgenomicDNAtoinitiatechronicinfection.OndifferentcelltypesfromthoseaffectedbyHIV-1,influenzavirusbindsviahaemagglutinin1(HA1)toterminalsialicacidspresentonglycoproteinsoronglycolipids(step1b).Thevirusissubsequentlyinternalizedbyreceptor-mediatedendocytosisintoalowpHcompartment(endosome),triggeringconformationalchangesthatexpose the viral fusion peptide that is located in HA2 (step 2b). Subsequently, the genomicribonucleoproteincomplexistransportedtothenucleustoinitiatetranscriptionandreplicationoftheviralgenome(step3b).
Figure1:Processofinfluenzavirusinfection.
Karlssonetal.SchematicdiagramsofHIV-1andinfluenzavirusfusionandentryprocesses.Diagram.ThechallengesofelicitingneutralizingantibodiestoHIV-1andtoinfluenzavirus.Fall2008;6(2):148.AvailablefromNatureReviewsMicrobiology.AccessedMay62017.

14
Influenzavirusbindstoterminalsialicacidspresentonglycoproteinsor
glycolipidsviaHA1.Thevirusissubsequentlyinternalizedbyreceptor-mediated
endocytosisintoanendosome.Thisinitiatesconformationalchangesthatexpose
theviralfusionpeptidelocatedonHA2.Theentireviralproteincomplexisthen
transportedtothenucleus,wheretranscriptionandreplicationoftheviralgenome
willoccur
ReplicationandtranslationoftheviralRNAproducesnewviral
macromolecules,whichareultimatelyassembledintocompleteviralparticlesfor
releaseandinfectionofnewcells62.
1.5.1 α2,6-linkedSialicAcidsinZebrafish
Thepresenceofcompatiblesialicacidmodificationsonhostglycolipidsor
glycoproteinsisoneofthemaindeterminantsofviralinfectioninmostanimal
species.Thepresenceofα2,6-linkedsialicacidinthezebrafishmakesthisa
valuablemodelsystemforthestudyoftheinfluenzaAvirus.Inspiteofthis,there
arenoknownreportednaturallyoccurringviralinfectionsofthezebrafish63.Inan
efforttodeterminewhetherzebrafishembryoscouldserveasahostforinfection
withmammalian/humanisolatesofIAV,Gaborconductedastudytoanalyzethe
sialicacidlinkagetypespresentonzebrafishglycoproteinsorglycolipids2.Analysis
usingHighPerformanceAnionExchangeChromatographywithPulsed
AmperometricDetection(HPAEC-PAD)demonstratedthatα2,6-linkedsialicacids
werepresentinzebrafishembryostwodaysafterfertilization.Therewere,
however,noα2,3-linkedsialicacidsdetectedinthetwoday-oldembryos2.There

15
areanumberofchemicalvariantsofsialicacids,andinfluenzastrainsvaryintheir
affinityforthem.Terminallylinkedsialicacidscanoccurinα-2,3,α-2,6,orα-2,8
linkages.Eventhoughthestudywasnotintendedtoidentifyα2,8-linkedsialic
acids,othershavepreviouslydemonstratedthatzebrafishhavetheabilityto
synthesizeavarietyofsialylatedglycoconjugatesduringtheonsetoftheir
development64,65,66.Theseuniquecharacteristicsofthezebrafishmodelsystem
makeitanattractivecandidateforstudyinginfectiousviraldiseases.Withfurther
studiesintosialylatedglycoconjugatespresentatvariousstagesofdevelopmentof
thezebrafish,additionalhumanviraldiseasescanfurtherbecharacterized.Someof
theknownhumanviralpathogensthathaveanaffinityforα2,6-linkedsialicacids
includecertainmembersofthefollowingvirusfamilies:
coronaviridae,paramyxoviridae,caliciviridae,picornaviridae,reoviridae,polyomavi
ridae,adenoviridaeandparvoviridae.Mostofthesegroupscontainmajorpathogens
forhumansthataffectallpartsofthebody,rangingfromsmallorganslikethe
salivarygland(inthecaseofmumps)tocellularnetworksasintricateasthe
immunesystem2,67,68.
Avianinfluenzavirusstrainshavethetendencytobindtosialicacids
attachedtogalactoseviaanα2,3-linkage,whichisthemajorsialicacidonepithelial
cellsoftheduckgut. Ciliatedepithelialcellsinthehumantrachealack α2,3linked
sialicacids,suggestingthatthelackofasuitablereceptoraccountsfortheinefficient
replicationofcertainhumanvirusesinduckintestineandofcertainavianvirusesin
humans55,69.

16
1.6 ZebrafishAntiviralDefenses
Althoughthezebrafishanimalmodelhasbeenproveneffectivefortesting
medicaltreatmentsandvirologicalassays,itcanserveasanidealtoolfor
understandinghostbiologicalresponsesafterviralinfections,particularlythe
InfluenzaAvirus.Aclearandthoroughunderstandingofahost’simmunity(both
innateandadaptive)canprovideinsightintonewtreatmentsandeffective
therapies.Thezebrafish,likemanyorganismspossessesinnateandadaptive
immunity,howeverthezebrafishreliessolelyonitsinnateimmuneresponse
systemforthefist4-6weeksofdevelopment2.Withinthisperiodofgrowth,
zebrafishdevelopfromtheembryotothelarvaestage.Thistemporalseparationof
innateandadaptiveimmunityinzebrafishallowsforthestudyoftheinnate
immuneresponsetoviralinfectionindependentofadaptiveimmunity.Uponviral
attack,oneoftheprimaryprocessesinitiatedistheinductionofhostantiviralstate,
whichslowsdowntherateofprogressionofviremia2.Themajormolecule
responsiblefortheantiviralstateisinterferon(IFN).Cellsthatarealreadyinfected
withavirussecreteinterferonstopreventfurthervirusexposureofhealthycells.
ResponsetoIFNinvolvesarapidanddirectsignaltransductionmechanismthat
quicklyreportsthepresenceofextracellularcytokinestothecellnucleus.This
preservesthespecificityinherentincytokine-receptorinteractionstoinduce
expressionofasetofgenesviatranscription,leadingtoencodingofimportant
antiviralproteins.Establishmentoftheresultingantiviralstateprovidesacrucial
initiallineofdefenseagainstviralinfection.StudiesofIFN-deficientcellsand
animalsderivedbygenetargetinghavedemonstratedtheessentialnatureofIFN-

17
mediatedinnateimmunity70,71,72.
Allbloodcellsinhumansbeginashematopoieticstemcells,andthendifferentiate
intomyeloidcells(erythrocytes,megakaryocytes,monocytes,neutrophils,
basophils,oreosinophils)orlymphoidcells(T-lymphocytesandB-lymphocytes).
Theseimmunecellstypicallyperceiveandrespondtomicrobialstimuli.Zebrafish
arelargelysimilartohumanswithrespecttocelldifferentiation.Theyhavedistinct
stagesofhematopoiesisthatoccurindiscreteyetfunctionallyanalogoussites.
Zebrafishmyeloidlineages,includingmonocytes,macrophages,neutrophilsand
eosinophils,havebeenreviewedandhavebeencharacterizedassimilartothosein
humans47.Theinnateimmunerolesofthesecellsofferanefficientdefense
mechanismduringtheonsetofdevelopment.Inthecaseofaviralinfectionsuchas
influenza,theimmunecellsaretriggeredandfunctionaccordinglytoquarantinethe
virusbyinducingtheantiviralstate.Macrophagesandneutrophils(large,mobile
phagocyticcells)canalsoberecruitedtothesiteofinfectiontoengulfinvading
species.Macrophageshavebeenknowntorespondmorequicklytoinfectionthan
neutrophilsatabout30hpf73.Therelativelyslowerneutrophilsrespondatabout
52hpftocomplementactivitiesofthemacrophages,partlybecausethephagocytic
capabilityofneutrophilsisrelativelylowcomparedtothatofmacrophages74.
Precursorsofthelymphoidcells(T-cellandB-cells)begintheprocessof
rearrangementandrecombinationofthegenesforimmunoglobulinandTcell
receptormoleculesbyday4.Thisistheonsetofadaptiveimmunesystem
development.Thethymus,whichisalsoalymphoidorgan,producesTcellsandB
cellsforthezebrafishimmunesystem,thepancreashasalsobeenreportedto

18
produceBcells75,76.Eventhoughtheadaptiveimmunesystembeginsformingearly
inzebrafishdevelopment,studieshaveshownthatthezebrafishasincapableof
mountingafullantibodyresponseuntilearlyadulthood,whichisabout4weeks
postfertilization.Themodeofpathogenicdefenseupuntilthispointisexclusively
dependentontheinnateimmunesystem77.
Inanefforttounderstandthedifferentmaturationstatesoftheimmune
systemduringzebrafishdevelopment,S.H.Lametalinvestigatedtheexpressionof
siximmune-relatedgenesofthezebrafishnamely:Ikaros,thatencodesa
transcriptionfactorwhichisusedasanearlylymphoidmarker;Rag-1
(Recombinationactivatinggene-1),whichencodesaproteininvolvedingenomic
rearrangementoftheT-cellreceptor,immunoglobulinloci,andisasuitablemarker
formaturinglymphocytes;T-cellReceptorAlphaConstantregiongenes(TCRAC)
whichencodeportionsoftheantigenreceptorsofmatureT-lymphocytes;three
immunoglobulinlightchainconstantregiongenes(IgLC)whichencodeportionsof
theantigenreceptorsofmatureBlymphocytes.Althoughtherehavebeencloning
andcharacterizationstudiesonimmune-relatedgenespriortothisstudy78,79,there
wasverylittleknownaboutthematurationofthezebrafishimmunesystemwith
regardtoitsformandfunction.Byemployingstandardprofilingtechniquesto
assessgeneexpression(quantitativereal-timepolymerasechainreaction,insitu
hybridization(ISH),immuno-affinitypurificationandWesternblotting),Ikaroswas
detectedfirst,at1dpf.Theexpressionthereafterincreasedgraduallytomorethan
two-foldbetween28and42dpfbeforedecreasingtolessthantheinitial1dpf
expressionlevelinadultfish(aged105dpf).Rag-1expressionlevelsincreased

19
rapidly(by10-fold)between3and17dpf,reachingamaximumbetween21and28
dpfbeforedecreasinggradually.Inadultfishaged105dpfhowever,theexpression
levelofRag-1droppedsignificantly,andwasequivalenttotheexpressionlevelat3
dpf.T-cellreceptoralphaconstantregiongenesandmRNAsofimmunoglobulin
lightchainconstantregion(IgLC)isotype-1,2and3weredetectedatlowlevelsby3
dpfandtheirexpressionlevelsincreasedsteadilytotheadultrangebetween4and
6weekspost-fertilization(wpf).Usingtissue-sectionISH,Rag-1expressionwas
detectedinheadkidneyby2wpfwhileIgLC-1,2and3weredetectedinthehead
kidneyandthethymusby3wpfonwards.SecretedIgwasonlydetectableusing
immuno-affinitypurificationandWesternblottingby4wpf.Ahumoralresponseto
T-independentantigen(formalin-killedAeromonashydrophila)andT-dependent
antigen(humangammaglobulin)wasobservedinzebrafishthatwereimmunizedat
4and6wpf,respectively,indicatinganormalimmuneresponse.Thefindings
revealedthatthezebrafishimmunesystemismorphologicallyandfunctionally
matureby4–6wpf77.
1.7 InjectionofZebrafishEmbryos
Oneofthemostcommontechniquesusedtointroducematerialsinto
zebrafishembryosandlarvaeisinjection.Althoughinvasive,itallowsprecise
targetingoftissuesandorgans.Whenconductedunderappropriateconditions,such
asmildsedation,injectionsposeminimalrisktozebrafishlarvae.Duringinjection,
certainsubstancessuchasnucleicacids,proteins,ordrugsareintroducedintothe
zebrafish,usuallyattheearlystagesofdevelopment80.Injectionsaretypically

20
conductedwithglassneedlesfilledwiththematerialtobedelivered.Theglass
needlescanbefabricatedeasilybycarefullydrawingoutaheatedcapillarytube
withtheappropriateinnerandouterdiameter81.Theglassneedlesarethenheldby
amicromanipulatorthatcontrolsandchangesthepositionoftheneedlerelativeto
theembryowithhighprecision.Pressurefromcompressedairisusedtoreleasethe
contentintotheembryoorlarva.
Figure2:Atypicalmicromanipulatorformanualinjectionneedleoperationinallthreeaxes(x,yandz).Itallowsforcoarseadjustmentreadingswithanaccuracyof100micrometers.WarnerInstruments,ModelMM-33,HamdenCT.
Injectionallowsrapiddeliveryofmaterialintothezebrafish.Aftermanipulationof
embryosorlarvaeintoasuitablearray,anexperiencedresearchercaninjectseveral
hundredembryosinonehour,althoughtherelativelyhighchancesoferror
associatedwiththistechnique,cannegativelyaffecttheconsistencyofinjection.A
detailedinjectionprotocolconsistingofzebrafishloadingandpreparationhasbeen
characterized82.

21
1.7.1 StandardTechniques
Injectionofzebrafishhasbeencommonplacefordecadesandnumerous
procedureshavebeendevelopedtoinjectalmosteverypartofthelivefish,notably
thebrainventricle,thekidney,theswimbladderandtheDuctofCuvier(DC).The
DCisawidebloodcirculationvalleyontheyolksacconnectingthehearttothe
trunkvasculature.Injectionthroughthisductallowsquickandefficientsystemic
infectionofzebrafishandisparticularlyusefulforintroductionofbacteriainto
embryosbetween1and3dpf.Fortheseinjections,theneedleisinsertedintothe
startingpointoftheductofCuvierjustdorsaltothelocationwheretheductstarts
broadeningovertheyolksac.Thislocationisthedeepestsectionoftheductand
thereforeprovidesthelowestriskofpuncturingtheyolksac.Theinjectedmaterial
willfollowthebloodflowthroughtheductofCuvier,overtheyolksactowardthe
heart,andcanbemonitoreddirectlypost-injectionbymonitoringtheexpanding
volumeoftheduct83.
1.7.2 RapidPrototypingandInfluenceofTechnologyonResearch
Rangesoftechnologicaladvancementshavepavedthewayfornewand
innovativedevicesandtechniquesthathelpmakelaboratoryworkmoreefficient
andreliable.Therateoftechnologicaladvancementhasbeenhighwithinthispast
decade,whichmaybeattributableinparttotheincreaseincomputationalpower84.
Anothermajorinfluenceontechnologicaladvancementistheemergenceof3D
printingtechnologiesforrapidprototyping,whichispoisedtorevolutionizedevice
developmentinmanyindustries,includinghealthcare.Sinceitsinitialuse,additive

22
prototypingtechnologyhasadvancedtouseinawiderangeofapplications
includingtissueengineering,dentistry,construction,automotiveandaerospace
85,84,86.Thetechnologyisalreadychangingthewayobjectsaremade,withresulting
productsrangingfromtoolsandtoystoclothingandevenbodyparts.3Dprintingis
partofaprocessknownasadditivemanufacturingwhereanobjectiscreatedby
addingmateriallayerbylayer,usuallyatafractionofthecostandtimeofstandard
meanssuchasforging,moldingandsculpting.Inthemedicalworld,physiciansare
testingbiomaterialsforregenerativemedicine.Usingmedicalimagingand3D-
modelingtechnology,surgicalteamscannowalsouse3Dprinterstocreate
temporarytoolsthatareaffixedtotheskeletalstructureofthepatienttoprovidea
precise"blueprint"forreshapingbonestructuretoperfectlyaccommodate
standard-sizeimplants.Custom-printeddrillingguidespermitscrewstobeplaced
preciselytoensurethebestfitwithapatient'sbody87,88.
Withinthedynamicbranchofzebrafishresearch,prototypingtechnologies
aregraduallybeingintroducedtoimprovethespeedandqualityoflabwork30,92,97.
Thesetechnologiesmayhavethepotentialtomakecomplexandtedious
procedures,suchasinjectionandimagingofchorionatedembryos,relativelysimple.
Thefollowingparagraphhighlightsdevicesthatimplementrapidprototyping
techniquestomakeworkingwithzebrafishmoreefficient.
1.7.3 NotableDevicesforZebrafishInjection
Pardo-Martinetal.atMassachusettsInstituteofTechnologyandHarvard
Universitydevelopedahigh-throughputplatformforcellular-resolutioninvivo

23
pharmaceuticalandgeneticscreensinzebrafishlarvae30.Theautomatedsystemis
capableofsamplingzebrafishlarvaeandthenpositioningthemforimaging.The
systemisalsocapableofmanipulatingbothsuperficialanddeeporgans(via
femtosecondlasermicrosurgery).Eachcycleofthesystemincludesthefollowing
steps:loading,sensing,trapping,injectionandflushing.Duringloading,thesystem
extractslarvaefromareservoirfilledwithdistilledwater.Ahigh-speedoptical
detectionsystemcomposedofaphotodiodeandtwoLEDsdetectstheentryof
larvaeintotheloadingtube.Thephotodiodesensestransmittedandscatteredlight
fromthelaserlightsource.Bymonitoringthelightsignals,thesystemcan
distinguishalarvafromairbubblesanddebriswith99.99%reliability(n=100).
Afterloadinganddetection,thelarvamovesintoacapillarypositionedwithinthe
angleofviewofanopticalimagingandmanipulationsystem.Usingafastcamera
andanautomatedimage-processingalgorithm,thelarvaiscoarselypositionedby
thesyringepumpwithinfocusoftheopticalimagingsystem.Attheendofone
completecycle,animalscanbedispensedbackintoeitherindividualwellsorlarger
containersbyexecutingtheloadingprocessinreverse.Whilethisdeviceboastsof
effortlesszebrafishmanipulationforimaging,itfallsshortofaddressingthetedious
processofmicroinjection:afrequentandnecessarysteppriortoimaging.Designof
anentrapmentmechanismaspartofasamplingdevicetoaccommodateaninjection
needlewouldaddanimportantmanipulationcapability.
RohdeetalatMassachusettsInstituteofTechnologydevelopedahigh-speed
microfluidicsorterwiththeabilitytoisolateandimmobilizesmallliveanimalsina
well-definedgeometry(withinachip)forscreeningphenotypicfeaturesin

24
physiologicallyactiveanimals.Theintegratedchipcontainsindividually
addressablechambersthatcanbeusedforincubationandexposureofindividual
animalstobiologicalcompoundsandhighresolutionimagingwithouttheneedfor
anesthesia.Besidesitssortingandimmobilizingfunction,thechannelswithinthe
microfluidicchipcanbecombinedinvariousconfigurationstopermitamultitudeof
complexfunctionssuchassmallanimalincubation(eg.C.elegans),immobilization
andindependentsubcellularresolutionimaging89.Thissystemisversatilebut
sharesasimilarlimitationwiththesystemmadebyPardo-Martin30andhisteam:it
doesnotaddresstheupstreamprocedureofanimalinjection.Furthermore,thehigh
multifunctionalcapabilitiesofthissystemmightmakeitsusceptibletocross-
contaminationduringuse.Boththemechanicalandelectricalcomplexityofthe
systemalsoraisesdurabilityconcerns.
BischeletalatUniversityofWisconsin-Madisondesignedazebrafish
entrapmentdevicethatcanbeusedtoquicklyandrepeatedlypositionzebrafish
embryosinapredictablearraywiththehelpofapipette.TheZebrafishEntrapment
byRestrictionArray(ZEBRA)iswellsuitedforusewithautomatedmicroscope
stages,therebyreducingtheamountofimagingtimeandfurtherincreasing
throughputcomparedtotraditionalmethods. ZEBRAwasdesignedandoptimized
toimmobilize3–5dpfzebrafishembryosinapredictablearraywithouttheuseof
agarose.Thedeviceconsistsofanenclosedchannelwitharestrictionwidththatis
accessedthroughaninputandanoutputport90.Fornormalfunction,ZEBRAmust
befilledwithwater(thisprocessrequiresoxygen-plasmatreatmenttorenderthe
insideofthechannelshydrophilic).Afterinitialfilling,apipettecanbeusedtoadda

25
dropletofwatercontainingaselectedzebrafishembryototheinputportofthe
device.Themainfunctionofthedeviceisgovernedbypassivepumping:a
phenomenonbywhichsurfacetensioninducespressuredifferencestodrivefluid
movementinclosedchannels91.Asaresult,waterisdrivenfromthesmallerinput
port(withhighersurfacetension)tothelargeroutputport(withlowersurface
tension).Thezebrafishisthentrappedinplaceuponreachingthechannel
restriction.Userscanchoosewhethertoloadthezebrafishintothedeviceheadfirst
ortail-firstdependingonthedesiredapplication.Passivepumpingworks
reproduciblyaslongasthereisaclosedchannelbetweenaninletandanoutlet91.
Thismakesthedevicemorepracticalandportablesinceitisnotdependentona
powersource.Despiteitsinherentadvantages,thedevicedoesnotaccommodate
zebrafishinjection.Theflowrateandaspirationconditionswithinthechannels
appeartobedependentonthesizeofthedropletplacedattheinletofthedevice.As
thedropletcollapses,theaspirationratedecreasesthuscreatingadeviceconstraint
thatcanpotentiallylimitthelengthofthechannelbetweentheinletandtheoutlet
ports91.Althoughlessenergy-efficient,employingactivepumpinginthisscenario
canpotentiallyeliminatethisconstrainttopermitlongerchannelsthatcan
accommodatezebrafish,aswellasotheranimals,suchasC.elegans.
WesthoffetalatUniversityofHeidelbergformedapieceofbrasstoserveas
anegativemoldtocastagarosetoyieldaseriesofwellscapableofholdingzebrafish
forimagingandmicromanipulation92.Thebrassmoldcastsindentationsintoan
agarosegelbyinsertingthetipsofthemoldintounsetagarsolutionandallowing
thesolutiontogel.Themoldisremoved,leavingvoidsthatarecontouredtohold

26
zebrafishlarvaeforimaging.Whilethisdevicerequiressignificantmanual
interfacing,itminimizestheissueofhavingtoinsertandorientspecimensinan
agarmedium.Userssimplypipettetheirspecimenintothewellspriortoimaging.
Evenlyspacedwellsallowscreeningoflargenumbersofspecimensandcanbe
automatedwithatranslationstage.Whilethedeviceoffersasimplewayforusersto
probeandorganizelargequantitiesofspecimens,itleaveslittleroomfor
automationoftrans-locatingfishintothedevice.Asaresult,moreuserinteractionis
neededtomoveandorganizethespecimens.Automatingthisprocesswouldallow
forminimaluserinteractionbetweentheuserandthedevice.
Thedevicesdiscussedabovealldemonstrateinnovativeapproachesto
eliminatingcommonproblemsofworkingwithzebrafish,suchasmanipulation;
however,noneofthemisapplicabletotheprincipalfunctionofzebrafishinjection.
UsingthezebrafishasananimalmodelforIAVstudiesinvolvessystematicallyand
reliablyintroducingthevirusintoindividuallarvaeviainjection.Adeviceto
conductandimmobilizezebrafishspecimensforinjectionwillbeinvaluabletoIAV
studies,particularlywheresamplesizesareverylarge.Thisprojecthasbeen
directedtowarddesignandfabricationofanautomateddevicethatwillhelpsolve
thisproblem.Themaingoaloftheproposeddeviceistoprovideameansfor
storingandgentlyconductingzebrafishfromareservoirtoachamberfor
immobilizationandinjection.Thiswillminimizedirectinteractionbetweentheuser
andspecimen,withthegoalofreducinginjection/handlingtimeinexperimentsand
tomaximizereliabilityandlaboratoryefficiency,aswellastominimizelong-term
costs.

27
CHAPTER2
METHODS
2.1 Overview
Thedeviceemploysasystemtoconduct48-72houroldzebrafishthrougha
liquidmedium(eggwater)usingasyringepump.Thecompletesystemconsistsof
threemainsubsystemsnamelythepump,opticaldetectionandentrapment
components.Thedeviceworksbyaspiratingindividualfishthroughatubeviaa
pressuregradientcreatedwithasyringepump.Eachcycleofthedeviceinvolvesthe
followingsteps:(1)loading,(2)sensing,(3)trapping,(4)injectionand(5)flushing.
Duringloading,asinglelarvaisextractedfromthereservoirandconductedthrough
atubepasttheopticaldetectionsubsystem.Atthesensingstage,theoptical
detectionsubsystemcomposedofaphotodiodeandalaser,sensestransmittedlight
fromthelaseranddiscernstheentryoflarvafromairbubblesanddebriswith
precision.Uponlarvarecognition,thespecimenisthenconductedtothe
entrapmentdock(step3)whereitwillbeimmobilizedforinjection(step4).The
finalstep(5)involvesconductingthelarvaoutoftheentrapmentdockand
subsequentlyoutofentiresystemforfurtheranalysis.Thesyringepumpispre-
programmedandmanipulatedwithLabView,thecontrolsoftware.Thisdevicewill
primarilyserveIAVresearcherswhointendtointroducevaccines,pathogensand
otherexperimentalmaterialsintomanyindividualzebrafishlarvae.

28
Figure3:Deviceschematicanddesign.Thedeviceoffersminimalinteractionandseamlessaspirationofzebrafishspecimen,ultimatelytrappingandimmobilizingitformicroinjectionofbiomaterials.

29
Thefollowingsectionsdescribeindetailthedesignandconstructionofthe
devicecomponents.
2.2 PumpSubunitComponents
Figure4:Animageoftheinputreservoir.TheassembledreservoirisshowninFigure4.1;Figure4.2andFigure4.3showthedesignrenderingsofthetopandbottomreservoircaps,respectively.

30
Thedevicepicturedaboveservesastheentrypointofthezebrafish
specimenintothedevice.Withamaximuminternalvolumeof60mL,thereservoiris
capableofaccommodatingabout50individual48-72hpflarvae.Whenfilledwith
eggwater(Figure4.1),theuniqueshapeanddesignallowsenoughroomforthefish
toswimandeventuallyberemovedbysuctionthroughtheoutlettubepositioned
alongthewallofthereservoir’sbottomcap.Duringtesting,itwasobservedthatthe
zebrafishlarvaehadthetendencytosinkdownward,towardthedeepestpartofthe
bottomcap.Thismadeitdifficultforspecimentoberandomlyremovedfromthe
reservoirthroughthetube,thusdramaticallyreducingthelarvaeoutputrate.To
remedytheproblem,abubblerwasintroduced.Anarrowholewasdrilledthrough
thedeepestpartofthebottomcaptoallowforasiliconetubetobethreaded
throughandconnectedtothebubbler.AnAquaCulturesingleoutletaquariumair
pumpwasusedastheairsourceFigure5.Itincludesacheckvalvethatprevents
thesiphoningofreservoirwaterintotheairpumpintheeventofapowerloss.
Figure5:AquaCulturesingleoutletaquariumairpump.Thepumpisoriginallyanairpumpdesignedfora5-to15-gallonaquarium.Itincludesacheckvalve(bluevalvepictured)thatcanhelppreventthesiphoningofreservoirwaterintotheairpumpintheeventofapowerloss.

31
PressureisprovidedtothesystemwithaNewEraNE-1000XBidirectional
SyringePump.Thepumpsuppliesnegativepressure(withdrawmode)forabrief
periodtocausespecimentobewithdrawnfromthereservoirandintothesystem
whereitwillbesensedbythelaserdiode/photodiodephotodetectionsystem.The
pumpthenswitchestopositivepressure(infusemode)toallowthespecimento
advancethroughthesystem.
Figure6:NewEraNE-1000Xbidirectionalprogrammablesyringepump.Withfullyprogrammableandstand-alonecapabilities,thepumpcanbeprogrammedforupto41pumpingphasesthatchangepumpingrates,setdispensingvolumes,insertpauses,controlandrespondtoexternalsignals.Ithasadispensingaccuracyof±1%.
2.3 OpticalDetectionSubunitComponents
Theopticaldetectionunithelpswiththedirectionoftravelofspecimen
withinthesystembysensingthepresenceofzebrafish,whichpromptsthesyringe
pumptoswitchbetweensupplyingpositiveandnegativepressure.Thetwomain
elementsusedforsensingare:a5mmtransparentcylinderheadphotodiode(PD)

32
1
1 22
(LLSO5-A,SenbaOpticalElectricalCo.,Ltd)anda4.5V650nmredlaserdiode
module(#01444878,LightInTheBoxCo.,Ltd).Duringnormaloperation,thered
laserbeamisfocusedontothephotodiodeandabase(unobstructed)voltage
readingisobservedandrecorded(voltagecomparatorcircuityieldsaHIGHsignal).
Themovementofspecimenpastthelightbeamreducesthevoltagereading,
resultinginthevoltagecomparatorcircuitproducingaLOWsignal,indicatingthe
presenceofalarvaasdepictedinthecartoon(Figure7.1).
Figure7:Animageoftheopticaldetectionunitcomponents.Figure7.1depictsacartoonthathighlightsazebrafishlarvamovingpasttheredlaserbeamtoindicatethepresenceofaspecimen;theblueselectionwithinFigure7.2showstheactualsetupofthedetectionsystemmountedona3Dprintedharness.2.4 EntrapmentDock
Acriticalcomponentoftheinjectionsystemistheentrapmentdock,adevice
thatimmobilizesthespecimenforinjection.ThedockwasdesignedusingSolidEdge
rapid-prototypingsoftwareand3Dprintedusingatransparentphotopolymerthat
PEEK Tubing
Photodiode
Laser

33
providesasuitablelevelofsurfacesmoothness.Thepolymer(VeroClear-RGD810)
isarigid,nearlycolorlessmaterialwithdimensionalstabilityforgeneralpurpose,
fine-detailmodelbuildingandvisualsimulationoftransparentthermoplastics.The
entrapmentdockdimensionsarerepresentedinFigure8.
Figure8:Imageoftheentrapmentdockwithhighlightedchannelsanddimensionsinmillimeters.
Thechannelswithinthedockaredesignedtoaccommodatetheshapeofa2-
dayoldzebrafishlarva.Beginningfromoneendofthedock,thechannelbeginswith
a1mmdiameteropeningtoaccommodatethecapillarytubefromtheautomated
deliverysystem.Itthennarrowstoamicro-constrictionthatissmallenoughto
allowsomeexcesswatertoflowpastthespecimentotheotherendofthedock
whilesimultaneouslytrappingandimmobilizingthespecimenforinjection.An

34
openingperpendiculartothetrappedspecimenservesasanentryfortheinjection
needle.
Despitetheconvenienceandpracticalityof3Dprinting,theabilityto
fabricatesmall,intricatedesignssuchasmicrochannelscansometimesbelimited.
Althoughprintersandtechniquesexistformicrofabrication,thecostofsecuringa
high-resolutionprintingunitishigh,withpricesstartingat20,000USD.Less
expensiveunitscanperformsimilarfunctionstothemoreexpensivemachines,but
withlessprecision.Thelabinwhichtheentrapmentdockwasprinted(Innovative
MediaResearch&CommercializationCenter,UniversityofMaine)wasequipped
withanintermediateresolution3Dprinter(StratasysObjet30,SUP705support
material),andsoseveralchallengesinfluencedthefinaldesignoftheport.Most
notableamongstthesewastheproblemoflodgedsupportmaterialwithinthe
microchannels.Most3DprintersemploytheAdditiveManufacturing(AM)design
techniquewheresuccessivelayersofmaterialareformedundercomputercontrol
tocreateanobject;itisanalogoustobuildingabrickwalllayerbylayer.Ifa
particulardesignissupposedtohaveanenclosedopeningorchannel,support
materialisdepositedintothecavitiestoholdupthestructureastherestofthe
objectisbeingbuilt.Asaresultofthisprocess,alloftheentrapmentdockswere
filledwithsupportmaterialandpost-printingprocedurestoremoveitprovedfutile.
Someoftheproceduresusedwere:applyingmildheat,usingbiodegradable
solventsandmanualexcavationofthematerialusingathinsyringeneedle.These
problemsspawnedtheideaofbisectingtheoriginaldesignoftheporthorizontally
andprintingbothpiecesindependently,asdisplayedinFigure9.2.Thisexposesthe

35
channelsandallowsforeasyremovalofthesupportmaterialwithoutdamagingthe
integrityofthechannels.Asupportingharnesswaslaterdesignedand3Dprintedto
rejoinandclamptogetherthepiecesofentrapmentdockFigure9.3.

36
1 2
3 4
5
Figure9:ImagesoftheentrapmentdockanditssupportingcomponentsdesignedusingSolidEdgeCADsoftware.The3Dprintedsolidisatransparentmaterial(VeroClear-RGD810)thatprovidesdimensionalstabilityandarigidfinish.Figure9.1isanimageoftheassembledentrapmentdock;Figure9.2depictsthetwosplit-piecesoftheentrapmentdock;Figure9.3showstherenderingofthesupportingharnessthatclampsandkeepsthesplit-piecesinplace;Figure9.4showstherendingofthecompletelyassembleddevicewiththeentrapmentdockhighlightedingreen;Figure9.5isanimageofthecompletelyassembleddevice.

37
2.4.1 Biocompatibility
3Dprintingandphotopolymercompositionhaveimprovedtremendously
withinthepastfewyears,andproblemswithbiocompatibilityhavemostlybeen
overcomebypost-processingthe3Dprinteddeviceandoptimizingthechemistryof
theresinsusedtocreatemicro-andmilli-fluidicplatforms.Thepolymerusedto
printtheentrapmentdock(VeroClear-RGD810)isdimensionallystableandis
approvedformedicalapplications93.Infuturepursuitstoperfecttheentrapment
dockandtoaccommodatedifferentorganisms,anadvancedpolymer(MED610,
madebyStratasys)couldbeused.Thisplastichasawiderbiocompatibilityrange
andmightserveasabetteralternative94.
Macdonaldet.alattheUniversityofGlasgowassessedthebiocompatibilityof
fourcommerciallyavailable3Dprintingpolymers(VisiJetCrystalEX200,Watershed
11122XC,FototecSLA7150ClearandABSplusP-430)withzebrafishandobserved
thekeydevelopmentalmarkersinthedevelopingembryos.Theirresultsshowed
thatthefourphotopolymerswereverytoxictotheembryos,resultinginfatalityin
mostcases95.Theseresultsre-emphasizetheimportanceofusingbiocompatible
materialstoachievereliableresults,especiallywhenconductingexperimentsin
whichmorphologicalchangesareanalyzed.Itisbecomingwidelyacknowledged
thatmoredetailedbiocompatibilitytestingmustbeprovidedbymanufacturers
before3Dprintedobjectsareusedindifferentbiosystems96,97.

38
2.5 SpecificationsandParameters
1. The48-72hpfzebrafishlarvaecanbeaspirateduptoamaximumflowrate
of330mm3s-1,giventubingdimensionsof0.8mminnerdiameterx2.4mm
outerdiameter.
2. Themaximumspeedthelarvaecanattaininthistubingwithoutdeformation
is656mms-1
3. Inthissystem,Re=528at20◦C.
4. Themaximumaspirationrateatthedeliverytube/capillaryjunctionshould
bebelow83μLs-1toavoiddamagingthelarvae.
5. Theinnerdiameteroftheinputcapillaryis800μm
2.6 Electronics
Toautomatemovementofthelarvaeinthepresentsystem,electronic
sensors(e.g.photodiode)andactuators(e.g.syringepump),wereusedtocontrol
fishmovementthroughaLabVIEWinterface.ThemyDAQ(NationalInstruments,
Austin,TX)wasusedtodetecteventsandtocontroldevices,suchastheSC5
solenoidcontroller(RWAutomations,Figure10)usedtocontrolthesolenoidvalves.

39
Figure10:ElectronicComponents.RWAutomationSC5SolenoidControllerboardequippedwithpower,communication,andstatusLEDs(Left);NationalInstrumentsmyDAQ(Right)
ThemyDAQfromNationalInstruments(NI)isaDataAcquisition(DAQ)unit
thatinterfaceswiththegraphicalprogrammingsoftwarepackage,LabView,and
allowsforsimpleexternalcontrolofconnecteddevices.NImyDAQisasimpleand
intuitiveDAQdevicewithanaloginputs,analogoutputs,digitalinputsanddigital
outputs.ThedigitaloutputchannelsareusedtocontroltheTransistor-Transistor
Logiccommands(TTL)thattriggerthesolenoidpinchvalves.ThemyDAQwasalso
beusedasapowersupply.
TheRWAutomationSC5solenoidcontrollerpermittedprogrammable
controlofthesolenoidpinchvalvedepictedinFigure11.Theboardcontrolsfive
independentvalvesbyutilizingapeak-and-holdalgorithm.Thisallowstheuserto

40
energizeeachsolenoidvalveatitsfullratedvoltageforfastactuationandstrong
pull.TheSC5thenautomaticallyreducestheaveragevoltageappliedtothesolenoid
toconservepowerandpreventoverheatingofthesolenoidintheopen/onposition.
Figure11:Cole-Parmertwo-waynormallyclosedsolenoidpinchvalve.12VDC,1/16"IDx1/8"ODtubing.Thesolenoidcontrolledpinchvalveclampswithaforceof15PSIwitha25msecresponsetime.
Whentriggered,thesolenoidcontrollerapplies12Vtothesolenoidvalvesfor
0.5sec,thenreducesthevoltagetoaholdinglevelof~4.5V.Thisminimizes
overheatingofthesolenoid.TheSC5controllerisunderTTLcontrolandopensor
closesthesolenoidpinchvalveswithhighorlowvoltagesignalsfromtheDIO

41
channelsofthemyDAQcontroller.LowtohighvoltageTTLinputtransitionstrigger
thesolenoidvalveopening,whilehightolowTTLvoltagetransitionsturnoffthe
solenoidvoltagetodisablethesolenoidandclosethevalve(NCstate).TTLinputsof
thesolenoidcontrollerboardareinterfacedwithDIOports0-4onthemyDAQand
controlledbyaLabViewVIwithBooleancontrols.
Toenhancetheclarityofdataandtopermithigherdataacquisitionrates
fromtheopticaldetectionsystem,aprecisionhigh-speedvoltagecomparatorwas
used(TexasInstrumentsLM339N)toconvertthephotodiodevoltageoutputtoa
TTLsignal.Thevoltagecomparatorfunctionsbycomparingthedifferentialvoltage
betweenthepositivepin(sporadicvoltagefromPD)andnegativepin(stable
predeterminedvoltagefrommyDAQ),thengeneratingaTTLsignalbasedonthe
inputdifferentialpolarityFigure12.1.Thiscircuitrendersthesystemlesssensitive
topotentialfalsereadingsfromthePD.

42
Figure12:Imageofthefunctionalcircuitdiagram.Voltagecomparator(left);wiringofcomparatoronabreadboard(Right)
TheLM339NseriesICconsistsoffourindependentprecisionvoltage
comparatorswithanoffsetvoltagespecificationaslowas2mVmaximumforall
fourcomparators.Thecomparatorisdevisedtooperatefromasinglepowersupply
overawiderangeofvoltagesandtodirectlyinterfacewithTTL(Transistor-
TransistorLogic)andCMOS(ComplementaryMetal–Oxide–Semiconductor).
2.6.1 ControlMaps/SoftwareProgramming
Mostofthesystemismanagedmanuallythroughavirtualinstrument
dashboarddesignedinLabView,Figure13.Theplatformallowsautomationofthe
solenoidvalvesandthesyringepumptoaspiratezebrafishlarvaeintothe
entrapmentdock.
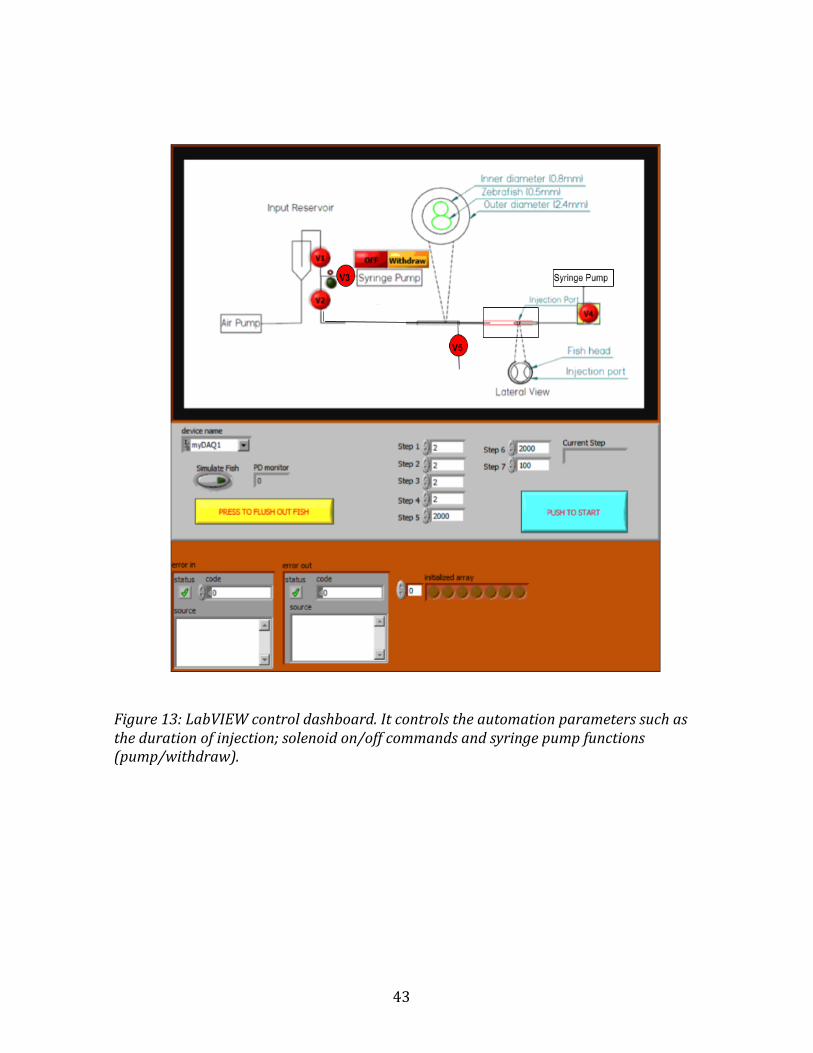
43
Figure13:LabVIEWcontroldashboard.Itcontrolstheautomationparameterssuchasthedurationofinjection;solenoidon/offcommandsandsyringepumpfunctions(pump/withdraw).

44
LabViewTestSetupRoutine
ThefollowingisthedataflowdiagramfortheLabViewroutine,which
controlsthecoordinatedfunctionsofthesyringepump,thepinchvalvesandthe
opticaldetectionsystem.
Figure14:Valve/syringepumpcontrolmap
Allthesolenoidvalvesarenormallyclosed.Acyclebeginswiththeopening
ofvalve1andvalve3.Thesyringepumpstartswithdrawingatagivenflowrate
untilalarvaispickedupfromreservoir1,conductedthroughthetubeanddetected
bytheopticaldetectionsystem.Thesyringepumpimmediatelystopswithdrawing
andvalve1andvalve3close.Next,valve2andvalve3open,followedbythe
syringepump,ininfusemode,aspiratingthelarvaatthesetflowratetothe
entrapmentdock.Valve2closestopreventbackflow.Injectiontakesplace.Valve4

45
andvalve5openafterinjection.Syringepump2,ininfusemode,flushesthelarva
outoftheentiresystem.
Thedetailedsetupandprogramcommandsarehighlightedintheappendix.

46
CHAPTER3
EXPERIMENTALRESULTSANDDISCUSSION
3.1 InputReservoirEfficacyTest
Toevaluatethespecimenoutputrateofthereservoir,asystemtestwas
conducted.
3.1.1 Procedure
Thereservoirwasfilledtocapacity(60mL)witheggwater(60µg/mL)at
roomtemperature(21°C),Figure4.1.Thebubblerwasthenturnedontoprevent
settling,therebyincreasingthelikelihoodoflarvauptakeintothesystem.50
individualzebrafishlarvaewerethenintroducedintothereservoir.Thezebrafish
wereanesthetizedwitha200µg/mLsolutionoftricainebeforeuse.Withthelarvae
floatingfreelywithinthereservoir,theaspirationtubewasloweredintothe
chamberandthesyringepumpwassettowithdrawmodeatthefollowingflow
ratesfor60seconds:10mL/min,8mL/min,6mL/minand4mL/min.Thenumberof
larvaeaspiratedfromthereservoirwasthenrecordedandtheentireprocedurewas
repeated10times,consecutively.
3.1.2 Results
Fortheentrapmentsystemtobemoreefficientthanthetraditionalinjection
method,thereservoirshouldexpelthelarvaefromthesystematabaserateof3fish
perminute.Thetestofthereservoiratthehighestflowrate(10mL/min)generated
anaverageoutputofabout6larvaeperminuteFigure15.Thistranslatesto1larva

47
outputevery10seconds,whichisarelativelyshorterdurationthantheremaining
timeittakesalarvatocompletetherestofthecycle.Thisimpliesthatthereservoir
musthaveanoutputratehighenoughtosupporttheentiresystem.Inspiteofthe
highoutputrate,onlyonelarvacanbeprocessedbythesystemforanygivencycle.
Thehighoutputrateofthereservoirsuggeststhatithasthecapacitytofeed
multipleentrapmentsystemssimultaneously.

48
Figure15:Abargraphdepictingthenumberofzebrafishlarvaecollectedafter60secatdifferentflowrates.3.2 OpticalDetectionUnitEfficacyTest
Toevaluatethesensitivityoftheopticaldetectionunit,asystemtestwas
conductedroutinely.
3.2.1 Procedure
First,the650nmlasermodulewaspoweredon,withmajorityofthelight
beamcenteredonthephotodiode(PD).AtthispointtheLabViewsoftware,
registeredabasalhighTTLsignal(1,TRUE).Tenindividualzebrafishlarvae
(~48hpf)werethenfedthroughthetube,pastthelaser/PD,at10mL/min.The

49
decreaseinlightdetectedbythePDresultedinalowTTLsignal(0,FALSE)fromthe
comparatorasthelarvapassedbetweenthelaserandthephotodiodeandwas
monitoredbythecontrolsoftware.
3.2.2 Results
Theresultsfromsensortestsprovedthatthelightdetectionsystem
effectivelydetectedlarvae.Thewaveformgraph,Figure16,displays10individual
peaksthatcorrespondtothe10-zebrafishspecimensusedinthistest.Theinitial
voltagereading(withoutacomparatorcircuit)from7-14secondsindicatesthe
observedvoltagebeforewaterorzebrafishlarvaewereintroducedintothetube.
Thebackgroundnoiseapparentinseveralpositionsonthechart(ex.14-25
seconds)canbeattributedtoairbubbles.Thecontrolsoftwaretracestherecorded
voltagereadingsthatregisteredbelowthebasalreading(tube+water+specimen),
thusallowingustodiscernbackgroundnoisefromusefuldata.

50
1 234 1095 86
7
Figure16:Awaveformchartofthephotodiodevoltagespikeswithrespecttotime.ThebasereadingfromthePDis1V.Therestofthelabeledpeaksregisteringsub-basalvoltagesindicatethepresenceofazebrafishspecimen.Thecomparatorcircuitwasnotusedtoconditiontheserawvoltagesignals.
3.3 EntrapmentDockTests
Theentrapmentdockisthemainpartofthesystemwiththecorefunctionof
immobilizing48-72hpfzebrafishtoallowtailinjection.Aseriesoftesttrialswere
conductedtoassessitsabilitytoentrapandimmobilize48-72hpfzebrafish.
3.3.1 Procedure
Totestitsabilitytoaccomplishtrapping,individual72hpfzebrafishwere
introducedintothedockviatheinletchannelandsubsequentlyflushedout.For
everytrialthatwasconducted(n=100),thelarvawassuccessfullyimmobilized
regardlessoforientation(head/tailentry)asillustratedinFigure17.

51
Figure17:Imageoftheentrapmentdockillustratingtheimmobilizedzebrafishlarvaeinthetwopossibleorientations.Head-onarrival(leftimage)andtailarrival(rightimage)3.4 AverageDurationofScreeningSteps(1Cycle)
Theoveralldurationforonecyclethroughthesystemwasdeterminedinthistest.
3.4.1 Procedure
Bytimingeachautomatedstageoftheprocesses(loading,trapping,injection
andflushing)withinonecycle,theaveragecycletimewascalculated.Sincetheflow
rateisvariedatcertainstageswithinagivencycle,timingeachstepprovidesamore

52
accurateprojectionoftheoveralldurationofonecycle.
Table1:Averagedurationofscreeningstepsfromthereservoirtothelightdetectoratdifferentflowrates.(0.08cminnerdiameterand88cmlength)
Table2:Calculateddurationofscreeningstepsfromthephoto-detectortotheentrapmentdockatdifferentflowrates.(0.08cminnerdiameterand80cmlength)
Table3:AveragetimerequiredforinjectionStep 5mL/min 4mL/min 3mL/min 2mL/minEntrapmentDock→Exit 4 5 6 9Table4:Calculateddurationofscreeningstepsfromtheentrapmentdocktotheexit(oftheentiresystem)atdifferentflowrates.(0.08cminnerdiameterand62cmlength)
Step FlowRate(mL/min) Time(sec)
Reservoir→PD 10mL/min 9±6.5
Reservoir→PD 8.0mL/min 10±10.4
Reservoir→PD 6.0mL/min 13±19.5
Reservoir→PD 4.0mL/min 16±15.3
Step FlowRate(mL/min) Time(sec)
PD→EntrapmentDock 2mL/min 12
PD→EntrapmentDock 1.75mL/min 14
PD→EntrapmentDock 1.50mL/min 16
PD→EntrapmentDock 1.25mL/min 19
Step Time(sec)Injectionprocedure Userdependent:10±3

53
3.5 QuantitativeAssessmentofAnimalHealth
Theconditionofeachindividualzebrafishwasassessedafteritpassed
throughthesystem.Thisassessmentwasbasedonbothfunctionaland
morphologicalcriteria.Atallflowrates(2mL/min,1.75mL/min,1.5mL/min,
1.2mL/min),heartbeat,touch-response,structuralintegrityoftheyolksacand
melanocytes,wereevaluatedandcomparedtoacontrolsample.Thelarvaeinthe
controlsetup(sameage)wereaspiratedinanunobstructedtubespanningthe
lengthoftravelencounteredbylarvaeintheexperimentalsetup.
3.5.1 Procedure
Thezebrafishfromthetestsdescribedinsection3.4werecollectedand
viewedunderamicroscopetoevaluatethefollowingparameters:survival
(heartbeat)andmorphology(structuralintegrityoftheyolksacandmelanocytes).
ThefollowingdatawererecordedinFigure18.
3.5.2 Results
Atalloftheflowratesused,100%ofthelarvaesurvived(n=100)foratleast.
Tearingoftheyolksacwasneverobserved.Atthehighestinitialflowrateof
2mL/min,48%ofthelarvaeexhibitedmorphologicalabnormalities,specifically
slightdistortionofmelanocytesalongthetailandoftheyolkextension,Figure18.
Withtheslightlyslowerinitialaspirationratesof1.75mL/min,allhealthcriteria
matchedthoseofthecontrols.

54
Figure18:Comparisonofmorphologicalabnormalitiesofexperimentalzebrafish,atdifferentflowrates,withacontrolzebrafish.Theredarrowsshowdistortionsofthemelanocytesandthehighlightedregionshowsdistortionsoftheeyeandtheyolk.

55
Figure19:Quantitativeassessmentofzebrafishhealthatdifferentflowrates(n=100)

56
CHAPTER4
SUMMARY,CONCLUSIONSANDFUTUREWORK
4.1 Summary
Adevicetoaccelerateexperimentsinvolvingmicroinjectionoflargenumbers
oflarvaewasdesignedandfabricatedusinginexpensiverapidprototyping
techniques.Chapter1introducestheinfluenzaAvirusanddescribesimportant
aspectsofthevirus,includingitsgenetics,distribution,transmissionandclinical
symptomsofthediseasethatitcauses.Thischapteralsoplacesemphasisoncurrent
IAVresearchadvancementsandreviewstheuseofanimalmodelsforinfectious
diseaseresearch.Chapter2describesthedevicecomponentsseparatelyand
reviewsprototypingmethods,modesofdevicefunctionandsupportingcontrol
software.Thedevicespecificationsandparametersarealsohighlightedinthis
chapter.Chapter3presentsefficacytestsofthemaincomponentsofthesystem.
4.2 Conclusions
Afullyfunctionalzebrafishimmobilizationdevicewasdesigned,fabricated,
andtested.48-72hpfzebrafishlarvaeweresuccessfullyimmobilizedwithina3D
printeddock.Theexperimentaltestsetupallowedmanipulationofthevolumetric
flowratewithinthetubes,pressurefromtheaircompressorusedtocreateair
bubblesinreservoir1,voltagewithintheopticaldetectionsystemandtheoverall
deviceoutputfrequency.Theloadingstagewasimprovedwiththeadditionofthe
bubblertoreservoir1.Thisplayedamajorroleindistributinglarvaeevenlywithin
thechamberduringoperation.

57
Automationofthesystemenhancedtestreproducibilitybyreducingthe
chancesforhumanerror.Theopticaldetectionsystemprovedeffective(see
waveformchart,Figure16.Individuallarvaeweresuccessfullyidentifiedbythe
detectionsystemwith100%precision(nofalsepositives).Thecontrolsoftware
tracedtherecordedvoltagereadingsthatregisteredbelowthebasalreading;
moreover,thecomparatorincreasedthedataacquisitionratetwo-fold,from62Hz
to124Hz.
Oneofthemostchallengingaspectsoftheprojectwasthefabricationofthe
immobilizationdock.The3Dprinterthatwasusedforthisprojectdeposited
supportmaterialwithinthechannelsandnomethodusedtoremovethematerial
wassuccessful.Bydevelopingthesplitdesigntechnique,Ieliminatedthisproblem
byexposingthechannelsforsimplifiedpost-printprocessing.Thetestsofthe
entrapmentdockweresuccessful,i.e.afterrunning100cyclesthroughthedockat
variousflowrates:48%oflarvae(n=25)wereimmobilizedwithoutany
deformationat2mL/min,76%oflarvae(n=25)wereimmobilizedwithoutany
deformationat1.75mL/min,100%oflarvae(n=25)wereimmobilizedwithoutany
deformationat1.50mL/min,100%oflarvae(n=25)wereimmobilizedwithoutany
deformationat1.25mL/min.
4.3 FutureWork
Atthisstageofdevicedevelopment,adjustingtheflowratewithinthesystem
duringcertainpartsofagivencyclecanalterthefrequencyofthedevice’soutput.In
spiteoftheautomationincorporatedintothedevice,theusermustmanuallytrigger

58
thecontrolprogramtoexecutecertaincommandssuchaswhentoinjectthe
specimen.Byaddingaminimalnumberofstepstotheexistingcontrolprogram,an
automaticinjectorequippedwithatranslationstagecouldbeincorporatedintothe
designtoreducetheneedformanualinput.Furthermore,arecoverysystemcould
easilyintegratedtowardtheendofthedevicetoreceivetheinjectedlarvaeafter
ejectionfromthesystem.Ultimately,thedevicewillprovideaseamlessuser
interfacethatrequiresminimaluserintervention.Bydepositingaknownnumberof
zebrafishlarvaeintotheinputreservoir,andinputtingthedosageofthecompound
orchemicalunderstudy,thedevicewillbeabletoinjectanddispenselarvaefor
analysis.
Microsystemsdesignedforcell-basedstudiesorapplicationssuchasthisone
requirefluidhandling.Flowwithinthesesystemsinevitablygeneratesfluidshear
stressthatmayadverselyaffectthehealthoftheorganismsunderstudy.Simple
assaysofspecimenviabilityandmorphologywereusedinthisprojecttodetectany
grossdisturbancestolarvalanatomyandphysiology.However,itwillbebeneficial
tospecificallyevaluatephysiologicalimplicationsoffluidshearstresswithinthe
tubesandtheentrapmentdock.Varmaetal.atMassachusettsInstituteof
Technologyexploredausefultechniquethatcanassistuswithunderstandingthe
fluiddynamicswithinthisdevice.Itinvolvesusingageneticallyencodedcellsensor
thatfluorescesinaquantitativefashionuponencounteringfluidshearstress
(FSS)98.Varmaetal.choseawidelyusedcellline(NIH3T3s)andcreateda
transcriptionalcell-sensorthatfluoresceswhentranscriptionofarelevantFSS-
inducedproteinisinitiated.Thesensorpathwayspecificityandfunctionalitywere

59
verifiedbynotinginducedfluorescenceinresponsetochemicalinductionoftheFSS
pathway,seenboththroughmicroscopyandflowcytometry.Inaddition,these
novelcellsensorscanbeinducedwitharangeofFSSintensitiesanddurations,with
alimitofdetectionof2dynes/cm2whenappliedfor30minutes,makingitmore
versatileinsimulatingseveralmicrofluidicflowconditions.Undertakingthe
improvementsdiscussedabovewillenableforamoreseamlessoperationofthis
deviceandpossiblypavethewayforsubsequentengineeringsystemswithwide
applicationsinthemicrosystemscommunity.

60
BIBLIOGRAPHY
1. YoderJA,NielsenME,AmemiyaCT,LitmanGW.Zebrafishasanimmunologicalmodelsystem.MicrobesInfect.2002;4(14):1469-1478.doi:10.1016/S1286-4579(02)00029-1.
2. GaborKa,GoodyMF,MowelWK,etal.InfluenzaAvirusinfectioninzebrafishrecapitulatesmammalianinfectionandsensitivitytoanti-influenzadrugtreatment.DisModelMech.2014;7(11):1227-1237.doi:10.1242/dmm.014746.
3. VarkiA,GagneuxP.Multifariousrolesofsialicacidsinimmunity.AnnNYAcadSci.2012;1253:16-36.doi:10.1111/j.1749-6632.2012.06517.x.
4. KimmelCB,BallardWW,KimmelSR,UllmannB,SchillingTF.Stagesofembryonicdevelopmentofthezebrafish.DevDyn.1995;203(3):253-310.doi:10.1002/aja.1002030302.
5. WorobeyM,HanG-Z,RambautA.Asynchronizedglobalsweepoftheinternalgenesofmodernavianinfluenzavirus.Nature.2014;508(7495):254-257.doi:10.1038/nature13016.
6. CouchRB.Orthomyxoviruses.1996.http://www.ncbi.nlm.nih.gov/books/NBK8611/.AccessedMay18,2016.
7. HILLEMANM.Realitiesandenigmasofhumanviralinfluenza:pathogenesis,epidemiologyandcontrol.Vaccine.2002;20(25-26):3068-3087.doi:10.1016/S0264-410X(02)00254-2.
8. MosnierA,CainiS,DaviaudI,etal.ClinicalCharacteristicsAreSimilaracrossTypeAandBInfluenzaVirusInfections.PLoSOne.2015;10(9):e0136186.doi:10.1371/journal.pone.0136186.
9. LemaitreM,CarratF.Comparativeagedistributionofinfluenzamorbidityandmortalityduringseasonalinfluenzaepidemicsandthe2009H1N1pandemic.BMCInfectDis.2010;10(1):162.doi:10.1186/1471-2334-10-162.
10. KeyFactsaboutInfluenza(Flu)&FluVaccine|SeasonalInfluenza(Flu)|CDC.http://www.cdc.gov/flu/keyfacts.htm.AccessedMarch14,2016.

61
11. ThompsonWW,ShayDK,WeintraubE,etal.Influenza-associatedhospitalizationsintheUnitedStates.JAMA.2004;292(11):1333-1340.doi:10.1001/jama.292.11.1333.
12. FouchierRAM,SchneebergerPM,RozendaalFW,etal.AvianinfluenzaAvirus(H7N7)associatedwithhumanconjunctivitisandafatalcaseofacuterespiratorydistresssyndrome.ProcNatlAcadSciUSA.2004;101(5):1356-1361.doi:10.1073/pnas.0308352100.
13. HayAJ,GregoryV,DouglasAR,LinYP.Theevolutionofhumaninfluenzaviruses.PhilosTransRSocLondBBiolSci.2001;356(1416):1861-1870.doi:10.1098/rstb.2001.0999.
14. PotterCW.Ahistoryofinfluenza.JApplMicrobiol.2001;91(4):572-579.doi:10.1046/j.1365-2672.2001.01492.x.
15. CDCNovelH1N1Flu|The2009H1N1Pandemic:SummaryHighlights,April2009-April2010.http://www.cdc.gov/h1n1flu/cdcresponse.htm.AccessedMay20,2016.
16. Laboratory-ConfirmedInfluenzaHospitalizations.http://gis.cdc.gov/grasp/fluview/FluHospChars.html.AccessedMay19,2016.
17. NoWV.Estimatesofdeathsassociatedwithseasonalinfluenza---UnitedStates,1976-2007.MMWRMorbMortalWklyRep.2010;59(33):1057-1062.doi:mm5933a1[pii].
18. PreventionandControlofInfluenzawithVaccines.http://www.cdc.gov/mmwr/preview/mmwrhtml/rr59e0729a1.htm.AccessedMay19,2016.
19. FluTransmission.https://www.niaid.nih.gov/topics/flu/understandingflu/Pages/transmission.aspx.AccessedMay20,2016.
20. ThomasY,VogelG,WunderliW,etal.Survivalofinfluenzavirusonbanknotes.ApplEnvironMicrobiol.2008;74(10):3002-3007.doi:10.1128/AEM.00076-08.

62
21. ThomasY,Boquete-SuterP,KochD,PittetD,KaiserL.Survivalofinfluenzavirusonhumanfingers.ClinMicrobiolInfect.2014;20(1).doi:10.1111/1469-0691.12324.
22. P.SivaramakrishnaRachakonda:STUDIESONCONFORMATIONALSTABILITYOFTHEECTODOMAINOFINFLUENZAVIRUSHEMAGGLUTININ.http://edoc.hu-berlin.de/dissertationen/rachakonda-p-sivaramakrishna-2005-02-15/HTML/chapter1.html.AccessedMay20,2016.
23. PneumoniaSymptoms&Treatment|ClevelandClinic.http://my.clevelandclinic.org/health/diseases_conditions/hic_Pneumonia.AccessedMay20,2016.
24. MedinaRa,García-SastreA.InfluenzaAviruses:newresearchdevelopments.NatRevMicrobiol.2011;9(8):590-603.doi:10.1038/nrmicro2613.
25. NicholsonKG,WoodJM,ZambonM.Influenza.Lancet(London,England).2003;362(9397):1733-1745.doi:10.1016/S0140-6736(03)14854-4.
26. TsaiKN,ChenGW.Influenzagenomediversityandevolution.MicrobesInfect.2011;13(5):479-488.doi:10.1016/j.micinf.2011.01.013.
27. KanekiyoM,WeiC-J,YassineHM,etal.Self-assemblinginfluenzananoparticlevaccineselicitbroadlyneutralizingH1N1antibodies.Nature.2013;499(7456):102-106.doi:10.1038/nature12202.
28. MelcherL.Recommendationsforinfluenzaandpneumococcalvaccinationsinpeoplereceivingchemotherapy.ClinOncol.2005;17(1):12-15.doi:10.1016/j.clon.2004.07.010.
29. TaubenbergerJK,MorensDM.Thepathologyofinfluenzavirusinfections.AnnuRevPathol.2008;3:499-522.doi:10.1146/annurev.pathmechdis.3.121806.154316.
30. Pardo-MartinC,ChangT-Y,KooBK,GillelandCL,WassermanSC,YanikMF.High-throughputinvivovertebratescreening.NatMethods.2010;7(8):634-636.doi:10.1038/nmeth.1481.

63
31. Flu(Influenza)BasicResearch.https://www.niaid.nih.gov/topics/Flu/Research/basic/Pages/default.aspx.AccessedMay23,2016.
32. GenomeSequencingCenters.http://www.niaid.nih.gov/labsandresources/resources/dmid/gsc/influenza/Pages/default.aspx.AccessedMay23,2016.
33. DavidsonJM.Animalmodelsforwoundrepair.ArchDermatolRes.1998;290(14):S1-S11.doi:10.1007/PL00007448.
34. LeeKY,DeMayoFJ.Animalmodelsofimplantation.Reproduction.2004;128(6):679-695.doi:10.1530/rep.1.00340.
35. BrownL,PanchalSK.Rodentmodelsformetabolicsyndromeresearch.JBiomedBiotechnol.2011;2011.doi:10.1155/2011/351982.
36. JohnsonTV.,TomarevSI.Rodentmodelsofglaucoma.BrainResBull.2010;81(2-3):349-358.doi:10.1016/j.brainresbull.2009.04.004.
37. TheUseofAnimalModelsinStudyingGeneticDisease.http://www.nature.com/scitable/topicpage/the-use-of-animal-models-in-studying-855.AccessedMay25,2016.
38. BouvierNM,LowenAC.AnimalModelsforInfluenzaVirusPathogenesisandTransmission.Viruses.2010;2(8):1530-1563.doi:10.3390/v20801530.
39. TredeNS,LangenauDM,TraverD,LookAT,ZonLI.Theuseofzebrafishtounderstandimmunity.Immunity.2004;20(4):367-379.doi:10.1016/S1074-7613(04)00084-6.
40. VanDerSarAM,AppelmelkBJ,Vandenbroucke-GraulsCMJE,BitterW.Astarwithstripes:Zebrafishasaninfectionmodel.TrendsMicrobiol.2004;12(10):451-457.doi:10.1016/j.tim.2004.08.001.
41. SullivanC,KimCH.Zebrafishasamodelforinfectiousdiseaseandimmunefunction.FishShellfishImmunol.2008;25(4):341-350.doi:10.1016/j.fsi.2008.05.005.

64
42. RobertsonGN,McGeeCAS,DumbartonTC,CrollRP,SmithFM.Developmentoftheswimbladderanditsinnervationinthezebrafish,Daniorerio.JMorphol.2007;268(11):967-985.doi:10.1002/jmor.10558.
43. GratacapRL,RawlsJF,WheelerRT.MucosalcandidiasiselicitsNF-κBactivation,proinflammatorygeneexpressionandlocalizedneutrophiliainzebrafish.DisModelMech.2013;6(5):1260-1270.doi:10.1242/dmm.012039.
44. ZhengW,WangZ,CollinsJE,AndrewsRM,StempleD,GongZ.Comparativetranscriptomeanalysesindicatemolecularhomologyofzebrafishswimbladderandmammalianlung.PLoSOne.2011;6(8):e24019.doi:10.1371/journal.pone.0024019.
45. WinataCL,KorzhS,KondrychynI,ZhengW,KorzhV,GongZ.Developmentofzebrafishswimbladder:TherequirementofHedgehogsignalinginspecificationandorganizationofthethreetissuelayers.DevBiol.2009;331(2):222-236.doi:10.1016/j.ydbio.2009.04.035.
46. LamS.,ChuaH.,GongZ,LamT.,SinY.Developmentandmaturationoftheimmunesysteminzebrafish,Daniorerio:ageneexpressionprofiling,insituhybridizationandimmunologicalstudy.DevCompImmunol.2004;28(1):9-28.doi:10.1016/S0145-305X(03)00103-4.
47. MeekerND,TredeNS.Immunologyandzebrafish:Spawningnewmodelsofhumandisease.DevCompImmunol.2008;32(7):745-757.doi:10.1016/j.dci.2007.11.011.
48. TredeNS,LangenauDM,TraverD,LookAT,ZonLI.Theuseofzebrafishtounderstandimmunity.Immunity.2004;20(4):367-379.http://www.ncbi.nlm.nih.gov/pubmed/15084267.AccessedDecember2,2016.
49. DobsonJT,SeibertJ,TehEM,etal.CarboxypeptidaseA5identifiesanovelmastcelllineageinthezebrafishprovidingnewinsightintomastcellfatedetermination.Blood.2008;112(7).
50. KantherM,RawlsJF.Host-microbeinteractionsinthedevelopingzebrafish.CurrOpinImmunol.2010;22(1):10-19.doi:10.1016/j.coi.2010.01.006.

65
51. EncinasP,Rodriguez-MillaMA,NovoaB,EstepaA,FiguerasA,CollJ.Zebrafishfinimmuneresponsesduringhighmortalityinfectionswithviralhaemorrhagicsepticemiarhabdovirus.Aproteomicandtranscriptomicapproach.BMCGenomics.2010;11:518.doi:10.1186/1471-2164-11-518.
52. BurgosJS,Ripoll-GomezJ,AlfaroJM,SastreI,ValdiviesoF.Zebrafishasanewmodelforherpessimplexvirustype1infection.Zebrafish.2008;5(4):323-333.doi:10.1089/zeb.2008.0552.
53. ManicassamyB,ManicassamyS,Belicha-VillanuevaA,PisanelliG,PulendranB,García-SastreA.AnalysisofinvivodynamicsofinfluenzavirusinfectioninmiceusingaGFPreportervirus.ProcNatlAcadSciUSA.2010;107(25):11531-11536.doi:10.1073/pnas.0914994107.
54. Hemagglutinin.http://www.sinobiological.com/Influenza-Hemagglutinin-Function-a-147.html.
55. ItoT,CouceiroJN,KelmS,etal.MolecularbasisforthegenerationinpigsofinfluenzaAviruseswithpandemicpotential.JVirol.1998;72(9):7367-7373.http://www.ncbi.nlm.nih.gov/pubmed/9696833.AccessedNovember14,2016.
56. LeungHSY,LiOTW,ChanRWY,ChanMCW,NichollsJM,PoonLLM.EntryofinfluenzaAViruswithaα2,6-linkedsialicacidbindingpreferencerequireshostfibronectin.JVirol.2012;86(19):10704-10713.doi:10.1128/JVI.01166-12.
57. LakadamyaliM,RustMJ,ZhuangX.Endocytosisofinfluenzaviruses.MicrobesInfect.2004;6(10):929-936.doi:10.1016/j.micinf.2004.05.002.
58. ChuVC,WhittakerGR.InfluenzavirusentryandinfectionrequirehostcellN-linkedglycoprotein.ProcNatlAcadSciUSA.2004;101(52):18153-18158.doi:10.1073/pnas.0405172102.
59. deVriesE,deVriesRP,WienholtsMJ,etal.InfluenzaAvirusentryintocellslackingsialylatedN-glycans.ProcNatlAcadSciUSA.2012;109(19):7457-7462.doi:10.1073/pnas.1200987109.

66
60. deVriesE,TscherneDM,WienholtsMJ,etal.DissectionoftheInfluenzaAVirusEndocyticRoutesRevealsMacropinocytosisasanAlternativeEntryPathway.PekoszA,ed.PLoSPathog.2011;7(3):e1001329.doi:10.1371/journal.ppat.1001329.
61. KarlssonHedestamGB,FouchierRAM,PhogatS,BurtonDR,SodroskiJ,WyattRT.ThechallengesofelicitingneutralizingantibodiestoHIV-1andtoinfluenzavirus.NatRevMicrobiol.2008;6(2):143-155.doi:10.1038/nrmicro1819.
62. OguinTH,SharmaS,StuartAD,etal.PhospholipaseDFacilitatesEfficientEntryofInfluenzaVirus,AllowingEscapefromInnateImmuneInhibition.JBiolChem.2014;289(37):25405-25417.doi:10.1074/jbc.M114.558817.
63. CrimMJ,RileyLK.Viraldiseasesinzebrafish:whatisknownandunknown.ILARJ.2012;53(2):135-143.doi:10.1093/ilar.53.2.135.
64. MarxM,RutishauserU,BastmeyerM.Dualfunctionofpolysialicacidduringzebrafishcentralnervoussystemdevelopment.Development.2001;128(24):4949-4958.http://www.ncbi.nlm.nih.gov/pubmed/11748132.AccessedNovember14,2016.
65. DehnertKW,BaskinJM,LaughlinST,etal.Imagingthesialomeduringzebrafishdevelopmentwithcopper-freeclickchemistry.Chembiochem.2012;13(3):353-357.doi:10.1002/cbic.201100649.
66. LanghauserM,UstinovaJ,Rivera-MillaE,etal.Ncam1aandNcam1b:twocarriersofpolysialicacidwithdifferentfunctionsinthedevelopingzebrafishnervoussystem.Glycobiology.2012;22(2):196-209.doi:10.1093/glycob/cwr129.
67. NeuU,BauerJ,StehleT.Virusesandsialicacids:rulesofengagement.CurrOpinStructBiol.2011;21(5):610-618.doi:10.1016/j.sbi.2011.08.009.
68. MatrosovichM,HerrlerG,KlenkHD.SialicAcidReceptorsofViruses.In:SpringerInternationalPublishing;2013:1-28.doi:10.1007/128_2013_466.

67
69. MM,AT,NB,etal.Earlyalterationsofthereceptor-bindingpropertiesofH1,H2,andH3avianinfluenzavirushemagglutininsaftertheirintroductionintomammals.JVirol.2000;74.
70. LevyDE,Garcı́a-SastreA.Thevirusbattles:IFNinductionoftheantiviralstateandmechanismsofviralevasion.CytokineGrowthFactorRev.2001;12(2):143-156.doi:10.1016/S1359-6101(00)00027-7.
71. GoodbournS,DidcockL,RandallRE.Interferons:Cellsignalling,immunemodulation,antiviralresponsesandviruscountermeasures.2000;81(10):2341-2364.
72. SchindlerC,BrutsaertS.Interferonsasaparadigmforcytokinesignaltransduction.1999;55(12):1509-1522.doi:10.1007/s000180050391.
73. HerbomelP,ThisseB,ThisseC.Ontogenyandbehaviourofearlymacrophagesinthezebrafishembryo.Development.1999;126(17):3735-3745.http://www.ncbi.nlm.nih.gov/pubmed/10433904.AccessedDecember13,2016.
74. LeGuyaderD,ReddMJ,Colucci-GuyonE,etal.Originsandunconventionalbehaviorofneutrophilsindevelopingzebrafish.Blood.2008;111(1):132-141.doi:10.1182/blood-2007-06-095398.
75. WillettCE,CherryJJ,SteinerLA.Characterizationandexpressionoftherecombinationactivatinggenes(rag1andrag2)ofzebrafish.Immunogenetics.1997;45(6):394-404.http://www.ncbi.nlm.nih.gov/pubmed/9089097.AccessedDecember13,2016.
76. DanilovaN,SteinerLA.Bcellsdevelopinthezebrafishpancreas.ProcNatlAcadSciUSA.2002;99(21):13711-13716.doi:10.1073/pnas.212515999.
77. LamSH,ChuaHL,GongZ,LamTJ,SinYM.Developmentandmaturationoftheimmunesysteminzebrafish,Daniorerio:ageneexpressionprofiling,insituhybridizationandimmunologicalstudy.DevCompImmunol.2004;28(1):9-28.http://www.ncbi.nlm.nih.gov/pubmed/12962979.AccessedDecember13,2016.

68
78. DirscherlH,McConnellSC,YoderJA,deJongJLO.TheMHCclassIgenesofzebrafish.DevCompImmunol.2014;46(1):11-23.doi:10.1016/j.dci.2014.02.018.
79. TakeuchiH,FigueroaF,O’hUiginC,KleinJ.CloningandcharacterizationofclassIMhcgenesofthezebrafish,Brachydaniorerio.Immunogenetics.1995;42:77-84.doi:10.1007/BF00178581.
80. RosenJN,SweeneyMF,MablyJD.Microinjectionofzebrafishembryostoanalyzegenefunction.JVisExp.2009;(25):2-5.doi:10.3791/1115.
81. TipsandTricksfortheLab:HowtoMakeaCapillaryTLCSpotter ::Education ::ChemistryViews.http://www.chemistryviews.org/details/education/2102991/Tips_and_Tricks_for_the_Lab_How_to_Make_a_Capillary_TLC_Spotter.html.AccessedJanuary3,2017.
82. KardashE.CurrentMethodsinZebrafishResearch.MaterMethods.2012;2.doi:10.13070/mm.en.2.109.
83. CuiC,BenardEL,KanwalZ,etal.InfectiousDiseaseModelingandInnateImmuneFunctioninZebrafishEmbryos.In:MethodsinCellBiology.Vol105.;2011:273-308.doi:10.1016/B978-0-12-381320-6.00012-6.
84. TechnologicalProgress-OurWorldInData.https://ourworldindata.org/technological-progress/.AccessedJanuary11,2017.
85. AlhnanMA,OkwuosaTC,SadiaM,WanK-W,AhmedW,ArafatB.Emergenceof3DPrintedDosageForms:OpportunitiesandChallenges.PharmRes.2016;33(8):1817-1832.doi:10.1007/s11095-016-1933-1.
86. 3DPrintingIndustryExamplesandCaseStudies.http://www.javelin-tech.com/3d-printer/industry/.AccessedJanuary15,2017.
87. Surgeon’sHelper:3DPrintingIsRevolutionizingHealthCare.http://www.livescience.com/49913-3d-printing-revolutionizing-health-care.html.AccessedJanuary15,2017.

69
88. RengierF,MehndirattaA,vonTengg-KobligkH,etal.3Dprintingbasedonimagingdata:reviewofmedicalapplications.IntJComputAssistRadiolSurg.2010;5(4):335-341.doi:10.1007/s11548-010-0476-x.
89. AlternateImmobilizationtechnique-MFchannels/Suction.http://www.pnas.org.prxy4.ursus.maine.edu/content/104/35/13891.full.pdf.AccessedMarch19,2016.
90. BischelLL,MaderBR,GreenJM,HuttenlocherA,BeebeDJ.ZebrafishEntrapmentByRestrictionArray(ZEBRA)device:alow-cost,agarose-freezebrafishmountingtechniqueforautomatedimaging.LabChip.2013;13(9):1732-1736.doi:10.1039/c3lc50099c.
91. RestoPJ,MogenB,WuF,BerthierE,BeebeD,WilliamsJ.Highspeeddroplet-baseddeliverysystemforpassivepumpinginmicrofluidicdevices.JVisExp.2009;(31):3-7.doi:10.3791/1329.
92. WesthoffJH,GiselbrechtS,SchmidtsM,etal.DevelopmentofanAutomatedImagingPipelinefortheAnalysisoftheZebrafishLarvalKidney.HwangS-PL,ed.PLoSOne.2013;8(12):e82137.doi:10.1371/journal.pone.0082137.
93. VeroClear,RGD810|Flavors|NA|Materials&Accessories|StratasyseStore.https://store.stratasys.com/stratasysstorefront/stratasys/en/USD/Materials-%26-Accessories/NA/Flavors/VeroClear%2C-RGD810/p/P030.AccessedFebruary22,2017.
94. BioCompatible,MED610|Flavors|NA|Materials&Accessories|StratasyseStore.https://store.stratasys.com/stratasysstorefront/stratasys/en/USD/Materials-%26-Accessories/NA/Flavors/Bio-Compatible%2C-MED610/p/P015.AccessedFebruary22,2017.
95. MacdonaldNP,ZhuF,HallCJ,etal.Assessmentofbiocompatibilityof3Dprintedphotopolymersusingzebrafishembryotoxicityassays.LabChip.2016;16(2):291-297.doi:10.1039/c5lc01374g.
96. OskuiSM,DiamanteG,LiaoC,etal.AssessingandReducingtheToxicityof3D-PrintedParts.EnvironSciTechnolLett.2016;3(1):1-6.doi:10.1021/acs.estlett.5b00249.

70
97. HoCMB,NgSH,LiKHH,YoonY-J.3Dprintedmicrofluidicsforbiologicalapplications.LabChip.2015;15(18):3627-3637.doi:10.1039/c5lc00685f.
98. VarmaS,VoldmanJ.Acell-basedsensoroffluidshearstressformicrofluidics.LabChip.2015;15(6):1563-1573.doi:10.1039/c4lc01369g.
99. PhennicieRT,SullivanMJ,SingerJT,YoderJA,KimCH.SpecificresistancetoPseudomonasaeruginosainfectioninzebrafishismediatedbythecysticfibrosistransmembraneconductanceregulator.InfectImmun.2010;78(11):4542-4550.doi:10.1128/IAI.00302-10.
100. ZFINTheZebrafishModelOrganismDatabase.http://zfin.org/.AccessedMarch22,2017.

71
APPENDIXA
ZEBRAFISHHUSBANDRY
ZebrafishwerehousedattheUniversityofMaineZebrafishFacilityin
recirculatingsystemswherethewatertemperaturewasmaintainedat28°Cwitha
totalsystemflowrateof150liters/min.Zebrafishweremaintainedinaccordance
withtheInstitutionalAnimalCareandUseCommitteestandardsoftheUniversityof
Maine.Fertilizedzebrafisheggsarecollectedaccordingtomethodsproposedby
PhennecieattheUniversityofMaineZebrafishFacility99.Collectedeggsareraised
for24hoursat28.0̊Cin100mm-diameterpetridisheshalffullofeggwater,60µg/L
InstantOceanSaltsinpurifiedreverseosmosiswater.Nomorethan100eggswere
raisedinone100mm-diameterpetridishatatime.
At24hourspostfertilization,eggwaterisreplacedineachpetridish.Chorions
areremovedfromtheembryosbyhandanddiscardedasbiologicalwaste.Live
embryosarestoredat28.0̊Cuntilthefollowingday.
At48hpf,embryosaremotile.Eggwaterisreplacedineachpetridishanddead
embryosareremovedfromthecontainer.Embryosarestoredat28.0̊Covernight.
At72hpf,specimensareconsideredlarvae.Zebrafisheggwaterisreplacedand
specimensareusedasneededinfurtherlaboratoryprocedures.
Zebrafishwereeuthanizedina600µg/LconcentratedstockofTricanefor
30minandfixedtominimizetheamountoflarvaeneededforrepeatedtesting.Fish
areremovedfromanestheticandplacedinfixativesolutionof1.5%gluteraldehyde,
0.5%para-formaldehyde,50mMPBS(phosphatebufferedsaline)forlongterm
storage100.Larvaewerefixedforatleast12hourspriortouse.Forflowtesting,

72
chemicallyfixedfishwereremovedfromfixativesolutionandplacedineggwater
withnomorethan0.25%gelatintominimizelarvaladhesiontotubing.
Forproceduresinvolvinglivespecimens,larvaeareanaesthetizedina
200µg/mLsolutionoftricanewithnomorethan0.25%gelatinaddedtominimize
specimenadhesiontotubing.Fishwereaspiratedthroughthedeviceinthe
anestheticgelatinsolution.

73
APPENDIXB
VALVE/SYRINGEPUMPCONTROLMAP
SETUP:
MyDAQDIOchannels(0-5)
• Input–laserdetector(detectedfish=1,nothingdetected=0)
• Output–valve1(open=1,close=0)
• Output–valve2(open=1,close=0)
• Output–valve3(open=1,close=0)
• Output–valve4(open=1,close=0)
• Output–valve5(open=1,close=0)
• Output–syringepumpdirection(inject=1,withdraw=0)
• Output–syringepumpstart/stop(start=1,stop=0)
ProgramCommands:
• Allvalvesclosed,pumpstopped
• Setpumptowithdrawmode
• Closevalve2,Closevalve4,Closevalve5(shouldbeclosedalready)
• Openvalve1andvalve3
• Startpump1
• Monitordetector
� Ifpositivedetect→stoppump,closevalve1,closevalve3,openvalve2,
openvalve3setpumptoinject,startpump
• Pumpinjects.Duration:(inputvalue)seconds→stoppump,valve2close

74
• Inject.Duration:(inputvalue)seconds
• Openvalve4,openvalve5
• Startpump2(infuse)
• Waitforfishtoflushout.Duration:(inputvalue)seconds
• Stoppump
• Closevalve5
• GotoStep1

75
APPENDIXC
MATERIALSLIST
PumpSystemMaterials:
� TygonMicroBoreTubing,Part#:TGY-030.ComponentSupplyCo.
� SiliconeTubing,Part#:SCT-063A.ComponentSupplyCo.
� BD60mLSyringe,Part#:309654.Becton,DickinsonandCompany.
� NewEraNE-1000XProgrammableSyringePump
� 2-waynormallyclosedsolenoidpinchvalve,Part#98302.ColePalmer
InstrumentCompany,LLC.
� AquaCultureSingleOutletAquariumAirPump,Part#0079285405132
OpticalDetectionMaterials
� 5mmtransparentcylinderheadphotodiode,Part#LLSO5-A.SenbaOptical
ElectricalCo.,Ltd
� 4.5V650nmredlaserdiodemodule,Part#01444878.LightInTheBoxCo.,
Ltd
� PROCELL1.5Vdrycellbatteries,Part#PC1500.DuracellInc.
EntrapmentDockMaterials
� Objet30RapidPrototypingSystem,SUP705SupportMaterial(WaterJet
Removable).StratasysLtd.
� CustomStageMount
� MakerBotReplicatorDesktop3DPrinter,PLAFilament.MakerBotIndustries,
LLC.

76
Electronics
� NationalInstrumentsmyDAQUnit
� SC5Five-ChannelSolenoidController/Driver.RWAutomations,LLC.
� SolidEdgeST7.SiemensProductLifecycleManagementSoftwareInc.
� NationalInstrumentsLabVIEW2016.NationalInstruments.

77
BIOGRAPHYOFTHEAUTHOR
FuoadSaliou-SulleywasborninAccra,Ghanaon16thJuly1991.Helived
withhisfamilyandattendedschoolinAccratillhewas17,whenhecametothe
UnitedStatesasanexchangestudent.Fuoadlivedwithahostfamilyin
Kennebunkport,MEandgraduatedfromKennebunkHighSchoolin2009.InMay
2014,hegraduatedMagnaCumLaudefromtheUniversityofMaineinOronowitha
degreeinBiologyandaminorinChemistry.Inthefallof2014,heenrolledinthe
schoolofChemicalandBiologicalEngineeringattheUniversityofMainedoing
researchinthelaboratoryofDr.PaulJ.Millard.
Fuoadintendstocontinuepursuingscientificresearchstudies,particularlyin
abiomedicalorpharmaceuticalinstitution.Wheneverhehasachanceto,Fuoad
likestospendtimewithfriends.Heisalsoveryfondofoutdoorsportsandactivities
likehiking,mountainbiking,soccerandbasketball.HeisacandidatefortheMaster
ofSciencedegreeinBiologicalEngineeringfromtheUniversityofMaineinAugust
2017.


















