Altered heart and kidney phospholipid fatty acid composition are associated with cardiac hypertrophy...
7
Altered heart and kidney phospholipid fatty acid composition are associated with cardiac hypertrophy in hypertensive rats Oh Yoen Kim a , Young-Sang Jung b , Yoonsu Cho c , Ji Hyung Chung d , Geum-Sook Hwang b, ⁎, Min-Jeong Shin c, ⁎⁎ a Department of Food Science and Nutrition, Dong-A University, Busan 604-714, Republic of Korea b Korea Basic Science Institute, Seoul 136-703, Republic of Korea c Department of Food and Nutrition, Korea University, Seoul 136-703, Republic of Korea d Yonsei Cardiovascular Hospital and Cardiovascular Research Institute, Yonsei University College of Medicine, Seoul 120-752, Republic of Korea abstract article info Article history: Accepted 6 April 2013 Available online 19 April 2013 Keywords: Hypertension Fatty acid Cardiac hypertrophy Left ventricle Kidney Objective: We examined the association of cardiac hypertrophy or fibrosis with the phospholipid fatty acid (FA) composition of heart and kidney in hypertensive rats. Design and methods: Eight-week-old spontaneously hypertensive rats (SHRs) (n = 8) and Wistar Kyoto rats (WKYs, n = 8) as a normotensive control, were fed ad libitum for 6 weeks with regular AIN-76 diet. Phospholipid FA compositions in the left ventricle and kidney were measured and histological analyses were performed. Results: Compared with WKYs, SHRs had lower proportions of γ-linolenic acid, α-linolenic acid, eicosadienoic acid, eicosatrienoic acid, dihomo-γ-linoleic acid, docosadienoic acid and nervonic acid in heart, and stearic acid (SA), γ-linolenic acid, and eicosapentaenoic acid (EPA) in kidney. After adjusting for food intake, SHRs still maintained higher proportions of SA, and total saturated FAs in the heart and a lower proportion of eicosapentaenoic acid in the kidney. Additionally, compared with WKYs, SHRs showed larger cardiomyocyte diameters in the left ventricles, indicating cardiac hypertrophy and interstitial fibrosis. Cardiomyocyte diameters also positively correlat- ed with cardiac SA (r = 0.550, p b 0.05) and negatively with kidney EPA (r = −0.575, p b 0.05). Conclusion: Tissue FA compositions were associated with cardiac hypertrophy in a hypertensive setting, impli- cating the pathogenic role of tissue FAs in hypertension and related complications. © 2013 The Canadian Society of Clinical Chemists. Published by Elsevier Inc. All rights reserved. 1. Introduction Abnormal fatty acid (FA) metabolism has been implicated in several health conditions such as hypertension (HTN), metabolic syndrome (MetS), diabetes mellitus, and cardiac failure. High concentrations of palmitic acid (C16:0) and low concentrations of linoleic acid (C18:2ω6, LA) in serum phospholipids (PL) are observed in individuals with insulin resistance and MetS [1,2]. Long-chain polyunsaturated fatty acid (PUFA) intake has also been associated with a decreased risk of cardiovascular disease [3,4]. Alterations in FA of membrane PLs affect membrane proper- ties and functions. For example, reduced unsaturated FA and membrane fluidity have been reported in vascular smooth muscle cells of patients with hypertension [5] and abnormal FA profiles are also observed in various tissues (i.e., kidney, medulla, liver) from spontaneously hyperten- sive rats (SHR) [6–8]. Additionally, several studies have reported that alterations in FA of membrane PLs are associated with severity of aortic stiffness, which is a clinical feature of HTN [9,10]. According to Kim et al. [11], brachial-ankle pulse wave velocity, a simple and non-invasive index of arterial stiffness, is highest in individuals with a lower proportion of LA and a higher proportion of dihomo-γ-linolenic acid (C20:3ω-6, DGLA) in serum PLs. Mori et al. also showed that docosahexaenoic acid lowers blood pressure (BP) by influencing endothelial-dependent or independent relaxation of forearm blood vessels [12]. In this regard, blood and tissue FA composition is as an important factor for detecting and managing HTN. Cardiac hypertrophy along with fibrosis is a severe form of hyperten- sive vascular disease because of its continuous progression into cardiac failure even after treatment of HTN. Furthermore, a prolonged hyperten- sive condition manifested by increased blood pressure damages tissue structures and causes functional consequences such as cardiac failure [13] and kidney disease. Dietary fat could change myocardial structure in obese animals [14]. That study suggested that modulated dietary fat results in increased myocyte size, contributing to early left ventricle (LV) thickening in obese patients [14]. However, no information is available on the association between tissue FA composition and cardiac hypertrophy in a hypertensive setting. Clinical Biochemistry 46 (2013) 1111–1117 Abbreviations: ALNA, α-linolenic acid; BP, blood pressure; DGLA, dihomo-γ-linolenic acid; EPA, eicosapentaenoic acid; ETA, eicosatrienoic acid; FA, fatty acid; HTN, hypertension; LV, left ventricle; MetS, metabolic syndrome; PL, phospholipid; PUFA, polyunsaturated fatty acid; SA, stearic acid; SFA, saturated fatty acid. ⁎ Correspondence to: G.-S. Hwang, Korea Basic Science Institute, Seoul 136-713 and Graduate School of Analytical Science and Technology, Chungnam National University, Daejeon 305-764, Republic of Korea. Fax: +82 2 920 0779. ⁎⁎ Correspondence to: M.-J. Shin, Department of Food and Nutrition, Korea University, Seoul 136-703, Republic of Korea. Fax: +82 2 940 2850. E-mail addresses: [email protected] (G.-S. Hwang), [email protected] (M.-J. Shin). 0009-9120/$ – see front matter © 2013 The Canadian Society of Clinical Chemists. Published by Elsevier Inc. All rights reserved. http://dx.doi.org/10.1016/j.clinbiochem.2013.04.008 Contents lists available at SciVerse ScienceDirect Clinical Biochemistry journal homepage: www.elsevier.com/locate/clinbiochem
Transcript of Altered heart and kidney phospholipid fatty acid composition are associated with cardiac hypertrophy...

Clinical Biochemistry 46 (2013) 1111–1117
Contents lists available at SciVerse ScienceDirect
Clinical Biochemistry
j ourna l homepage: www.e lsev ie r .com/ locate /c l inb iochem
Altered heart and kidney phospholipid fatty acid composition areassociated with cardiac hypertrophy in hypertensive rats
Oh Yoen Kim a, Young-Sang Jung b, Yoonsu Cho c, Ji Hyung Chung d, Geum-Sook Hwang b,⁎, Min-Jeong Shin c,⁎⁎a Department of Food Science and Nutrition, Dong-A University, Busan 604-714, Republic of Koreab Korea Basic Science Institute, Seoul 136-703, Republic of Koreac Department of Food and Nutrition, Korea University, Seoul 136-703, Republic of Koread Yonsei Cardiovascular Hospital and Cardiovascular Research Institute, Yonsei University College of Medicine, Seoul 120-752, Republic of Korea
Abbreviations:ALNA,α-linolenic acid; BP, bloodpressureEPA, eicosapentaenoic acid; ETA, eicosatrienoic acid; FA, faleft ventricle; MetS, metabolic syndrome; PL, phospholipacid; SA, stearic acid; SFA, saturated fatty acid.⁎ Correspondence to: G.-S. Hwang, Korea Basic Scienc
Graduate School of Analytical Science and Technology, CDaejeon 305-764, Republic of Korea. Fax: +82 2 920 07⁎⁎ Correspondence to: M.-J. Shin, Department of Food aSeoul 136-703, Republic of Korea. Fax: +82 2 940 2850
E-mail addresses: [email protected] (G.-S. Hwang), m
0009-9120/$ – see front matter © 2013 The Canadian Shttp://dx.doi.org/10.1016/j.clinbiochem.2013.04.008
a b s t r a c t
a r t i c l e i n f oArticle history:
Accepted 6 April 2013Available online 19 April 2013Keywords:HypertensionFatty acidCardiac hypertrophyLeft ventricleKidney
Objective:Weexamined the associationof cardiac hypertrophyorfibrosiswith the phospholipid fatty acid (FA)composition of heart and kidney in hypertensive rats.
Design andmethods: Eight-week-old spontaneously hypertensive rats (SHRs) (n = 8) and Wistar Kyoto rats(WKYs, n = 8) as a normotensive control, were fed ad libitum for 6 weeks with regular AIN-76 diet. PhospholipidFA compositions in the left ventricle and kidney were measured and histological analyses were performed.
Results: Compared with WKYs, SHRs had lower proportions of γ-linolenic acid, α-linolenic acid, eicosadienoicacid, eicosatrienoic acid, dihomo-γ-linoleic acid, docosadienoic acid and nervonic acid in heart, and stearic acid(SA), γ-linolenic acid, and eicosapentaenoic acid (EPA) in kidney. After adjusting for food intake, SHRs stillmaintainedhigher proportions of SA, and total saturated FAs in theheart and a lowerproportion of eicosapentaenoic
acid in the kidney. Additionally, compared with WKYs, SHRs showed larger cardiomyocyte diameters in the leftventricles, indicating cardiac hypertrophy and interstitial fibrosis. Cardiomyocyte diameters also positively correlat-ed with cardiac SA (r = 0.550, p b 0.05) and negatively with kidney EPA (r = −0.575, p b 0.05).Conclusion: Tissue FA compositions were associated with cardiac hypertrophy in a hypertensive setting, impli-cating the pathogenic role of tissue FAs in hypertension and related complications.
© 2013 The Canadian Society of Clinical Chemists. Published by Elsevier Inc. All rights reserved.
1. Introduction
Abnormal fatty acid (FA) metabolism has been implicated in severalhealth conditions such as hypertension (HTN), metabolic syndrome(MetS), diabetes mellitus, and cardiac failure. High concentrations ofpalmitic acid (C16:0) and low concentrations of linoleic acid (C18:2ω6,LA) in serum phospholipids (PL) are observed in individuals with insulinresistance and MetS [1,2]. Long-chain polyunsaturated fatty acid (PUFA)intake has also been associated with a decreased risk of cardiovasculardisease [3,4]. Alterations in FA ofmembrane PLs affectmembrane proper-ties and functions. For example, reduced unsaturated FA and membranefluidity have been reported in vascular smooth muscle cells of patientswith hypertension [5] and abnormal FA profiles are also observed in
;DGLA, dihomo-γ-linolenic acid;tty acid; HTN, hypertension; LV,id; PUFA, polyunsaturated fatty
e Institute, Seoul 136-713 andhungnam National University,79.nd Nutrition, Korea University,[email protected] (M.-J. Shin).
ociety of Clinical Chemists. Publishe
various tissues (i.e., kidney,medulla, liver) from spontaneously hyperten-sive rats (SHR) [6–8]. Additionally, several studies have reported thatalterations in FA of membrane PLs are associated with severity of aorticstiffness, which is a clinical feature of HTN [9,10]. According to Kim et al.[11], brachial-ankle pulse wave velocity, a simple and non-invasiveindex of arterial stiffness, is highest in individualswith a lower proportionof LA and a higher proportion of dihomo-γ-linolenic acid (C20:3ω-6,DGLA) in serum PLs. Mori et al. also showed that docosahexaenoic acidlowers blood pressure (BP) by influencing endothelial-dependent orindependent relaxation of forearm blood vessels [12]. In this regard,blood and tissue FA composition is as an important factor for detectingand managing HTN.
Cardiac hypertrophy along with fibrosis is a severe form of hyperten-sive vascular disease because of its continuous progression into cardiacfailure even after treatment of HTN. Furthermore, a prolonged hyperten-sive condition manifested by increased blood pressure damages tissuestructures and causes functional consequences such as cardiac failure[13] and kidney disease. Dietary fat could change myocardial structurein obese animals [14]. That study suggested that modulated dietary fatresults in increased myocyte size, contributing to early left ventricle(LV) thickening in obese patients [14]. However, no information isavailable on the association between tissue FA composition and cardiachypertrophy in a hypertensive setting.
d by Elsevier Inc. All rights reserved.

1112 O.Y. Kim et al. / Clinical Biochemistry 46 (2013) 1111–1117
Therefore, we examined the PLFA composition of heart and kidneyin SHRs and compared that with a normotensive control, Wistar Kyoto(WKY) rats. We hypothesized that cardiac hypertrophy or fibrosismay be associatedwith alterations in cardiac or renal PLFA compositionin rats with HTN.
2. Materials and procedures
2.1. Animals and systolic blood pressure (SBP) measurement
Eight-week-oldmaleWKY rats and SHRswere purchased fromHarlanLaboratory (Barcelona, Spain). SHRs were used as the hypertensive ani-mal model and WKY rats were used as the normotensive controls. Therats were housed in a temperature (18–24 °C) and humidity (50–60%)controlled room. Rats were acclimated for 2 weeks and were fed adlibitum for 6 weeks with regular AIN-76 diet. Animal body weight anddiet consumption were measured weekly. SBP was measured with a tailcuff plethysmograph using a BP measuring system (LE5002, Panlab s.l.,Barcelona, Spain). Briefly, rats were warmed to 37 °C for 5 min andallowed to rest in a chamber. Occluding cuffs and pneumatic pulse trans-ducers were placed on the tail. Five readings were obtained for each rat,and the average value of three readings, after removing the highest andlowest, was considered the SBP. All experimental procedures wereapproved by the Committee on Animal Experimentation and Ethics ofKorea University.
2.2. Tissue collection
Rats were fasted overnight prior to sacrifice. To measure tissue PLFAacid composition, the heart and kidney were extracted and weighed,and the left ventricle (LV) was excised and weighed. The tissues wereimmediately stored in polyethylene tubes at−70 °C until analysis.
2.3. Heart and kidney PLFA composition
LV total lipids were extracted from frozen tissue using the methoddescribed by Jeckel et al., [14]. Briefly, 0.05 g of tissue was placed in aglass homogenizer. Mass spectrometry (MS) grade methanol (0.75 mL)and 2.5 mL methyl-tert-butyl ether were added, and the tissue was ho-mogenized briefly on ice. The samples were capped off under nitrogengas and incubated at room temperature for 1 h, followed by adding0.625 mL of MS grade water. After vortexing, the samples were cappedoff under nitrogen and incubated at room temperature for an additional10 min, then centrifuged at 1000 ×g relative centrifugal force for 10 min.The upper (organic) phase was collected, and the sample was driedunder a stream of nitrogen. Kidney total lipids were extracted accordingto the Folchmethod [15]. The tissue PL fractionwas isolated by thin-layerchromatography using a development solvent composed of hexane:diethyl ether: and acetic acid (80:20:2). The PL fractions were thenmethylated to fatty acid methyl esters (FAMEs) using the Lepage andRoy method [16]. The FAMEs of individual PLFAs were separated by gaschromatography (model 6890, Agilent Technologies Inc., Palo Alto, CA,USA) using a capillary column (Omegawax™ 250; 30 m, Supelco,Bellefonte, PA, USA), as described previously [17]. Peak retention timeswere obtained by comparison with known standards (37 componentFAME mix and PUFA-2, Supelco; GLC37, NuCheck Prep, Elysian, MN,USA) and analyzed with ChemStation software (Agilent Technologies).Tissue PLFAs were expressed as a percentage of total FAs.
2.4. Histological analysis
Five-μm-thick sections were cut from blocked tissue and stainedwith Masson's trichrome to quantify areas of interstitial fibrosis. Tissueslices were examined with a magnification of 200× (Olympus BX41,Olympus America Inc., Center Valley, PA, USA). At least 20 field imageswere randomly digitized with a light microscope coupled to a high
resolution color camera interfaced to a computer. A gridwith 10 intersec-tion points was superimposed on a digitized image, and one section wasselected to analyze each image. The amount of fibrosis was expressed asa percentage of the total area of the heart section. LV sections weresubjected to periodic acid-Schiff staining and analyzed by microscopy todetermine cardiomyocyte transverse diameters (Olympus BX41) at amagnification of 400×. The diameters of at least 50 myocytes with alongitudinal orientation were determined using the analySIS getITprogram (Olympus Soft Imaging Solution). Diameters were obtained bytaking the distance passing through the center of the nucleus to thelongest part of the myocyte.
2.5. Statistical analysis
Multivariate data analysis, principal component analysis (PCA), andpartial least squares-discriminant analysis (PLS-DA) were performedusing SIMCA-P (version 12, Umetrics, Umea, Sweden). PCA, unsupervisedpattern recognition (PR) method, was performed to examine intrinsicvariations in the data set, and PLS-DA, a supervised PR method, wasemployed to investigate the difference between the WKY and SHRgroups. The quality of the models was described by R2 and Q2 values. R2
is defined as the proportion of variance in the data explained by themodels and indicates goodness of fit, and Q2 is defined as the proportionof variance in the data predictable by the model and indicates pre-dictability. Loading plots and the variable influence on projection(VIP) were examined to identify which metabolites contributedmost to clustering. Statistical analysis, including the significancetests, was performed using SPSS version 11.0 for Windows (SPSSInc., Chicago, IL, USA). Results are presented as means ± standarddeviations. Normal distribution tests were conducted with theShapiro–Wilk method. If both variables were normally distributed(p > 0.05), then the t-test was applied, otherwise the Mann–WhitneyU-test was used. Differences between the groups after adjusting forvariables were further analyzed using a general linear model (GLM).Partial correlation coefficients were used to test the associations amongvariables. p b 0.05 was considered significant.
3. Results
3.1. Body weight, food intake, and SBP
The average initial body weight in the WKY group was signifi-cantly lower than that in the SHR group (WKY, 217.9 ± 9.7 g; SHR260.5 ± 11.5 g). At 6 weeks, cumulative body weight gain was lessin the SHR group compared to that in the WKY group. In contrast,the average daily food intake was higher in the SHR group (SHR,16.7 ± 1.5 g) than that in the WKY group (14.5 ± 1.2 g). SHRsdisplayed significantly higher SBP levels than those of WKY rats at6 weeks (WKY, 194.7 ± 23.5 mm Hg vs. SHR, 194.7 ± 23.5 mm Hg,p b 0.001).
3.2. Pattern analysis
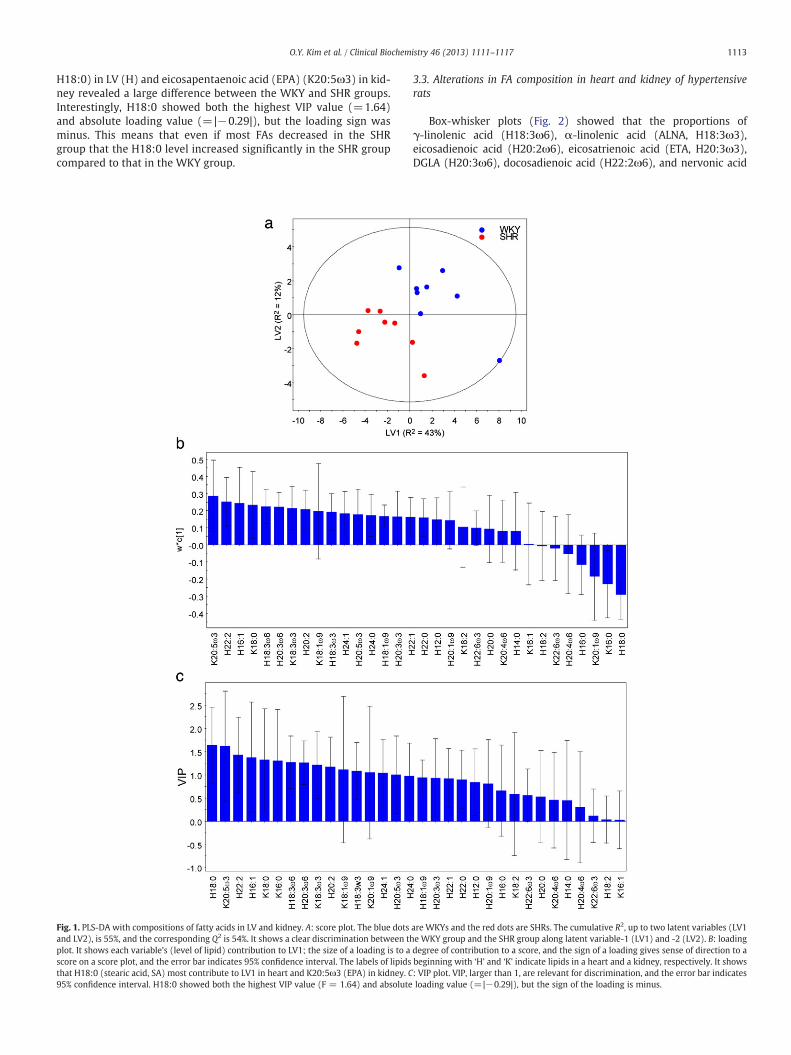
PCA was performed to examine the intrinsic variation and identifyoutliers. No strong outliers were observed in the PCA score plot,(p b 0.05) based on Hotelling's T2 (data not shown). PLS-DA was car-ried out to maximize separation between the groups. The PLS-DAscore plot showed a clear discrimination between the WKY and SHRgroups along latent variable 1 and 2 (Fig. 1A). R2 represents the good-ness of the fit for the PLS-DA model, and Q2 reveals the predictabilityof the PLS-DA model. The PLS-DA model revealed R2 and Q2 valuesof 55% and 54%, respectively, indicating high goodness of fit andpredictability for the PLS-DA model. PLS-DA loading and a VIPvalue were generated to identify the proportions of FA responsiblefor distinguishing between the two groups in the score plots. Asshown in Figs. 1B and 1C, the proportions of stearic acid (SA,
1113O.Y. Kim et al. / Clinical Biochemistry 46 (2013) 1111–1117
H18:0) in LV (H) and eicosapentaenoic acid (EPA) (K20:5ω3) in kid-ney revealed a large difference between the WKY and SHR groups.Interestingly, H18:0 showed both the highest VIP value (=1.64)and absolute loading value (=|−0.29|), but the loading sign wasminus. This means that even if most FAs decreased in the SHRgroup that the H18:0 level increased significantly in the SHR groupcompared to that in the WKY group.
Fig. 1. PLS-DA with compositions of fatty acids in LV and kidney. A: score plot. The blue dotsand LV2), is 55%, and the corresponding Q2 is 54%. It shows a clear discrimination between thplot. It shows each variable's (level of lipid) contribution to LV1; the size of a loading is to ascore on a score plot, and the error bar indicates 95% confidence interval. The labels of lipidsthat H18:0 (stearic acid, SA) most contribute to LV1 in heart and K20:5ω3 (EPA) in kidney. C95% confidence interval. H18:0 showed both the highest VIP value (F = 1.64) and absolute
3.3. Alterations in FA composition in heart and kidney of hypertensiverats
Box-whisker plots (Fig. 2) showed that the proportions ofγ-linolenic acid (H18:3ω6), α-linolenic acid (ALNA, H18:3ω3),eicosadienoic acid (H20:2ω6), eicosatrienoic acid (ETA, H20:3ω3),DGLA (H20:3ω6), docosadienoic acid (H22:2ω6), and nervonic acid
are WKYs and the red dots are SHRs. The cumulative R2, up to two latent variables (LV1e WKY group and the SHR group along latent variable-1 (LV1) and -2 (LV2). B: loadingdegree of contribution to a score, and the sign of a loading gives sense of direction to abeginning with ‘H’ and ‘K’ indicate lipids in a heart and a kidney, respectively. It shows: VIP plot. VIP, larger than 1, are relevant for discrimination, and the error bar indicatesloading value (=|−0.29|), but the sign of the loading is minus.

Fig. 2. Box-whisker plots and the significance tests with lipid levels between WKYs and SHRs. The bottom of the box is lower quartile (Q1, 25%), and the red line in the box is median (Q2, 50%), and the top of the box is upper quartile (Q3,75%), and the lower bar is the smallest observation among non-outliers, and the upper bar is largest observation among non-outliers. If observation is smaller than Q1 − 1.5 ∗ IQR or is larger than Q3 + 1.5 ∗ IQR, then it is considered out-liers; IQR (inter-quartile range) is defined as Q3–Q1, and the outlier is marked with a red cross. Normal distributions tests were performed with the Shapiro–Wilk method. If both variables were normally distributed (p > 0.05), then t-testwas applied; otherwise, Mann–Whitney U-test was done. The p-value indicates the difference of lipid levels between WKYs and SHRs is statistically significant (p b 0.05).
1114O.Y.K
imet
al./ClinicalBiochem
istry46
(2013)1111
–1117

Fig. 3. Proportions of fatty acids in LV and kidney between WKYs and SHRs after adjusting for food intake. Mean ± S.D. *p b 0.05, **p b 0.01, tested by general linear model (GLM)with adjustment for food intake.
1115O.Y. Kim et al. / Clinical Biochemistry 46 (2013) 1111–1117
(H24:1ω9) in LV were significantly lower in SHR compared to thosein WKY rats, whereas the proportion of SA (H18:0) was significantlyhigher in SHRs compared to that in WKY rats, Additionally, the pro-portions of SA (K18:0), ALNA (K18:3ω3), and EPA (K20:5ω3) in thekidneys of the SHR group were significantly lower than those in theWKY group. As dietary fat intake may modulate the incorporation ofFA in tissues [18], we compared FA tissue composition adjusted byfood intake using a GLM. FAs that were significantly different be-tween the groups in the significance test were selected for furtherGLM analyses. The results showed that the proportion of SA (H18:0)was significantly higher in SHRs compared to that in WKY rats afteradjusting for food intake (adjusted mean values for H18:0, SHRs:28.3 ± 2.08% vs. WKYs: 23.43 ± 2.08%, p = 0.003). The sum of
Fig. 4. Comparison of cardiac hypertrophy and interstitial fibrosis between WKYs and SHRsections were subjected to periodic acid-Schiff staining and analyzed by microscopy (OlymThe diameters of at least 50 myocytes with longitudinal orientation were determined using tby taking the distance passing through the center of the nucleus to the longest part of thestitial fibrosis. Five-μm-thick sections were cut from the blocked tissue and stained withwere examined with a magnification of 200× (Olympus BX41, Olympus America Inc., Cenmicroscope coupled to a high resolution color camera interfaced to computer. After a gridwas selected for analysis each image. The amount of fibrosis was expressed as percenta(nonpharametric t-test).
saturated fatty acids (ΔSFA) was also significantly higher in SHRscompared to that in WKY rats after adjusting for food intake (adjust-ed mean values for ΔSFA, SHRs: 48.3 ± 3.04% vs. WKYs: 43.6 ±3.04%, P = 0.023). As with the kidney, only a proportion of EPAremained to be significantly lower in SHRs after adjusting for food in-take (adjusted mean values for EPA (K20:5ω3), SHRs: 0.133 ± 0.015%vs. WKYs: 0.174 ± 0.015%, p = 0.004) (Fig. 3).
3.4. Cardiac hypertrophy and fibrosis in SHR
As shown in Fig. 4, periodic acid-Schiff staining of the myocardiumrevealed that the LV cardiomyocyte diameter was substantially largerin the SHR group than that in the WKY group. Interstitial fibrosis in
s. A: Cardiac hypertrophy. For cardiomyocyte transverse diameter determination, LVpus BX41, Olympus America Inc., Center Valley, PA, USA) at a magnification of 400×.he analySIS getIT program (Olympus Soft Imaging Solution). The diameter was obtainedmyocyte. *p b 0.05, tested by Mann–Whitney U-test (nonpharametric t-test). B: Inter-Masson's trichrome for quantification of the areas of interstitial fibrosis. Tissue slicester Valley, PA, USA). At least twenty field images were randomly digitized, via a lightwith 10 intersection points was superimposed on a digitized image then one sectionge of the total area of the heart section. *p b 0.05, tested by Mann–Whitney U-test

Table 1Relationships of heart and kidney fatty acid composition with cardiomyocyte diame-ters by partial correlation analysis.
Correlation coefficient (r) p-value
HeartC18:0, SA 0.550 0.042C18:3ω6, GLNA −0.454 0.103C18:3ω3, ALNA −0.191 0.513C20:1ω9, gondoic acid −0.513 0.061C20:2ω6, eicosadienoic acid −0.466 0.093C20:3ω6, DGLA −0.474 0.087C20:3ω3, 5-8-11-ETA −0.518 0.058C22:2ω6, docosadienoic acid −0.155 0.597
KidneyC18:0, SA −0.358 0.208C18:3ω3, ALNA −0.363 0.202C20:5ω3, EPA −0.575 0.032
GLNA:γ-linolenic acid (C18:3ω6); ALNA: α-linolenic acid (C18:3ω3); gondoic acid:cis-11-eicosenoic acid (C20:1ω9), eicosadienoic acid (C20:2ω6); ETA: eicosatrienoicacid (C20:3ω3); DGLA: dihomo-γ-linolenic acid (C20:3ω6).
1116 O.Y. Kim et al. / Clinical Biochemistry 46 (2013) 1111–1117
the LV was more extensive (p b 0.05) in the SHR group than that inthe WKY group.
3.5. Associations between tissue FA composition and cardiac hypertrophyand fibrosis
A partial correlation analysis after adjusting for food intake showedthat cardiomyocyte diameters were significantly related to SA in the LV(r = 0.550, p b 0.05) and EPA in the kidney (r = −0.575, p b 0.05)(Table 1). In addition, cardiomyocyte diameters tended to be negativelycorrelated with cardiac 5-8-11-ETA (H20:3ω3) and gondoic acid(cis-11-eicosenoic acid, H20:1ω9) but did not reach statistical signifi-cance. However, no significant relationships were observed betweenthemagnitude of fibrosis and the proportions of FAs in heart and kidney(Table 1).
4. Discussion
We showed significant differences in PLFA compositions in the heartand the kidney between SHRs and a normotensive control (WKY rats).It was also demonstrated that cardiac hypertrophy may be associatedwith alterations in cardiac and renal PLFA compositions, particularlySA and SFA in the heart and EPA in the kidney in SHRs.
Many in vivo and in vitro studies have reported abnormalities in lipidmetabolism in humans and animals suffering from genetic HTN [5–8].For example, changes in free cholesterol and PL composition of platelets,erythrocytes, and plasma lipoproteins have been reported in patientswith hypertension [19–21]. Abnormal PL composition was also reportedin jejunal brush-border membrane vesicles and heart cell cultures fromSHRs [22]. Additionally, alterations in PUFA metabolism are observed inthe heart, thoracic aorta, and liver from SHRs [9-23]. Huang et al. [24]and Chi et al. [9] found that the percentage of ω3-PUFAs is significantlylower in SHRs than that in WKY rats, although the percentage ofω9-PUFAs was higher in SHRs than that in WKY rats. Huang et al. alsoreported that the percentage ofω6-PUFAs is higher in heart membranesfrom SHRs than that from WKY rats and that no significant differencewas observed in the P/S ratio between the two strains [24]. In contrast,Chi et al. reported a decrease in the levels of FA unsaturation (doublebonds) in both the aorta and kidney of SHRs, and average FA chain lengthwas associated with HTN; they explained that these changes occurredbecause of altered turnover or modification of endogenous membranelipids, even when both normotensive and hypertensive animals werefed identical diets. In line with previous results [9-24], we found thatSHRs had significantly higher proportions of SA (C18:0) and lower pro-portions of several unsaturated FAs in the heart and kidney than thoseinWKY rats. After adjusting for food intake, SHRs still maintained higher
proportions of cardiac SA (C18:0) and total SFA and lower proportions ofkidney EPA than those in control WKY rats.
The exactmechanism by which abnormal lipid metabolism is associ-atedwith the development of HTN remains unclear, but several plausiblemechanisms can be addressed. Alterations inmembrane PLs in the heartand the kidney might contribute to the abnormal intracellular ionconcentrations and ion transport observed in the organs in patientswith HTN; thereby, causing significant modification of membranefunction, i.e., ion permeability, fluidity, and elasticity [25]. Alternatively,abnormalities in FA metabolism may have implications for local synthe-sis of eicosanoids, which greatly depend on the availability of C20 PUFAprecursors such as DGLA, arachidonic acid, and EPA. [26–28]. ThesePUFAprecursors, released frommembrane PLs, participate in the synthe-sis of vasoactive eicosanoids known as inflammatorymediators [26–28],which are involved in regulatingBP. Finally, changes inmyocellular lipidsare associated with altered intracellular signaling, including pathwaysthat may be important for modulating myocyte metabolism, hypertro-phy, contractile function, and survival [29,30]. In the present study, ahigher proportion of SFA and a lower proportion of ω3-PUFAs wereobserved in SHRs than those in the WKY rats. It can be speculated thatalterations in FA composition might partly participate in inflammatorypathways that can unfavorably influence the development and progres-sion of HTN.
Of interest is the finding that cardiomyocyte diameterwas associatedwith FA composition, whereas fibrosis was not. In particular, a higherproportion of SA, a kind of saturated fat, in LV of the heart was found inSHRs compared with that in the normotensive model, and the propor-tion of SAwas positively correlatedwith cardiomyocyte diameter. Sever-al studies have inconsistently reported a relationship between dietary fattype and cardiac hypertrophy. According to Jeckel et al., high fat intakecould modulate myocardial structure, causing enlargement of myocytesand contributing to early LV thickening, particularly in obese animals[14]. Thunder et al. also showed that the heart/bodyweight ratio is great-er in rats fed saturated fat diets compared with those fed unsaturatedfat after consuming an isoenergetic diet [31]. In contrast, Chess et al.suggested that increased saturated fat intake does not exacerbatepathological cardiac hypertrophy that develops in response to pressureoverload [32].While the association between dietary FAs and cardiachypertrophy has not been clearly demonstrated, several studies havesupported the notion that the type of dietary FAsmight influence cardiacfunction. Protein kinase C (PKC) plays a central role in modulatingcardiac hypertrophy in various kinds of animal and cell culture models[33] by modifying transcription factors and gene expression [34].Increased activation of PKCβ2 has been observed in rats fed saturatedfat diets [31], suggesting that dietary fat type can alter PKC β2 activationleading to a mild hypertrophic effect on the heart. It has also beenreported that saturated and ω6-PUFAs enhance myocyte apoptosis andnecrosis [35,36], whereas PUFAs and ω3-PUFAs attenuate apoptosis incardiac and endothelial cells [37]. Additionally, feedingω6-PUFAs to nor-mal pigs is associated with myocardial inflammation while feeding.ω3-PUFAs are associated with an anti-inflammatory effect [38]. Theseobservations together with our results raise the possibility that dietaryfat and/or tissue FA composition may differentially impact LV structureand contractile function. Further studies to examine the associationbetween FAs and cardiac hypertrophy are warranted.
In summary, our results confirmed that PL membrane FA composi-tions in the heart and the kidney were significantly different betweenhypertensive rats and normotensive controls even when dietary intakewas controlled. The results also show that cardiac hypertrophy, estimatedby cardiomyocyte diameter in the LV, was significantly associated withaltered cardiac (particularly SA and SFA) and renal PLFA compositions(particularly EPA) in the HTN rats. Our study has some limitations. First,we fed a controlled normal diet to our models, and did not try variouskinds of dietary FAs, which may have affected the alteration of PL mem-brane FA composition. Second, we used 8-week old rat models; thus,we could not observe the aging effect on PLFA composition. Considering

1117O.Y. Kim et al. / Clinical Biochemistry 46 (2013) 1111–1117
the discrepancy in study results among studies, further studies needto be conducted with various tissues or organ types, observationalperiods, age, gender, and feeding types. Despite these limitations,our study provides information on the association between tissueFA composition and cardiac hypertrophy in a hypertensive setting,implicating the pathogenic role of tissue FAs in HTN and relatedcomplications.
5. Conclusion
This study provided information for an association between tissueFA composition and cardiac hypertrophy in a HTN setting, implicatingthe pathogenic role of tissue FAs in HTN and related complications.
Conflict of interest
None.
Acknowledgment
This research was supported by the Basic Science Research Programthrough the National research Foundation of Korea (NRF) funded bythe Ministry of Education, Science and Technology (2012-0002119),the Creative Allied Project (CAP) grant funded by the Korea ResearchCouncil of Fundamental Science and Technology (KRCF) and KoreaBasic Science Institute (E33300).
References
[1] Vessby B. Dietary fat, fatty acid composition in plasma and the metabolic syndrome.Curr Opin Lipidol 2003;14:15–9.
[2] Leeson CP, Mann A, Kattenhorn M, Deanfield JE, Lucas A, Muller DP. Relationshipbetween circulating n − 3 fatty acid concentrations and endothelial function inearly adulthood. Eur Heart J 2002;23:216–22.
[3] Iso H, Kobayashi M, Ishihara J, Sasaki S, Okada K, Kita Y, et al. Intake of fish and n3fatty acids and risk of coronary heart disease among Japanese: the Japan PublicHealth Center-Based (JPHC) Study Cohort I. Circulation 2006;113:195–202.
[4] Hooper L, Thompson RL, Harrison RA, Summerbell CD, Ness AR, Moore HJ, et al. Risksand benefits of omega 3 fats for mortality, cardiovascular disease, and cancer: system-atic review. BMJ 2006;332:752–60.
[5] Tsuda K, Tsuda S, Minatogawa Y, Iwahashi H, Kido R, Masuyama Y. Decreased mem-brane fluidity of erythrocytes and cultured vascular smooth muscle cells in spontane-ously hypertensive rats: an electron spin resonance study. Clin Sci 1988;75:477–80.
[6] Singer P, Wirth M, Gerike U, Godicke W, Moritz V. Age dependent alterations oflinoleic, arachidonic and eicosapentaenoic acids in renal and medulla of sponta-neously hypertensive rats. Prostaglandins 1984;27:375–90.
[7] Watanabe Y, Huang YS, Simmons VA, Horrobin DF. The effect of dietary n − 6 andn − 3 polyunsaturated fatty acids on blood pressure and tissue fatty acid compositionin spontaneously hypertensive rats. Lipids 1989;24:638–44.
[8] Narce M, Poisson JP. Age-related depletion of linoleic acid desaturation in livermicrosomes from young spontaneously hypertensive rats. Prostaglandins LeukotEssent Fatty Acids 1995;53:59–63.
[9] Chi Y, Gupta RK. Alterations inmembrane fatty acid unsaturation and chain length inhypertension as observed by 1H NMR spectroscopy. A J Hypertens 1998;11:340–8.
[10] Vazques CM, Mate A, De la Hermosa MA, Plana J, Ruiz-Gutierrez V. Abnormalitiesin lipid composition of brush-border membranes isolated from renal cortex ofspontaneously hypertensive rat. Am J Hypertens 2001;14:578–84.
[11] Kim OY, Lim HH, Lee MJ, Kim JY, Lee JH. Association of fatty acid composition in serumphospholipids with metabolic syndrome and arterial stiffness. Nutr Metab CardiovascDis 2013;23:366–74.
[12] Mori TA, Bao DQ, Burke V, Puddey IB, Beilin LJ. Docosahexaenoic acid but noteicosapentaenoic acid lowers ambulatory blood pressure and heart rate in humans.Hypertension 2002;34:253–60.
[13] Chien KR, Knowlton KU, Zhu H, Chien S. Regulation of cardiac gene expression duringmyocardial growth and hypertrophy: molecular studies of an adaptive physiologicresponse. FASEB J 1991;5:3037–46.
[14] Jeckel KM, Miller KE, Chicco AJ, Chapman PL, Mulligan CM, Falcone PH, et al. Therole of dietary fatty acids in predicting myocardial structure in fat-fed rats. LipidsHealth Dis 2011;10:92.
[15] Folch JM, Lees GH, Stanley S. A simple method for the isolation and purification oftotal lipids from animal tissues. J Biol Chem 1957;226:497–509.
[16] Repage G, Roy CC. Direct transesterification of all classes of lipids in a one-stepreaction. J Lipid Res 1986;27:114–20.
[17] Lemaitre R, King IB, Mozaffarian M, Sotoodehnia N, Rea TD, Kuller LH, et al. Plasmaphospholipid trans fatty acids, fatal ischemic heart disease, and sudden cardiacdeath in older adults. Circulation 2006;114:209–315.
[18] Katan MB, van Birgelen AP, Penders M, Zegwaard M. Kinetics of the incorporationof dietary fatty acids into serum cholesteryl esters, erythrocyte membranes, andadipose tissue: an 18-month controlled study. J Lipid Res 1997;38:2012–22.
[19] Benjamin N, Graham JM, Robinson BF, Wilson RB. Cholesterol phospholipid ratiois elevated in platelets plasma membrane in patient with hypertension. J HumHypertens 1990;4:273–6.
[20] Annapurna V, Puniyani RR, Gupte RV. Red cell deformability and erythrocytelipids in hypertension. Clin Hemorheol 1990;10:95–8.
[21] Bagdale JD, Buchanan WF, Pollare T, Lithell H. Abnormal lipoprotein phospholipidcomposition in patientswith essential hypertension. Atherosclerosis 1995;117:209–15.
[22] Vazques CM, Zanetti R, Ruiz-Guierrez V. Lipid composition and fluidity in the jejunalbrush-border membrane of spontaneously hypertensive rats: effect on activities ofmembrane-bound proteins. Biosci Rep 1996;16:217–26.
[23] DelachambreMC, Marce M, Asdrubal P, Poisson JP. Changes in tissue polyunsaturat-ed fatty acids with age in spontaneously hypertensive rats. Lipids 1998;33:795–801.
[24] Huang Y, Mills DE, Ward RP, Horrobin DF, Simmons VA. Effect of essential fattyacid depletion on tissue phospholipid fatty acids in spontaneously hypertensiveand normotensive rats. Lipids 1989;24:565–71.
[25] Chi Y, Gupta RK. Alterations in heart and kidneymembrane phospholipids in hyperten-sion as observed by 31P nuclear magnetic resonance. Lipids 1998;33:1023–30.
[26] Sasrdesai VM. Nutritional role of polyunsaturated fatty acids. J Nutr Biochem1992;3:154–66.
[27] Oates JA, Fitzgerald GA, Branch RA, Jackson EK, Knapp HR, Robert LJ. Clinical implica-tions of prostaglandin and thromboxane A2 formation. N Engl JMed 1988;391:689–98.
[28] Kirtland SJ. Prostaglandin E1: a review. Prostaglandins Leukotreines Essent FattyAcids 1988;32:165–74.
[29] Jalili T, Manning J, Kim S. Increased translocation of cardiac protein kinase C beta2 ac-companies mild cardiac hypertrophy in rats fed saturated fat. J Nutr 2003;133:358–61.
[30] Akki A, Seymour AM. Western diet impairs metabolic remodelling and contractileefficiency in cardiac hypertrophy. Cardiovasc Res 2009;81:610–7.
[31] Thunder J, Manning J, Kim S. Increased translocation of cardiac protein kinase C β2accompanies mild cardiac hypertrophy in rats fed saturated fat. J Nutr 2003;133:358–61.
[32] Chess D, Lei B, Hoit B, Azimzadeh AM, Stanley W. Effects of a high saturated fatdiet on cardiac hypertrophy and dysfunction in response to pressure overload. JCard Fail 2008;14:82–8.
[33] Karns LR, Kariya K, Simpson PC. M-CAT, CArG, and Sp1 elements are required foralpha 1-adrenergic induction of the skeletal alpha-actin promoter during cardi-ac myocyte hypertrophy. Transcriptional enhancer factor-1 and protein kinase Cas conserved transducers of the fetal program in cardiac growth. J Biol Chem1995;270:410–7.
[34] Chien KR, Knowlton KU, Zhu H, Chien S. Regulation of cardiac gene expressionduring myocardial growth and hypertrophy: molecular studies of an adaptivephysiologic response. FASEB J 1991;5:3037–46.
[35] Ghosh S, An D, Pulinilkunnil T, Qi D, Lau HC, Abrahani A, et al. Role of dietary fatty acidsand acute hyperglycemia in modulating cardiac cell death. Nutrition 2004;20:916–23.
[36] de Vries JE, Vork MM, Roemen TH, de Jong YF, Cleutjens JP, van der Vusse GJ, et al.Saturated but not mono-unsaturated fatty acids induce apoptotic cell death inneonatal rat ventricular myocytes. J Lipid Res 1997;38:1384–94.
[37] Pfrommer CA, Erl W, Weber PC. Docosahexaenoic acid induces ciap1 mRNA andprotects human endothelial cells from stress-induced apoptosis. Am J PhysiolHeart Circ Physiol 2006;290:2178–86.
[38] DudaMK, O'Shea KM, Tintinu A, XuW, Khairallah RJ, Barrows BR, et al. Fish oil, but notflaxseed oil, decreases inflammation and prevents pressure overload-induced cardiacdysfunction. Cardiovasc Res 2008;81:319–27.