
Aktivitas Anticendawan Bacillus cereus 11UJ terhadap...
38
Volume 11, Nomor 2, April 2015 Halaman 35–42 DOI: 10.14692/jfi.11.2.35 ISSN: 0215-7950 35 Aktivitas Anticendawan Bacillus cereus 11UJ terhadap Rhizoctonia solani dan Pyricularia oryzae Antifungal Activity of Bacillus cereus 11UJ against Rhizoctonia solani and Pyricularia oryzae Yadi Suryadi*, I Made Samudra, Tri Puji Priyatno, Dwi Ningsih Susilowati, Puji Lestari, Sutoro Balai Besar Penelitian Bioteknologi dan Sumberdaya Genetik Pertanian, Bogor 16111 ABSTRAK Metabolit sekunder asal mikrob merupakan sumber senyawa penting untuk pengembangan senyawa anticendawan pada strategi pengendalian penyakit. Penelitian ini bertujuan menguji kemampuan produksi metabolit sekunder bakteri antagonis rizosfer Bacillus cereus 11UJ terhadap cendawan patogen padi Rhizoctonia solani dan Pyricularia oryzae. Efek antagonis ekstrak kasar B. cereus 11UJ diuji dengan metode cakram kertas saring steril pada medium PDA. Ekstrak bakteri dengan pelarut etil asetat menunjukkan aktivitas anticendawan yang lebih baik terhadap P. oryzae dibandingkan dengan R. solani. Efek penghambatan filtrat membuktikan potensi B. cereus 11UJ untuk aktivitas anticendawan. Analisis pirolisis gas kromatografi spektrometri massa menunjukkan bahwa B. cereus 11UJ menghasilkan 3 senyawa utama, yaitu 9,19-cyclolanostan-3-ol, acetate, (3.beta.)-(CAS) cycloartanyl acetate (13.14%); 4-(2’,2’-dimethyl-6’-methyliden-1’-cyclohexyliden)-3-methyl-2-butanone (9.72%); dan stigmast-5-en- 3-ol, oleat (9.09%) yang diduga berpotensi menekan pertumbuhan patogen. Kata kunci: antagonis, gas kromatografi spektrometri massa, metabolit sekunder, patogen padi ABSTRACT Microbial secondary metabolites is an important resource for antifungal development in disease control strategy. The objective of this study was to screen Bacillus cereus 11UJ, an antagonistic rhizosphere bacteria for potential secondary metabolite production against rice fungal pathogens, i.e. Rhizoctonia solani and Pyricularia oryzae. The antagonistic effect of crude exract was evaluated using sterile filter paper discs on PDA medium. The ethyl acetate extracts of the bacterium showed a better antifungal activity to P. oryzae than those of R. solani. The inhibitory effect of the filtrate proved the potency of the isolates to produce antifungal. Analysis of pyrolysis gas chromatography-mass spectrometry showed that B. cereus 11UJ produces 3 major compounds i.e; 9,19-cyclolanostan-3-ol, acetate, (3.beta.)- (CAS) cycloartanyl acetate (13.14%); 4-(2’,2’-dimethyl-6’-methyliden-1’-cyclohexyliden)-3-methyl-2- butanone (9.72%); and stigmast-5-en-3-ol oleat (9.09%) which suggested to play an important role in the suppression of rice fungal pathogens. Key words: antagonists, gas chromatography mass spectrometry, rice pathogens, secondary metabolytes *Alamat penulis korespondensi: Balai Besar Penelitian dan Pengembangan Bioteknologi dan Sumberdaya Genetik Pertanian, Jalan Tentara Pelajar 3A, Bogor 16111 Tel: 0251-8338820, Faks: 0251-8338820, Surel: [email protected]
-
Upload
truongtuyen -
Category
Documents
-
view
213 -
download
0
Transcript of Aktivitas Anticendawan Bacillus cereus 11UJ terhadap...

Volume 11, Nomor 2, April 2015Halaman 35–42
DOI: 10.14692/jfi.11.2.35ISSN: 0215-7950
35
Aktivitas Anticendawan Bacillus cereus 11UJ terhadap Rhizoctonia solani dan Pyricularia oryzae
Antifungal Activity of Bacillus cereus 11UJ against Rhizoctonia solani and Pyricularia oryzae
Yadi Suryadi*, I Made Samudra, Tri Puji Priyatno, Dwi Ningsih Susilowati, Puji Lestari, Sutoro
Balai Besar Penelitian Bioteknologi dan Sumberdaya Genetik Pertanian, Bogor 16111
ABSTRAK
Metabolit sekunder asal mikrob merupakan sumber senyawa penting untuk pengembangan senyawa anticendawan pada strategi pengendalian penyakit. Penelitian ini bertujuan menguji kemampuan produksi metabolit sekunder bakteri antagonis rizosfer Bacillus cereus 11UJ terhadap cendawan patogen padi Rhizoctonia solani dan Pyricularia oryzae. Efek antagonis ekstrak kasar B. cereus 11UJ diuji dengan metode cakram kertas saring steril pada medium PDA. Ekstrak bakteri dengan pelarut etil asetat menunjukkan aktivitas anticendawan yang lebih baik terhadap P. oryzae dibandingkan dengan R. solani. Efek penghambatan filtrat membuktikan potensi B. cereus 11UJ untuk aktivitas anticendawan. Analisis pirolisis gas kromatografi spektrometri massa menunjukkan bahwa B. cereus 11UJ menghasilkan 3 senyawa utama, yaitu 9,19-cyclolanostan-3-ol, acetate, (3.beta.)-(CAS) cycloartanyl acetate (13.14%); 4-(2’,2’-dimethyl-6’-methyliden-1’-cyclohexyliden)-3-methyl-2-butanone (9.72%); dan stigmast-5-en-3-ol, oleat (9.09%) yang diduga berpotensi menekan pertumbuhan patogen.
Kata kunci: antagonis, gas kromatografi spektrometri massa, metabolit sekunder, patogen padi
ABSTRACT
Microbial secondary metabolites is an important resource for antifungal development in disease control strategy. The objective of this study was to screen Bacillus cereus 11UJ, an antagonistic rhizosphere bacteria for potential secondary metabolite production against rice fungal pathogens, i.e. Rhizoctonia solani and Pyricularia oryzae. The antagonistic effect of crude exract was evaluated using sterile filter paper discs on PDA medium. The ethyl acetate extracts of the bacterium showed a better antifungal activity to P. oryzae than those of R. solani. The inhibitory effect of the filtrate proved the potency of the isolates to produce antifungal. Analysis of pyrolysis gas chromatography-mass spectrometry showed that B. cereus 11UJ produces 3 major compounds i.e; 9,19-cyclolanostan-3-ol, acetate, (3.beta.)- (CAS) cycloartanyl acetate (13.14%); 4-(2’,2’-dimethyl-6’-methyliden-1’-cyclohexyliden)-3-methyl-2-butanone (9.72%); and stigmast-5-en-3-ol oleat (9.09%) which suggested to play an important role in the suppression of rice fungal pathogens.
Key words: antagonists, gas chromatography mass spectrometry, rice pathogens, secondary metabolytes
*Alamat penulis korespondensi: Balai Besar Penelitian dan Pengembangan Bioteknologi dan Sumberdaya Genetik Pertanian, Jalan Tentara Pelajar 3A, Bogor 16111 Tel: 0251-8338820, Faks: 0251-8338820, Surel: [email protected]

J Fitopatol Indones Suryadi et al.
36
PENDAHULUAN
Penyakit penting pada padi di antaranya ialah penyakit hawar pelepah daun dan blas yang disebabkan berturut-turut oleh Rhizoctonia solani dan Pyricularia oryzae. Kedua cendawan patogen ini mempunyai banyak galur sehingga sangat sulit dikendalikan. Penggunaan pestisida untuk mencegah serangan penyakit pada tanaman sampai sekarang masih banyak digunakan. Beberapa bahan pestisida bersifat persisten dan tidak mudah diurai oleh mikroorganisme sehingga dapat mengakibatkan pencemaran lingkungan dan gangguan kesehatan manusia. Salah satu alternatif pengendalian yang ramah lingkungan ialah pemakaian agens hayati.
Saat ini galur bakteri Pseudomonas spp. dan cendawan Trichoderma spp. antagonis banyak mendapat perhatian (Kang 2012; Gomathi et al. 2013). Kedua mikrob tersebut dilaporkan mampu menghasilkan senyawa yang tidak menguap maupun senyawa yang mudah menguap dan menunjukkan aktivitas penghambatan kuat terhadap cendawan patogen tanaman. Galur Bacillus sp. (Jing dan Qian 2007) dan B. subtilis (Muskhazli et al. 2007) juga dilaporkan menghasilkan senyawa terlarut yang tidak menguap serta memiliki aktivitas anticendawan yang tinggi. Bakteri ini juga dapat berperan dalam menekan beberapa cendawan patogen seperti Rhizoctonia dan Fusarium (El-hamshary dan Khattab 2008; Soria et al. 2012; Sadrati et al. 2013; Wen et al. 2011). Spora bakteri dari beberapa jenis Bacillus sp. (B. cereus, B. clausii, dan B. pumilus) yang telah dikarakterisasi banyak dimanfaatkan untuk uji aktivitas anticendawan (Płaza et al. 2012).
Bacillus cereus 11UJ yang diisolasi dari tanah rizosfer tanaman tebu di daerah Jember dilaporkan dapat mengatalisis kitin yang terdapat pada dinding sel cendawan (Suryadi et al. 2013). Produksi senyawa metabolit sekunder yang dihasilkan B. cereus yang telah diuji secara in vitro terhadap cendawan patogen kemungkinan dapat dimanfaatkan sebagai sumber bahan anticendawan (Romeiro et al. 2010). Hasil penelitian Fiddaman dan Rossall (1994) menemukan senyawa mudah
menguap yang dihasilkan oleh bakteri B. subtilis dapat menghambat pertumbuhan dan perkecambahan spora cendawan patogen sehingga dapat menjadi agens pengendali hayati terhadap beberapa penyakit cendawan tular tanah.
Penelitian ini bertujuan menentukan aktivitas anticendawan yang dihasilkan B. cereus 11UJ terhadap R. solani dan P. oryzae, serta senyawa metabolit sekunder yang dihasilkannya.
BAHAN DAN METODE
Peremajaan dan Kultur Bakteri Bacillus cereus 11 UJ
Bacillus cereus 11UJ yang digunakan berasal dari koleksi Biogen-Culture Collection, Bogor. Cendawan R. solani dan P. oryzae asal tanaman padi digunakan sebagai cendawan uji. B. cereus 11UJ diremajakan pada medium agar-agar nutrien (AN) dan diinkubasi selama 24 jam pada suhu kamar (22 °C) selama 1 minggu. Selanjutnya biakan diperbanyak dalam 200 mL medium nutrien cair (nutrient broth). Suspensi dikocok menggunakan orbital shaker (SI 50, USA), kemudian masing-masing dimasukkan ke dalam 4 mL tabung ulir. Biakan bakteri diinkubasi selama 24–48 jam dalamalat kocok (IKA, KS 200) dengan kecepatan 120 rpm, selanjutnya suspensi bakteri di-kumpulkan dalam erlenmeyer 250 mL untuk dilakukan fraksinasi.
Fraksinasi Senyawa Anticendawan dengan Evaporator
Suspensi bakteri diekstraksi dalam bentuk fase cair menggunakan pelarut etil asetat untuk memperoleh metabolitnya. Medium cair berisi B. cereus 11UJ disentrifugasi menggunakan Himac CR21F (Hitachi) dengan kecepatan 10 000 × g pada suhu 4 °C selama 20 menit. Supernatan disaring menggunakan filter steril (0.45 µm). Supernatan kemudian diekstrak kembali dengan larutan etil asetat dengan perbandingan supernatan dan pelarut 1:1 sehingga terjadi fraksinasi antara fraksi etil asetat dan fraksi air. Eluat (fraksi air yang terlarut dalam medium dari proses inkubasi)

J Fitopatol Indones Suryadi et al.
37
kemudian dievaporasi menggunakan alat rotary evaporator (EYELA, Tokyo Ltd) pada suhu 50 °C hingga konsentrat berkurang 10% dari volume awal, pH diatur hingga 3.6 dengan HCl 1N, lalu fraksi air diekstrak kembali sebanyak 3 kali dengan perbandingan supernatan dan pelarut etil asetat 1:1. Selanjutnya fraksi air diambil dan diuapkan kembali di atas hot plate dengan suhu 50 °C sampai didapat hasil ekstrak kasar.
Uji Aktivitas Senyawa Metabolit B. cereus 11UJ terhadap R. solani dan P. oryzae
Uji antagonis senyawa metabolit asal B. cereus 11UJ terhadap cendawan dilakukan berdasarkan metode difusi sumur. Hasil ekstrak kasar sebanyak 1 mg dilarutkan dalam larutan 0.1% Tween 20 dan diencerkan dengan konsentrasi 1000, 500, 250, dan 100 ppm. Ekstrak metabolit tersebut masing-masing diuji dalam medium agar-agar dekstrosa kentang yang sudah disiapkan, kecuali untuk kontrol tidak diinokulasi metabolit (hanya air steril). Masing-masing perlakuan diulang sebanyak 4 kali. Bakteri B. cereus 11UJ diuji tantang dengan R. solani dan P. oryzae dan diinkubasi pada suhu 28 °C selama 5–7 hari. Daya anticendawan diamati setelah terlihat adanya aktivitas terhadap R. solani dan P. oryzae, dan zona hambat diamati pada 7 hari setelah inkubasi. Daya hambat diukur sebagai berikut: 1, sangat kuat (zona hambat > 2.0 cm); 2, kuat (zona hambat 1.1–2.0 cm); 3, sedang (zona hambat 0.51–1.0 cm); 4, lemah (zona hambat 0–0.5 cm).
Identifikasi Senyawa Metabolit Asal B. cereus 11UJ Menggunakan GC-MS
Ekstrak yang digunakan adalah ekstrak yang diperoleh dari hasil ekstraksi metabolit asal B. cereus 11UJ yang dipisahkan meng-gunakan etil asetat, kemudian dianalisis dengan GC-MS. Identifikasi senyawa meng-gunakan metode pirolisis GC-MS mengikuti Narayana et al. (2008). Contoh yang diambil dimasukkan ke dalam ruang kuarsa dalam unit pirolisis yang kemudian dipanaskan dalam lingkungan bebas oksigen pada suhu yang sudah ditentukan sebelumnya. Campuran
senyawa hasil ekstraksi kemudian dimasukkan dalam kolom GC-MS Shimadzu Type GCMS-QP2010 dengan kondisi GC-MS untuk analisis sebagai berikut: Gas: Helium; Detector: FID; Column: Capiler type phase Rtx-5MS (60 m; 0.25 mm); ID Column temperature: 50 °C; Inlet press (kPa): 100; Column flow (mL min-1): 0.85; Split ratio: 112.3; SPL temperature: 280 °C;MS Interface: 280 °C. Spektrometer massa dioperasikan dalam mode ionisasi elektron pada 70 eV dengan suhu 200 °C. Spektrummassa kemudian dibandingkan dengan basis data Mass Spectrometry.
HASIL
Aktivitas Anticendawan Metabolit B. cereus 11UJ terhadap R. solani dan P. oryzae
Dari proses ekstraksi dan evaporasi diperoleh larutan ekstrak yang lebih pekat dan murni. Hasil uji aktivitas anticendawan terhadap cendawan patogen padi menunjukkan bahwa ekstrak B. cereus 11UJ dapat menekan pertumbuhan P. oryzae (Gambar 1). Produksi metabolit sekunder B. cereus 11UJ dapat mengurangi pertumbuhan miselium P. oryzae. Senyawa metabolit menurunkan panjang miselium dan koloni cendawan sesuai per-lakuan konsentrasi metabolit dengan kisaran daya hambat 0.3–1.4 cm.
Penghambatan terhadap P. oryzae oleh metabolit B. cereus 11UJ lebih efektif di-bandingkan dengan penghambatannya ter-hadap R. solani 7 hari setelah inkubasi. Metabolit bakteri tidak sepenuhnya mematikan P. oryzae, tetapi memiliki efek yang jelas dalam menghambat pertumbuhan miselium cendawan (Tabel 1). Zona hambat yang dihasilkan oleh B. cereus terhadap P. oryzae termasuk dalam kategori daya hambat kuat (1.1–2.0 cm) pada masing-masing perlakuan 250, 500, dan 1000 ppm, sedangkan zona hambat yang dihasilkan oleh R. solani dapat dikategorikan ke dalam zona hambat lemah (< 0.5 cm).
Fraksi Metabolit Sekunder Asal B. cereus 11 UJSenyawa yang terdeteksi melalui GC-
MS, termasuk golongan alkohol (etanol),

J Fitopatol Indones Suryadi et al.
38
asam asetat, nonane, asam palmitat, dan keton (Gambar 2). Ekstrak B. cereus 11UJ yang diuji menunjukkan 30 puncak, tetapi hanya 3 senyawa dengan konsentrasi tertinggi yang terdeteksi dalam ekstrak, yaitu senyawa 9,19-cyclolanostan-3-ol, acetate, (3.beta.)- (CAS) cycloartanyl acetate; 4-(2’,2’-dimethyl – 6’ – methyliden - 1’ - cyclohexyliden) -3 –methyl -2- butanone; dan stigmast-5-en-3-ol, oleat (Gambar 2; Tabel 2).
PEMBAHASAN
Aktivitas anticendawan B. cereus 11UJ yang mudah menguap pada cawan petri menghambat pertumbuhan miselium cendawan
yang berbeda-beda sesuai dengan konsentrasi metabolit. Hasil penelitian menunjukkan bahwa ekstrak B. cereus 11UJ mempunyai aktivitas anticendawan agak lemah terhadap R. solani yang ditandai rendahnya daerah hambatan. Pada perlakuan kontrol negatif tidak menunjukkan adanya zona hambat, berarti bahwa pelarut tidak mempengaruhi aktivitas anticendawan sehingga diduga aktivitas hanya berasal dari metabolit yang diuji.
Ekstrak metabolit B. cereus 11UJ dengan konsentrasi 1000 ppm menghasilkan zona hambat paling besar dibandingkan dengan konsentrasi lainnya. Semakin rendah konsentrasi, semakin rendah pula zona hambat yang dihasilkan. Hal ini sama dengan
Metabolit B. cereus 11UJ (ppm)
Rerata diameter penghambatan (cm)Pyricularia oryzae Rhizoctonia solani
1000 1.40 ± 0.14 0.75 ± 0.04500 1.37 ± 0.04 0.25 ± 0.07250 1.05 ± 0.07 0.17 ± 0.04100 0.26 ± 0.04 0.10 ± 0.01
0 0.00 ± 0.00 0.00 ± 0.00
Tabel 1 Hasil uji aktivitas metabolit Bacillus cereus 11UJ terhadap Pyricularia oryzae dan Rhizoctonia solani
Gambar 1 Bioasai beberapa konsentrasi ekstrak kasar metabolit Bacillus cereus 11UJ (B)terhadap Pyricularia oryzae (P).
P
B B
P P
B
100 ppm 500 ppm250 ppm
B
PP
1000 ppm 0 ppm

J Fitopatol Indones Suryadi et al.
39
pendapat peneliti sebelumnya (Widiyati 2006). Pada penelitian yang dilakukan sebelumnya terhadap senyawa metabolit murni yang diperoleh dari filtrat Pseudomonas brassicacearum YC5420 dilaporkan bahwa besar zona hambat pada cendawan patogen berbeda-beda (Chung et al. 2008). Besarnya zona hambat yang dihasilkan kemungkinan berhubungan dengan aktivitas antimikrob. Dilaporkan oleh Dutta et al. (2013), bahwa faktor-faktor yang mempengaruhi aktivitas antimikrob di antaranya perbedaan pH lingkungan, stabilitas zat aktif, besarnya inokulum, masa inkubasi dan aktivitas metabolik bakteri.
Berdasarkan pada daya hambat yang dihasilkan oleh Bacillus sp. terhadap cendawan R. solani, Yuliar (2008) menyatakan bahwa Bacillus sp. berpotensi sebagai agens pengendali hayati R. solani. Bakteri B. cereus
mampu mensintesis protein yang dapat meningkatkan ketahanan tanaman tomat terhadap cendawan Corynespora casiicola (Romeiro et al. 2010). Kemampuan daya hambat yang dihasilkan oleh spesies Bacillus terhadap bakteri patogen pangan dilaporkan pula oleh Lisboa et al. (2006) yang menyatakan bahwa kultur supernatan B. amyloliquefaciens menunjukkan penghambatan pertumbuhan terhadap beberapa jenis bakteri seperti Listeria monocytogenes, Serratia marcescens dan Pasteurela haemolytica. Penelitian yang dilakukan oleh El-hamshary dan Khattab (2008) melaporkan bakteri B. subtilis dan B. cereus mempunyai aktivitas anticendawan yang tinggi terhadap cendawan patogen Fusarium solani.
Cyclolanostan merupakan salah satu metabolit sekunder yang dihasilkan oleh B. cereus 11UJ dengan kadar tertinggi.
Gambar 2 Kromatogram ekstrak metabolit Bacillus cereus 11UJ.
Senyawa Kadar (%)
Waktu retensi (menit)
Berat molekul(g mol-1)
9,19-cyclolanostan-3-ol, acetate, (3.beta.)- (CAS) cycloartanyl acetate 13.14 32.893 470
4-(2’,2’-dimethyl-6’-methyliden-1’-cyclohexyliden)-3-methyl-2-butanone 9.72 31.273 206
stigmast-5-en-3-ol, oleat 9.09 33.487 679
Tabel 2 Tiga senyawa metabolit sekunder dengan kadar tertinggi dalam ekstrak Bacillus cereus 11UJ
TIC*1.00
10.0 30.020.0 49.0 min40.0
12 904 231

J Fitopatol Indones Suryadi et al.
40
Lanostan merupakan turunan dari senyawa triterpena (steroid) (Escurriol et al. 2009). Sejumlah terpen atau terpenoid dilaporkan aktif melawan cendawan dan serangga. Beberapa senyawa turunan triterpena telah dijadikan sebagai anticendawan, tetapi mekanisme aksi terpen tidak sepenuhnya diketahui, namun diduga menyebabkan gangguan membran oleh sifat lipofil (Ghosh et al. 2013). Dimethyl-6’-methyliden-1’-cyclohexyliden)-3-methyl-2-butanone merupakan kelompok monoterpenoid (flavonoid) dari jalur asetat mevalonat dan juga bersifat anticendawan. Stigmast-5-en-3-ol, oleat merupakan senyawa turunan sterol dan metabolit sekunder yang dihasilkan dari ekstrak bakteri B. cereus. Senyawa ini digolongkan dalam kelompok fitoaleksin, oleh karena itu stigmasterol dapat bersifat sebagai anticendawan. Senyawa stigmasterol dapat bersifat anticendawan terhadap Malazesia furfureus, Candida albicans dan Aspergillus niger (Ridhay et al. (2012).
Senyawa n-decanal mudah menguap dari B. amyloqueafaciens NJN-6 dilaporkan dapat menghambat Fusarium oxysporum f. sp. cubense (Yuan et al. 2012), sementara alkil alkohol dapat meningkatkan populasi bakteri dan 2,3-butadienol memacu pertumbuhan tanaman (Dutta et al. 2013). Jika besarnya konsentrasi stigmast-5-en-3-ol, oleat dalam ekstrak yang dihasilkan oleh B. cereus terakumulasi kemungkinan besar dapat menyebabkan penumpukan sterol dalam cendawan. Pernyataan tersebut diungkapkan oleh Lazzarino et al. (2011), bahwa akumulasi sterol yang mengalami metilasi menyebabkan terjadinya perubahan sejumlah fungsi sel yang berhubungan dengan permeabilitas membran.
Senyawa benzena dan benzothiazoles fenol dan 2,3,6-trimetil-fenol asal metabolitsekunder B. amyloliqueafaciens NJN-6 dilaporkan mempunyai aktivitas anticendawan terhadap F. oxysporum. Secara in vitro senyawa alkohol aldehida, ester, eter, dan naftil pada konsentrasi 200 µL hampir sepenuhnya menghambat F. oxysporum, sedangkan keton diproduksi pada tingkat tinggi (Yuan
et al. 2012). Pada penelitian ini senyawa keton (4-(2’,2’-dimethyl-6’-methyliden-1’-cyclohexyliden)-3-methyl-2-butanone) yang diproduksi oleh B. cereus 11UJ kadarnya juga ditemukan cukup tinggi. Sementara berdasarkan analisis menggunakan GC-MS yang dilakukan oleh Dutta et al. (2013) terhadap B. cereus, juga diperoleh 3 jenis senyawa organik. Tiga jenis senyawa tersebut adalah senyawa golongan sulfida (disulfida dan dimetil), golongan aldehida (nonanal atau decanal), dan satu senyawa asam (1,2 benzenedicarboxylic acid).
Dari hasil analisis GC-MS, diketahui sterol memiliki puncak paling tinggi dan diduga menunjukkan efek anticendawan yang kuat terhadap cendawan patogen. Hasil percobaan ini menyarankan bahwa mekanisme pe-ngendalian hayati yang dihasilkan oleh B. cereus 11UJ terhadap P. oryzae dan R. solani diduga disebabkan senyawa metabolit sekunder tersebut. Kegiatan uji antagonis berupa penghambatan pertumbuhan miselium kedua patogen hasilnya berguna untuk memahami tentang mekanisme biokontrol B. cereus 11UJ terhadap P. oryzae dan R. solani. Perlu dilakukan penelitian lebih lanjut untuk menentukan karakteristik masing-masing senyawa utama anticendawan yang dihasilkan oleh bakteri B. cereus 11UJ terhadap R. solani dan P. oryzae melalui analisis lainnya seperti kromatografi lapis tipis (KLT), bioautografi dan nuclear magnetic resonance (NMR).
Metabolit sekunder yang dihasilkan dari ekstrak bakteri B. cereus 11UJ dapat menghambat pertumbuhan cendawan R. solani dan P. oryzae yang cukup baik pada konsentrasi 1000 ppm, serta dapat diketahui bahwa aktivitas anticendawan yang dihasilkan lebih efektif menghambat pertumbuhan P. oryzae. Hasil analisis senyawa menggunakan GC-MS menunjukkan 3 senyawa dengan kadar tinggi, yaitu 9,19-cyclolanostan-3-ol, acetate, (3.beta.)-(CAS) cycloartanyl acetate (13.14%); 4-(2’,2’-dimethyl-6’-methyliden-1’-cyclohexyliden)-3-methyl-2-butanone (9.72%); dan stigmast-5-en-3-ol, oleat (9.09%).

J Fitopatol Indones Suryadi et al.
41
UCAPAN TERIMA KASIH
Penelitian ini didanai oleh DIPA APBN 2013-BB Biogen atas nama Sutoro dengan No kode Projek:1798.009.011. Penulis mengucapkan terima kasih kepada teknisi Lab Mikrobiologi BB Biogen dan Siti Sadiah yang telah membantu kelancaran penelitian ini. Penulis juga mengucapkan penghargaan kepada Ika Mariska atas diskusi, koreksi, dan saran yang bermanfaat pada penafsiran analisis metabolit sekunder.
DAFTAR PUSTAKA
Chung BS, Aslam Z, Kim SW, Kim GG, Kang HS, Ahn JW, Chung YR. 2008. A bacterial endophyte Pseudomonas brassicacearum YC5480 isolated from the root of Artemisia sp producing antifungal and phytotoxic compounds. Plant Pathol J. 24(4):461–468. DOI: http://dx.doi.org/10.5423/PPJ.2008.24.4.461.
Dutta S, Rani TS, Podile AR.2013. Root exudate-induced alterations in Bacillus cereus cell wall contribute to root colonization and plant growth promotion. J PlosOne. 8(10):e78369. DOI: http://dx.doi.org/10.1371/journal.pone.0078369.
El-hamshary OIM, Khattab AA. 2008. Evaluation of antimicrobial activity of B. subtilis and B. cereus and their fusant against Fusarium solani. Res J Cell Mol Biol. 2(2):24–29.
Escurriol V, Cofan M, Serra M, Bullo M, Basora J, Salvado JS, Corella D, Zazpe I, Gonzalez MAM, Gutierrez VR, Estruch R, Ros E. 2009. Serum sterol responses to increasing plant sterol intake from natural foods in the Mediterranean diet. Eur J Nutr. DOI: http://dx.doi.org/10.1007/s00394-009-0024-z.
Farag MA, Ryu CM, Sumner LW, Pare PW. 2006. GC-MS SPME profiling of rhizobacterial volatiles reveals prospective inducers of growth promotion and induced systemic resistance in plants. Phytochemistry. 67:2262–2268.
D O I : h t t p : / / d x . d o i . o rg / 1 0 . 1 0 1 6 / j .phytochem.2006.07.021.
Fiddaman PJ, Rossall S. 1994. Effect of substrate on the production of antifungal volatiles from Bacillus subtilis. J Appl Bacteriol. 76:395–405. DOI: http://dx.doi.org/10.1111/j.1365-2672.1994.tb01646.x.
Ghosh P, Mandal A, Rasul MG. 2013. A new bioactive ursane-type triterpenoid from Croton bonplandianum Bail. J Chem Sci. 125(2):359–364. DOI: http://dx.doi.org/10.1007/s12039-013-0387-9.
Gomathi S, Ambikapathy V, Panneerselvam A. 2013. Separation of bioactive compounds from T. viride isolated from chilli field soil by using gas chromatography-mass spectrum, FTIR and UV. J Antimocrobials Photon. 128:141–146.
Jing L, Qian Y. 2007. Purification and properties of antifungal protein produced by Bacillus subtilis B29. Di dalam: The 5th International Symposium on Biocontrol and Biotechnology; 2007 Nov 1–3; Khon Kaen University, Nong Khai, Thailand. Nong Khai (TH): [King Mongkut’s Institute of Technology Ladkrabang and Khon Kaen University]. 37. O-27.
Kang BR. 2012. Biocontrol of tomato Fusarium wilt by a novel genotype of 2,4-diacetyl-phloroglucinol-producing Pseudomonas sp. NJ 134. Plant Pathol J. 28(1):93–100. DOI:http://dx.doi.org/10.5423/PPJ.NT.10.2011.0202.
Lazzarino G, Amorini AM, Di Pietro V, Tavazzi B. 2011. HPLC analysis for the clinical-biochemical diagnosis of inborn errors of metabolism of purines and pyrimidines.Methods Mol Biol. 708:99–117. DOI: http://dx.doi.org/10.1007/978-1-61737-985-7_5.
Lisboa MP, Bonatto D, Bizani D, Henriques JAP, Brandelli A. 2006. Characterization of a bacteriocin-like substance produced by B. amyloliquefaciens isolated from the Brazilian Atlantic forest. Inter Microbiol. 9:111–118.
Muskhazli M, Leong PP, Nor Azwady AA, Nalisha I, Nor Farizan T, Siraj SS, Faridah

J Fitopatol Indones Suryadi et al.
42
QZ. 2007. Studies on the activity of soluble non-volatiles antifungal secreted by B. subtilis. Malays Appl Biol. 3(2):7–13.
Narayana KJP, Prabhakar P,Vijayalakshmi M,Venkateswarlu Y, Krishna P. 2008. Study on bioactive compounds from Streptomyces sp. ANU 277. Polish J Microbiol. 57(1):35–39.
Płaza GA, Król E, Páociniczak MP, Seget ZP, Brigmon LR. 2012. Study of antifungal activity of Bacillus species cultured on agro-industrial wastes. Acta Sci Pol Hortorum Cultus. 11(5):169–182.
Ridhay A, Noor A, Soekamto NH, Harlim T. 2012. A stigmasterol glycoside from the root wood of Melochia umbellata (Houtt) stapf var. degrabrata K. Indonesian J Chem.12 (1):100–103.
Romeiro RS, Filho FL, Macagnan D, Garcia FAO, Silva HSA. 2010. Evidence that the biocontrol agent B. cereus synthesizes protein that can elicit increased resistance of tomato leaves to Corynespora casiicola. Trop Plant Pathol. 35(1):11–15.
Sadrati N, Daoud H, Zerroug A, Dahamna S, Bouharati S. 2013. Screening of antimicrobial and antioxidant secondary metabolites from endophytic fungi isolated from wheat (Triticum durum). J Plant Protec Res. 53(2):128–136.
Soria S, Alonso R, Battucci L. 2012. Endophytic bacteria from Pinus taeda L as biocontrol agents of Fusarium circinatum
Nirenberg & O’Donnell. Chilean J Agric Res. 72(2):281–284.
Suryadi Y, Priyatno TP, Susilowati DN, Samudra IM, Yudistira N, Purwakusumah ED. 2013. Isolasi dan karakterisasi kitinase asal B. cereus 11UJ. J Biol Indones. 9(1):51–62.
Wen CY, Yin ZG, Wang KX, Chen JG, Shen SS. 2011. Purification and structural analysis of surfactin produced by endophytic B. subtilis EB 505 and its antagonistic activity against R. cerealis. Plant Pathol J. 27(4):342–348. DOI: http://dx.doi.org/10.5423/PPJ.2011.27.4.342.
Widiyati E. 2006 Penentuan adanya senyawa triterpenoid dan uji aktivitas biologis pada beberapa spesies tanaman obat tradisional masyarakat pedesaan Bengkulu. J Gradien. 2(1):116–122.
Yuan J, Raza W, Shen Q, Huang Q. 2012. Antifungal activity of Bacillus amyloliquefaciens NJN-6 volatile compounds against Fusarium oxysporum f. sp. cubense. Appl Environ Microbiol. 78(16):5942–5944. DOI: http://dx.doi.org/10.1128/AEM.01357-12.
Yuliar. 2008. Skrining bioantagonistik bakteri untuk agen biokontrol Rhizoctonia solani dan kemampuannya dalam menghasilkan surfaktin. Biodiversitas. 9:83–86. DOI: ht tp: / /dx.doi .org/10.13057/biodiv/d090201.

Volume 11, Nomor 2, April 2015Halaman 43–50
DOI: 10.14692/jfi.11.2.43ISSN: 0215-7950
*Alamat penulis korespondensi: Departemen Proteksi Tanaman, Fakultas Pertanian, Institut Pertanian Bogor,Jalan Kamper, Kampus Darmaga IPB, Bogor 16680.Tel: 0251-8629364, Faks: 0251-8629362; surel: [email protected]
43
Identifikasi Polerovirus Penyebab Klorosis pada Cabai Asal Bali, Indonesia
Identification of Polerovirus Causing Chlorosis on Chilipepper in Bali, Indonesia
Rita Kurnia Apindiati, Gede Suastika*, Kikin Hamzah Mutaqin Institut Pertanian Bogor, Bogor 16680
ABSTRAK
Berdasarkan hasil survei yang dilakukan pada pertanaman cabai di Desa Kertha, Kecamatan Payangan, Kabupaten Gianyar, Provinsi Bali, ditemukan banyak tanaman cabai dengan gejala klorosis yang disebabkan oleh virus. Tanaman sakit menunjukkan gejala kekuningan pada lamina daun, tetapi tulang daun tetap berwarna hijau. Usaha identifikasi difokuskan pada Pepper vein yellows virus (PeVYV) melalui penularan menggunakan Aphis nasturtii, pengamatan bentuk partikel virus, deteksi RNA total dengan reverse transcription-polymerase chain reaction (RT-PCR), dan perunutan sikuen nukleotida. Hasil diagnosis menunjukkan bahwa penyakit dapat ditularkan dari tanaman cabai sakit ke tanaman cabai sehat menggunakan A. nasturtii. Partikel virus berbentuk heksagonal dengan diameter berukuran ~30 nm. RT-PCR RNA total dari tanaman sakit bergejala berat, sedang, dan ringan menggunakan pasangan primer spesifik gen protein selubung PeVYV berhasil mengamplifikasi pita DNA berukuran ~650 pb. Sikuen gen protein selubung PeVYV isolat asal Bali dari intensitas gejala klorosis yang berbeda memiliki homologi tertinggi dengan PeVYV isolat Taiwan berkisar 95.1–96.6%. Dengan demikian, PeVYV dikonfirmasi sebagai penyebab klorosis pada pertanaman cabai di Bali, Indonesia.
Kata kunci: Aphis nasturtii, Pepper vein yellows virus (PeVYV), protein selubung
ABSTRACT
Based on the results of a survey conducted on chilipepper plants in Kertha Village, Payangan Subdistrict, Gianyar District, Bali Province, many chilipepper plants with chlorosis symptoms caused by the virus was observed. Infected plants showed yellowing of the lamina, but the veins remain green. Identification was focused on the Pepper vein yellows virus (PeVYV) through transmission using Aphis nasturtii, observation on the morphology of virus particles, total RNA detection by reverse transcription-polymerase chain reaction (RT-PCR), and nucleotide sequence. The results showed that the disease can be transmitted from infected plant to healthy plant using A. nasturtii. Viral particle has hexagonal shape with the diameter ~30 nm. RT-PCR of total RNA from severe, moderate, and mild symptomatic infected plants, using specific primer sets for coat protein gene of PeVYV successfully amplified DNA with size of ~650 bp. Coat protein gene sequence of PeVYV isolates Bali, Indonesia showing different chlorosis intensity had the highest homology with PeVYV isolates from Taiwan 95.1–96.6%. Based on the data, PeVYV has been confirmed as the cause of chlorosis in chilipepper plants in Bali, Indonesia.
Key words: Aphis nasturtii, coat protein, Pepper vein yellows virus (PeVYV)

J Fitopatol Indones Apindiati et al.
44
PENDAHULUAN
Cabai (Capsicum annum) merupakan salah satu komoditas hortikultura yang memiliki nilai ekonomi penting di Indonesia. Berdasarkan data Badan Pusat Statistik produktivitas cabai besar berfluktuasi dari tahun 2009 sampai 2013 berturut-turut: 0.79, 0.81, 0.89, 0.95, dan 1.01 juta ton (BPS 2014a); sedangkan produksi cabai rawit: 0.591, 0.522, 0.594, 0.702, dan 0.714 juta ton (BPS 2014b). Hal tersebut dikarenakan adanya faktor pembatas produksi, yaitu teknik budi daya yang tidak intensif serta gangguan hama dan penyakit tanaman. Penyakit pada tanaman cabai di Indonesia disebabkan oleh beberapa virus penting di antaranya Geminivirus, Cucumber mosaic virus (CMV), Chili veinal mottle virus (ChiVMV), serta Pepper vein yellows virus (PeVYV).
Pada tahun 1981 tanaman paprika di Kitanakagusuku, Okinawa, Jepang menunjuk-kan gejala klorosis dengan tulang daun menguning dan daun menggulung. Gejala yang sama ditemukan pada area yang berbeda, yaitu di Okinawa, Ishigaki, dan Kepulauan Miyako yang diduga disebabkan oleh infeksi virus (Yonaha et al. 1995). Gejala ini awalnya diduga disebabkan oleh defisiensi magnesium, namun terdapat partikel virus yang diisolasi dari tanaman. Virus ini menyerupai partikel Luteovirus. Virus bereplikasi di jaringan floem dan dapat ditularkan melalui penyambungan dan vektor Aphis gossypii. Selanjutnya, virus ini diberi nama Pepper vein yellows virus (PeVYV) (Murakami et al. 2011). PeVYV (Luteoviridae; Polerovirus) dilaporkan me-nyebabkan penyakit klorosis. Identifikasi virus sebagai penyebab penyakit merupakan faktor kunci yang menentukan keberhasilan pengendalian di lapangan.
Pada bulan September 2011 di Desa Kertha, Kecamatan Payangan, Kabupaten Gianyar, Provinsi Bali ditemukan banyak tanaman cabai yang menunjukkan gejala klorosis, menguning di antara tulang daun dengan tulang daun dan jaringan sekitarnya tetap hijau sehingga tampak menyirip
(Suastika et al. 2012). Polerovirus ditularkan oleh kutudaun sebagai vektor (Gray dan Gildow 2003). Oleh karena itu, masih terdapat keraguan penyebab gejala klorosis ini maka perlu dilakukan diagnosis baik secara biologi dan molekul untuk mengonfirmasi keberadaan virus tersebut.
BAHAN DAN METODE
Pengambilan Tanaman SakitPengambilan tanaman sakit berasal
dari Desa Kertha, Kecamatan Payangan, Kabupaten Gianyar, Provinsi Bali, Indonesia. Tanaman cabai bergejala klorosis diambil dari lapangan dan dikategorikan menjadi 3 kelompok berdasarkan variasi gejala berat, sedang, dan ringan.
Penularan
Penularan dilakukan menggunakan kutudaun (Aphis sp.) sebagai vektor untuk menularkan Polerovirus melalui daun tanaman cabai. Imago kutudaun yang tidak bersayap dipuasakan selama 120 menit. Sebanyak 50 ekor kutudaun tersebut diletakkan pada tanaman cabai yang terinfeksi Polerovirus dengan periode makan akuisisi selama 15 menit, kemudian dipindahkan meng-gunakan kuas ke tanaman cabai sehat berumur 4 minggu sebanyak 5 ekor setiap tanaman dengan periode inokulasi 24 jam. Kutudaun tersebut dimatikan dan tanaman dipelihara di dalam kurungan kedap serangga. Pengamatan selama 14 hari dan dilakukan identifikasi terhadap spesies kutudaun menggunakan kunci identifikasi Blackman dan Eastop (2000).
Mikroskop ElektronDaun cabai bergejala dicacah halus
kemudian cairannya diteteskan ke atas grid secara negative staining menggunakan glutaraldehyde dengan 25% aqueous solution, (0.1 M Cacodylate Buffer pH 7.4 ditambah 3% Sukrosa). Spesimen tersebut diwarnai uranyl acetate dan lead citrate, selanjutnya diamati menggunakan mikroskop elektron (JOEL JEM 1010, 80 kV).

J Fitopatol Indones Apindiati et al.
45
Isolasi RNA Total dari Tanaman SakitIsolasi RNA total menggunakan 0.1 g daun
cabai bergejala menggunakan Xprep Plant RNA Mini Kit (Phile Korea) sesuai protokol yang dianjurkan.
Sintesis cDNAcDNA disintesis dengan reaktan RT-PCR
yang terdiri atas RNA total 2 µL, 10 mM dNTP(Thermo Scientific, US) 0.5 µL, 10 mM oligo d(T) (Thermo Scientific, US) 0.75 µL, bufer RT 2 µL (Thermo Scientific, US), 0.35 µL DTT (Thermo Scientific, US), RNase inhibitor 0.35 µL (Ribolock-Thermo Scientific, US), MMuLV reverse transcriptase 0.35 µL (Revertaid-Thermo Scientific, US), air bebas sampai volume total 10 µL (Thermo Scientific, US). Reaksi RT dilakukan pada suhu 42 °C selama 60 menit dan dilanjutkan inaktivasi enzim RT pada suhu 95 °C selama 5 menit.
Amplifikasi DNA
Reaktan PCR dilakukan dengan volumetotal 25 µL, terdiri atas Go Tag Green Master Mix 2x 12.5 µL (Promega, US), air bebas nuklease 6.5 µL, amplifikasi coat protein dilakukan dengan meng-gunakan sepasang primer spesifik PeVYV. Primer forward F-BamHII (5’-AATTAAGGATCCCAATACGGGAGGGG TTAGGAGAAAT-3’) dan primer reverse R-PstI (3’-AATTAACTGCAGTTTCGGGT TGTGCAATTGCACAGTA-5’) masing-masing 1 µL. Reaksi PCR dilakukan dengan Perkin Elmer 480 Thermocycler (Applied Biosystem, US). Program amplifikasi yang digunakan ialah denaturasi inisiasi pada 94 °C selama 5 menit, kemudian dilanjutkan 35 siklus yang terdiri atas denaturasi 94 °C selama 30 detik, penempelan primer pada suhu 50 °C selama 1 menit, pemanjangan pada suhu 72 °C selama 1 menit, dan diikuti pemanjangan akhir pada suhu 72 °C selama 10 menit.
Visualisasi DNAVisualisasi fragmen DNA hasil amplifikasi
dielektroforesis menggunakan gel agarosa 1% dalam bufer Tris-borate (TBE) 0.5X dengan tegangan 100 volt selama
30 menit. Pengamatan dilakukan dengan UV transiluminator setelah direndam dalam larutan etidium bromida 2% selama 15 menit.
Perunutan NukleotidaPerunutan nukleotida dilakukan di First
Base, Malaysia untuk merunut hasil amplifikasi gen protein terselubung (CP) Polerovirus. Sikuen gen protein selubung kemudian di-analisis dengan program BLAST (www.ncbi.com). Homologi gen CP Polerovirus terhadap anggota Polerovirus lainnya yang terdeposit dalam Genbank dianalisis menggunakan program ClustalW BioEdit. Pohon filogenetika berdasarkan runutan nukleotida gen CP dibuat menggunakan program perangkat lunak molecular evolutionary genetic analisis (MEGA 5.1) menggunakan neighbour joining dengan bootstrap sebanyak 1000 kali.
HASIL
Gejala PenyakitPenyakit klorosis pada tanaman cabai
menunjukkan variasi gejala dari gejala ringan, sedang, dan berat. Gejala ringan berupa adanya klorosis pada daun cabai (Gambar 1a). Gejala sedang berupa adanya klorosis dengan menguningnya lamina daun (Gambar 1b), sedangkan gejala berat berupa adanya klorosis dengan menguningnya lamina daun disertai dengan penebalan daun (Gambar 1c).
PenularanPenularan menggunakan kutudaun
(Gambar 2) yang diinokulasikan pada daun cabai sehat berhasil menularkan Polerovirus yang menyebabkan gejala klorosis pada semua tanaman uji. Keberhasilan penularan virus dikonfirmasi dengan RT-PCR yang me-nunjukkan Polerovirus positif teramplifikasi DNA berukuran ~650 pb pada semua tanaman uji hasil penularan (Gambar 3).
Bentuk PartikelPartikel Polerovirus diamati dari tanaman
sakit berbentuk heksagonal dengan diameter partikel berukuran ~30 nm (Gambar 4).

J Fitopatol Indones Apindiati et al.
46
Gambar 2 Kutudaun Aphis nasturtii sebagai vektor PeVYV pada tanaman cabai, dengan ciri: 1, terminal proses pada antena lebih panjang daripada pangkal antena; 2, antena tubercle tidak berkembang; 3, striduratory apparatus tidak ada; 4, cauda berbentuk lidah dan berwarna pucat; 5, siphunculli berwarna pucat dan ujungnya berwarna gelap dan; 6, rambut pada femur panjang.
Gambar 1 Gejala Polerovirus pada daun cabai asal Payangan, Bali, Indonesia, diklasifikasikan menjadi tiga kelompok gejala: a, ringan; b, sedang dan; c, berat.
a b c
Gambar 3 Hasil amplifikasi DNA dari daun tanaman cabai bergejala klorosis dari hasil penularan Aphis nasturtii menggunakan pasangan primer spesifik PeVYV. K(-), kontrol sehat; K(+), kontrol positif dari tanaman sakit; M, marker 1 kpb DNA ladder (Thermo Scientific, US).
~ 650 pb

J Fitopatol Indones Apindiati et al.
47
RT-PCRRT-PCR menggunakan primer spesifik
PeVYV berhasil mengamplifikasi fragmen DNA ~650 pb pada sampel dengan gejala berat, sedang, dan ringan yang menunjukkan gejala positif seperti halnya pada kontrol positif dari daun tanaman cabai asal Payangan, Bali, Indonesia (Gambar 5).
Homologi Genetika PolerovirusSikuen gen CP Polerovirus isolat Payangan,
Bali yang bergejala ringan, sedang, dan berat apabila dibandingkan di antara ketiganya memiliki tingkat homologi di atas 97% dan memiliki kekerabatan yang dekat dengan PeVYV isolat Taiwan, dengan homologi di atas 95%. Selain itu, PeVYV sangat mirip dengan PYLCV isolat Israel seperti yang dikemukakan oleh Dombrovsky et al. (2010). Semua isolat PeVYV yang dianalisis tersebut memiliki tingkat homologi di atas 89% dengan PYLCV isolat Israel (Tabel 1). Analisis filogenetika pada kladogram menunjukkan dan membuktikan kejelasan hubungan kekerabatan Polerovirus. Polerovirus isolat Bali, Indonesia baik yang bergejala klorosis ringan, sedang, maupun berat termasuk ke dalam spesies Pepper vein yellows virus (PeVYV) (Gambar 6).
PEMBAHASAN
Penyakit klorosis dengan gejala berupa menguningnya tulang daun yang ditemukan pada daun cabai di Bali, Indonesia tersebut sama dengan yang telah dilaporkan oleh Suastika et al. (2012) disebabkan oleh Polerovirus. Namun, gejala tersebut tidak memperlihatkan pemendekan internoda atau daun yang menggulung.
Berdasarkan Yonaha et al. (1995) tanaman cabai yang diinokulasi mekanis menggunakan cairan perasan dari daun tanaman cabai sakit tidak menunjukkan adanya gejala klorosis. Namun, setelah menggunakan penyambungan dan vektor kutudaun menunjukkan adanya gejala klorosis dengan menguningnya tulang daun secara jelas pada bagian atas daun dan setelah itu semua daun berkembang menjadi menggulung dan menguning. Gejala ini biasanya disertai dengan bentuk buah mengalami malformasi. Penularan ini dilakukan secara sirkulatif oleh kutudaun. Gray dan Gildow (2003) menyatakan bahwa virus tertelan dan bergerak pada saluran makanan melalui foregut dan terakumulasi dalam midgut atau hindgut kemudian virus terakumulasi dalam haemocoel
Gambar 5 Hasil amplifikasi DNA sampel daun tanaman cabai bergejala klorosis menggunakan pasangan primer spesifik PeVYV. B, gejala berat; S, gejala sedang; R, gejala ringan; K(-), kontrol sehat; K(+), kontrol positif dari tanaman sakit; M, penanda DNA 1 kb (Thermo).
~650 pb
Gambar 4 Partikel virus melalui pengamatan dengan mikroskop elektron. Diameter partikel virus berukuran ~30 nm.
± 100 nm

J Fitopatol IndonesA
pindiati et al.
48
Virus Asal Negara No. Aksesi 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15
Polerovirus Bali Idn-Bb - IDPolerovirus Bali Idn-Bs - 97.4 IDPolerovirus Bali Idn-Br - 97.5 97.0 IDPeVYV Jepang AB594828 96.4 95.4 94.9 IDPeVYV Taiwan JX427542 96.6 95.4 95.1 98.7 IDPeVYV Thailand JX4275p40 95.4 94.9 94.1 97.8 97.8 IDPeVYV Filipina JX427537 95.4 94.9 94.3 98.2 98.5 97.7 IDPeVYV India JX427531 95.1 94.6 93.8 97.5 97.5 99.6 97.4 IDPeVYV Mali JX427535 95.1 94.3 93.6 97.2 97.5 98.3 97.0 98.0 IDPeVYV Turki FN600344 94.1 94.0 93.2 96.9 97.2 97.2 97.7 96.9 96.2 IDCABYV Jepang EU000535 61.9 62.2 61.4 63.0 63.5 63.0 63.2 63.0 62.4 63.3 IDPABYV Mali KF4277698 61.4 61.4 60.8 62.7 63.3 62.5 63.2 62.4 62.2 64.0 85.8 IDPYLCV Israel HM439608 90.3 89.6 89.0 92.7 92.8 93.0 92.5 92.7 92.2 92.7 61.6 62.2 IDTVDV China EF529624 89.9 89.0 88.6 90.7 91.2 90.6 91.5 90.3 91.4 91.2 61.2 62.0 88.2 IDGeminivirus USA AF063139 35.2 34.7 34.7 35.7 35.5 35.7 35.5 36.0 35.9 35.7 34.1 32.6 34.6 34.4 ID
Tabel 1 Tingkat kesamaan sikuen nukleotida sebagian gen protein selubung (CP) Polerovirus isolat cabai Payangan, Bali, Indonesia dengan isolat lainnya yang telah dilaporkan menggunakan program Bioedit V.7.0.5

J Fitopatol Indones Apindiati et al.
49
selama berminggu-minggu. Virus kemudian diinjeksikan ke dalam tanaman melalui saluran air liur ketika kutudaun tersebut makan pada tanaman. Spesies kutudaun yang berhasil menularkan Polerovirus ialah Aphis nasturtii, berbeda dengan yang telah dilaporkan oleh Yonaha et al. (1995) yang menyebutkan bahwa Aphis gosypii dan Myzus persicae yang mampu menularkan virus Polerovirus. Apabila dilihat secara makroskopi ciri morfologi antara A. nasturtii dan A. gosypii sama, tetapi ketika diidentifikasi secara mikroskopi terdapat perbedaan, yaitu siphunculli yang berwarna pucat dengan ujung yang berwarna gelap pada A. nasturtii.
Deteksi menggunakan metode RT-PCR yang dilakukan menunjukkan virus yang menginfeksi cabai asal Bali, Indonesia ialah Pepper vein yellows virus (PeVYV) dan
memiliki tingkat homologi paling tinggi dengan isolat PeVYV asal Taiwan (96.6%). Menurut Fauquet et al. (2005) apabila terdapat persamaan sikuen nukleotida dari gen protein selubung antara satu virus dengan virus yang lain dengan nilai lebih dari 90%, virus tersebut merupakan spesies virus yang sama. Analisis filogenetika pada kladogram menunjukkan kejelasan hubungan kekerabatan Polerovirus. Polerovirus isolat Bali, Indonesia baik yang bergejala klorosis ringan, sedang, maupun berat termasuk ke dalam spesies Pepper vein yellows virus (PeVYV) serta memiliki kekerabatan yang dekat dengan PYLCV asal Israel dan TVDV asal Cina, tetapi berada paling jauh kekerabatannya dengan CABYV asal Jepang dan PABYV asal Mali. Perbedaan hubungan kekerabatan tersebut kemungkinan disebabkan oleh perbedaan inang masing-
Gambar 6 Filogenetika kekerabatan Polerovirus isolat cabai Bali, Indonesia (3 isolat dengan gejala berbeda, yaitu ringan, sedang, dan berat) dengan beberapa isolat virus lain yang dilaporkan di GenBank berdasarkan sebagian sikuen selubung protein. PeVYV_Jepang (AB594828), PeVYV_Taiwan (JX427542), PeVYV_Thailand (JX427540), PeVYV_Filipina (JX427537), PeVYV_India (JX427531), PeVYV_Mali (JX427535), PeVYV_Turki (FN600344), CABYV_Jepang (EU000535), PABYV_Mali (KF4277698), PYLCV_Israel (HM439608), TVDV_China (EF529624), Geminivirus_USA (AF063139). Analisis dilakukan menggunakan metode neighbour joining tree pada program MEGA 5.1 dengan bootstrap 1000.
Polerovirus_sedang
Polerovirus_berat
Polerovirus_ringan
PeVYV_Jepang_AB594828
PeVYV_Taiwan_JX427542
PeVYV_Thailand_JX427540
PeVYV_Filipina_JX427537
PeVYV_India_JX427531
PeVYV_Mali_JX427535
PeVYV_Turki_FN600344
CABYV_Jepang_EU000535PABYV_Mali_KF4277698
PYLCV_Israel_HM439608
TVDV_China_EF529624
Geminivirus_USA_AF063139

J Fitopatol Indones Apindiati et al.
50
masing virus dari genus Polerovirus, famili Luteoviridae. Geminivirus asal USA digunakan sebagai outgrup dalam analisis ini.
Berdasarkan gejala ringan, sedang, dan berat yang dianalisis secara biologi untuk melihat morfologi dari Polerovirus isolat Bali, Indonesia tersebut, penularan menggunakan A. nasturtii, deteksi menggunakan RT-PCR, analisis similaritas nilai penyejajaran dari sikuen nukleotida PeVYV yang diperoleh dari sikuen selubung protein maka dikonfirmasi bahwa Polerovirus isolat Bali, Indonesia merupakan PeVYV.
UCAPAN TERIMA KASIH
Penelitian ini dibiayai oleh kerja sama Fakultas Pertanian, Institut Pertanian Bogor dengan Utsunomiya University Jepang.
DAFTAR PUSTAKA
Blackman RL, Eastop VF. 2000. Aphids on the World’s Crops: An Identification and Information Guide. Ed ke-2. New Jersey (US): Wiley.
[BPS] Badan Pusat Statistik. 2014a. Produksi cabe besar menurut provinsi, 2009-2013. Berita resmi statistik BPS. http://www.pertanian.go.id/ATAP_2013_IP/PR_Cabe_Besar_%20(ATAP)%20.pdf”http://www.pertanian.go.id/ATAP_2013_IP/PR_Cabe_Besar_ (ATAP) .pdf. [diunduh 2015 Jan 20].
[BPS] Badan Pusat Statistik. 2014b. Produksi cabe rawit menurut provinsi, 2009-2013. Berita resmi statistik BPShttp://www.pertanian.go.id/ATAP_2013_IP/PR_Cabe_Rawit_%20(ATAP).%20pdf”http://
www.pertanian.go.id/ ATAP_2013_IP/PR_Cabe_Rawit_ (ATAP). pdf. [diunduh 2015 Jan 20].
Dombrovsky A, Glanz E, Perlsman M, Lachman O, Antignus Y. 2010. Characterization of Pepper yellow leaf curl virus, a tentative new Polerovirus species causing a yellowing disease of pepper. Phytoparasitica. 38(5):477–486. DOI: http://dx.doi.org/10.1007/s12600-010-0120-x.
Fauquet CM, Mayo MA, Maniloff J, Desselberger U, Ball LA, editor. 2005. Virus Taxonomy: Eighth Report of the International Committee on Taxonomy of Viruses. San Diego (US): Virol Div Int Union of Microb.
Gray S, Gildow FE. 2003. Luteovirus-aphid interactions. Annu Rev Phytopathol. 41:539–66. DOI: http://dx.doi.org/10.1146/annurev.phyto.41.012203.105815.
Murakami R, Nakashima N, Hinomoto N, Kawano S, Toyosato T. 2011. The genome sequence of Pepper vein yellows virus (family Luteoviridae, genus Polerovirus). Arch Virol. 156:921–923. DOI: http://dx.doi.org/10.1007/s00705-011-0956-5.
Suastika G, Hartono S, Nyana IDN, Natsuaki T. 2012. Laporan pertama tentang infeksi Polerovirus pada tanaman cabai di daerah Bali, Indonesia. J Fitopatol Indones. 8(5): 151–154.
Yonaha T, Toyosato T, Kawano S, Osaki T. 1995. Pepper vein yellows virus, a Novel Luteovirus from Bell Pepper Plants in Japan. Ann Phytopathol Soc Jpn. 61:178–184. DOI: http://dx.doi.org/10.3186/jjphytopath.61.178.

Volume 11, Nomor 2, April 2015Halaman 51–58
DOI: 10.14692/jfi.11.2.51ISSN: 0215-7950
Formulasi Bakteri Endofit Akar Kedelai untuk Pengendalian Pustul Bakteri
Formulation of Bacterial Endophytes from Soybean Root to Control Bacterial Pustule
Trimurti Habazar*, Zurai Resti, Yulmira Yanti, Sutoyo, Imelda Universitas Andalas, Padang 25163
ABSTRAK
Dua isolat bakteri endofit yang mampu mengendalikan Xanthomonas axonopodis pv. glycines, penyebab penyakit pustul kedelai, telah diperoleh pada penelitian sebelumnya. Bakteri tersebut perlu diformulasi agar dapat disimpan, ditransportasi, dan diaplikasikan. Penelitian bertujuan memperoleh formula bakteri endofit yang efektif dan dapat disimpan lama untuk pengendalian penyakit pustul bakteri. Penelitian menggunakan 3 macam bahan pembawa formula bakteri endofit (tanah gambut, air kelapa + 1% minyak sawit, dan tepung tapioka) yang dikombinasikan dengan lama penyimpanan (0, 1, 3, 5, dan 7 minggu). Perlakuan diaplikasi pada benih kedelai (perlakuan benih) sebelum ditanam. Kedelai diinokulasi dengan X. axonopodis pv. glycines pada umur 2 minggu dengan melukai permukaan bawah daun dan dioles dengan suspensi bakteri (kepadatan populasi 106 sel mL-1). Hasil penelitian menunjukkan bahwa hampir semua formula bakteri endofit mampu menekan intensitas pustul bakteri pada kedelai. Formulasi terbaik yang efektif mengendalikan penyakit pustul bakteri adalah bakteri endofit yang diformulasi dalam tanah gambut yang disimpan 1 dan 7 minggu (efektivitas 79.85% dan 77.02%) dan air kelapa + minyak sawit yang disimpan 3 minggu (efektivitas 77.46%).
Kata kunci: air kelapa, perlakuan benih, tanah gambut, tepung tapioka, Xanthomonas axonopodis
ABSTRACT
Two isolates of bacterial endophyte from soybean root were found to be effective to control bacterial pustule caused by Xanthomonas axonopodis pv. glycines. Formulation of the bacterial isolates is required to maintain the effectivity of this bacterial isolates during storage, transportation and application. The aim of this research was to obtain the best carrier for formulation to maintain the effectivity of bacterial endophyte in storage to control bacterial pustule on soybean. Three kind of carrier agent was evaluated for formulation of bacterial endophyte i.e. peat soil, tapioca flour and coconut water + 1% palm oil. Each carrier agent was combined with 5 treatment of storage time i.e. 0, 1, 3, 5 and 7 weeks. Soybean plants was inoculated by. X. axonopodis pv. glycines 2 weeks after planting by rubbing bacterial suspension (106 cel mL-1) on lower surface of leaves. The results showed that all formulas of bacterial endophyte were able to suppress the bacterial pustule on soybean. The best formulations were bacterial endophyte in peat soil stored for 1 and 7 weeks (effectivity rate 79.85% and 77.02%) and coconut water + palm oil and storaged for 3 weeks (effectivity rate 77.46%).
Key words: bacterial endophyte, formulation, soybean, Xanthomonas axonopodis pv. glycines
*Alamat penulis korespondensi: Jurusan Hama dan Penyakit Tumbuhan, Fakultas Pertanian, Universitas Andalas Padang, Kampus Limau Manis, Padang 25163.Tel: 0751-7059087, Faks: 0751-72702, Surel: [email protected]
51

J Fitopatol Indones Habazar et al.
PENDAHULUAN
Pustul bakteri yang disebabkan oleh Xanthomonas axonopodis pv. glycines merupakan salah satu penyakit utama pada kedelai di Indonesia (Khaeruni et al. 2007). Pengendalian pustul bakteri yang dianjurkan ialah dengan penggunaan varietas tahan, penggunaan bakterisida (Sinclair dan Backman 1989).
Bakteri endofit dapat mengendalikan penyakit hawar bakteri oleh Xanthomonas axonopodis pv. malvacearum pada kapas (Rajendran et al. 2006). Curtobacterium flaccumfaciens sebagai endofit pada tanaman jeruk mampu meningkatkan ketahanan jeruk terhadap Xylella fastidiosa (Araujo et al. 2002). Bakteri endofit dari kentang menghambat perkembangan bakteri patogen dari genus Erwinia dan Xanthomonas (Sessitsch et al. 2004). Habazar et al. (2012) melaporkan bahwa 2 isolat bakteri endofit akar kedelai (ST4E1.1 dan ST1E1.1) adalah isolat terbaik untuk mengendalikan pustul bakteri pada kedelai dengan efektivitas 45.1% dan 19.6%.
Penggunaan rizobakteri termasuk bakteri endofit dalam bentuk suspensi sel dapat menurunkan kemampuannya mengendalikan penyakit tanaman pada kondisi di lapangan. Untuk itu bakteri endofit perlu dimobilisasi dalam bentuk formulasi dengan bahan pem-bawa untuk mempertahankan daya hidupnya, memudahkan aplikasi, penyimpanan, dan pemasaran. Formulasi bakteri antagonis dalam bentuk cair dan tepung talkum digunakan untuk pengendalian beberapa jenis patogen tanaman, seperti hawar daun bakteri pada kapas oleh X. axonopodis pv. malvacearum (Rajendran et al. 2006). Aktivitas biokontrol B. megaterium B1301 yang diformulasi dalam tepung jagung baik untuk pengendalian layu bakteri oleh Ralstonia solanacearum ras 4 pada jahe (Yang et al. 2012). Tepung tapioka merupakan bahan pembawa terbaik untuk formulasi Pseudomonas fluorescens (Advinda 2009) dan formulasi bakteri rizoplan untuk mengendalikan X. axonopodis pv. alii pada bawang merah (Farlina et al. 2009). Penelitian ini bertujuan mendapatkan formula
bakteri endofit yang efektif dan stabil dalam pengendalian pustul bakteri pada kedelai.
BAHAN DAN METODE
Formula bakteri endofit diuji secara in vitro di laboratorium dan in planta di rumah kaca. Percobaan disusun dalam rancangan acak lengkap dengan 16 perlakuan dan 3 ulangan. Tiga macam bahan pembawa formula bakteri endofit: tanah gambut, air kelapa + 1% minyak sawit, dan tepung tapioka; dikombinasikan dengan lama penyimpanan: 0, 1, 3, 5, dan 7 minggu dan kontrol (diinokulasi dengan X. axonopodis pv. glycines). Perlakuan diaplikasi pada benih kedelai (perlakuan benih) sebelum ditanam. Kedelai diinokulasi dengan X. axonopodis pv. glycines pada umur 2 minggu dengan melukai permukaan bawah daun dan dioles dengan suspensi bakteri (kepadatan populasi 106 sel mL-1). Data dianalisis dengan uji Duncan pada taraf nyata 5%.
Perbanyakan Bakteri Endofit
Bakteri endofit isolat ST1E1.1 berasal dari penelitian Habazar et al. (2012) yang dikoleksi di Laboratorium Bakteriologi, Jurusan Hama dan Penyakit Tumbuhan, Fakultas Pertanian, Universitas Andalas Padang. Bakteri diremajakan pada medium agar-agar nutrien (AN) dan diinkubasi 48 jam,kemudian dipindahkan satu koloni ke dalam 25 mL medium cair nutrien dalam labu Erlenmeyer (volume 250 mL) dan diinkubasi 24 jam pada alat pengocok horizontal dengan kecepatan 250 rpm. Sebanyak 5 mL suspensi bakteri endofit ini dipindahkan ke dalam 500 mL air kelapa steril dalam labu Erlenmeyer (volume 1 L) dan diinkubasi 2 × 24 jam dengancara yang sama. Kepadatan populasi ditentukan melalui kekeruhannya dengan pembanding pada larutan McFarland skala 8 (diperkirakan 108 sel mL-1) (Klement et al. 1990).
Formulasi Bakteri Endofit
Formulasi bakteri endofit dibuat 3 macam formula, yaitu 2 padatan dan 1 cairan.Formula padat menggunakan bahan pembawa tapioka + 5% sukrosa dan tanah gambut + 5%
52

J Fitopatol Indones Habazar et al.
sukrosa. Sebanyak 50 g formula dimasukkan ke dalam kantong plastik tahan panas dan disterilkan pada suhu 121 °C selama 15 menit. Formula cair bahan pembawanya terdiri atas 98 mL air kelapa, 1 mL minyak sawit (1%) dan sukrosa 5%. Campuran bahan tersebut dimasukkan ke dalam labu Erlenmeyer. Formula ini disterilkan dengan cara yang sama dengan formula kering. Selanjutnya pada formula (padat dan cair) ditambahkan 1 mL suspensi isolat bakteri endofit (108 sel mL-1) dan disimpan (0, 1, 3, 5 dan 7 minggu) (Gambar 1) dan diinkubasi dalam ruangan pada suhu ± 20 °C dengan penerangan lampu 10 W.
Populasi bakteri endofit dari masing-masing formula diuji viabilitasnya pada beberapa pengenceran. Sebanyak 10 g formula kering dan 10 mL formula cair diambil dan diencerkan secara seri (10-1 sampai 10-6) dalam tabung reaksi steril. Dari pengenceran 10-6 diambil 1 mL, dipindahkan ke medium AN dan diinkubasi selama 2 × 24 jam. Jumlah koloni yang tumbuh dan kepadatan populasinya dihitung menggunakan rumus
JB = A × C, dengan
JB, Populasi bakteri (CFU mL-1); A, Jumlah koloni; C, tingkat pengenceran.
Uji Kestabilan Formula Bakteri Endofit untuk Mengendalikan Penyakit Pustul Bakteri pada Tanaman Kedelai
Tiga formula bakteri endofit (gambut, tepung tapioka, dan air kelapa + 1% minyak sawit) yang disimpan selama 0, 1, 3, 5 dan 7 minggu di ruang berpendingin, kemudian diuji kemampuannya dalam menekan pustul
bakteri pada tanaman kedelai di rumah kaca. Bakteri endofit diintroduksi 2 kali, yaitu pada benih dan tanaman kedelai. Pada benih, formula bakteri endofit disuspensikan dalam air (1:100), kemudian benih kedelai varietas Wilis direndam dalam suspensi tersebut selama 15 menit dan dikeringanginkan 24 jam. Benih kedelai ditanam dalam kantung plastik dengan medium tanam 5 kg campuran pupuk kandang dan tanah steril (2:1 v/v). Introduksi bakteri endofit pada tanaman kedelai umur 3 minggu dilakukan dengan menyiramkan suspensi formula bakteri endofit di sekeliling batang; kira-kira 3 cm dari pangkal batang.
Perbanyakan dan Inokulasi X. axonopodis pv. glycines
Isolat X. axonopodis pv. glycines diremajakan pada medium agar-agar glukosa nutrien selama 5 hari, lalu koloni bakteri disuspensikan dalam 9 mL akuades steril. Suspensi X. axonopodis pv. glycines yang digunakan dengan kepadatan populasi 106 sel mL-1 ditentukan dengan cara yang sama seperti untuk bakteri endofit. Sebanyak 3 daun muda dari tanaman yang berumur 50 hari,dilukai dengan jarum pentul dan diinokulasi dengan mengolesnya, lalu tanaman diselubungi dengan plastik bening selama 5 hari.
Tanaman dipupuk, dibersihkan dari gulma. Tanaman kedelai dipupuk dengan 0.23 g urea, 0.30 g SP 36, dan 0.45 g KCl per kantong plastik. Polong kedelai dipanen umur 88 hari.
Peubah yang diamati ialah viabilitas bakteri endofit dalam formula, perkembangan pustul bakteri (masa inkubasi, persentase dan intensitas pada daun dan polong). Reaksi peningkatan ketahanan kedelai terhadap pustul bakteri mengacu pada kriteria Dirmawati (2005) yang didasarkan pada intensitas serangan, yaitu <10%, tahan; 10–20%, agak tahan; 20–30%, agak rentan; 30–60%, rentan;dan >60%, sangat rentan. Efektivitas peng-gunaan bakteri endofit untuk memperpanjang masa inkubasi atau menurunkan persentase dan intensitas serangan pustul bakteri pada daun dan polong dihitung menggunakan rumus sebagai berikut:
53
Gambar 1 Formula bakteri endofit yang disimpan pada suhu 20 °C. a, Air kelapa + minyak sawit; b, Tanah gambut; c, Tepung tapioka.
a b c

J Fitopatol Indones Habazar et al.
E, efektivitas; P, perlakuan; K, kontrol.
HASIL
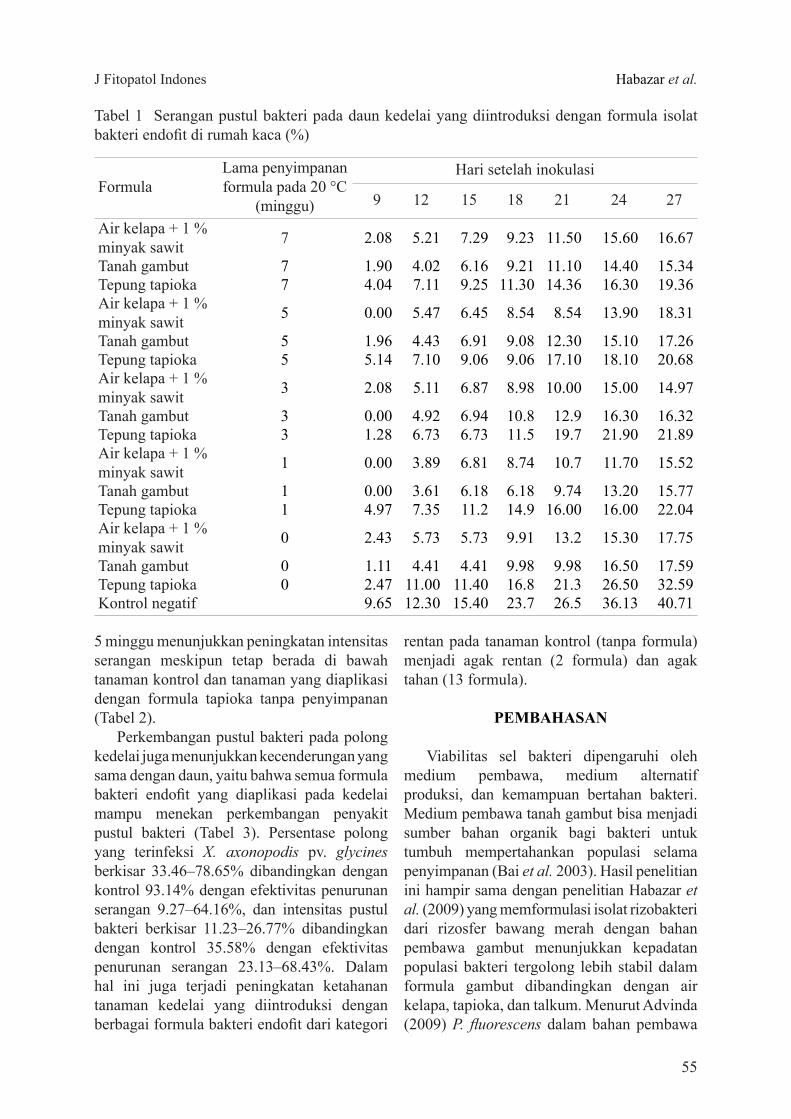
Viabilitas Bakteri Endofit dalam FormulaViabilitas bakteri endofit dalam formula
berbagai bahan pembawa (gambut, tapioka, dan air kelapa + 1% minyak sawit) terlihat berfluktuasi pada tahap awal dan cenderung menurun setelah disimpan sampai 7 minggu (Gambar 2). Meskipun demikian, dalam formula gambut terjadi peningkatan populasi bakteri endofit setelah disimpan 1 minggu (9.8 × 109 CFU g-1), tetapi kemudian menurun (109 CFU g-1). Viabilitas bakteri endofit dalam formula air kelapa relatif stabil, sedangkan dalam formula tepung tapioka meningkat setelah disimpan 3 minggu dan kemudian menurun setelah disimpan 5 dan 7 minggu. Kondisi ini masih memenuhi syarat viabilitas bakteri dalam suatu formula karena umumnya aplikasi bakteri endofit baik dalam bentuk perlakuan benih ataupun penyiraman pada tanah perakaran menggunakan kepadatan populasi 108 CFU g-1.
Perkembangan Penyakit Pustul BakteriSemua formula bakteri endofit yang
diinokulasikan pada tanaman kedelai mampu memperlambat masa inkubasi pustul bakteri 0.33–3.00 hari dibandingkan dengan kontrol
yang efiktivitasnya 3.67–33.33%. Masa inkubasi pustul bakteri pada kedelai yang paling lama ialah yang diperlakukan dengan bakteri endofit formula air kelapa + 1% minyak sawit disimpan 1 minggu. Formula bakteri endofit tersebut mampu menghambat perkembangan pustul bakteri pada daun, persentase daun sakit berkisar 14.97–32.20% dibandingkan dengan kontrol 49.08% dengan efektivitas 34.39–69.49%, sedangkan intesitas penyakit berkisar antara 4.13–12.00% dibandingkan dengan kontrol 20.50% dengan efektivitas 41.46–79.85%. Berdasarkan kategori ketahanan tanaman kedelai terhadap pustul bakteri, maka tanaman yang diinokulasi dengan formula bakteri endofit menunjukkan peningkatan ketahanan dari agak rentan pada tanaman kontrol menjadi agak tahan (1 formula yaitu bakteri endofit dalam tapioka tanpa penyimpanan) dan selebihnya menjadi tahan (Tabel 1).
Hampir semua formula yang diaplikasi pada kedelai mempunyai kemampuan yang hampir sama dalam menekan perkembangan persentase pustul bakteri, kecuali pada formula tapioka yang tidak disimpan.Perkembangan persentase serangan pustul bakteri pada kedelai yang diberi perlakuan formula ini cenderung meningkat, tetapi tetapdi bawah tanaman kontrol. Demikian juga halnya dengan perkembangan intensitas serangan pustul bakteri, namun terdapat 2 formula tapioka, yaitu yang disimpan 3 dan
54
E = P – KK × 100%, dengan
10.09.89.69.49.29.08.88.68.4
0 1 3 5 7
Popu
lasi
bak
teri
endo
fit
(log
CFU
mL-1
ata
u g-1
)
Lama penyimpanan (minggu)
Gambar 2 Perkembangan kepadatan populasi beberapa formula bakteri endofit akar setelah disimpan selama 7 minggu. , AK + 1% MS; , Gambut; , Tapioka.
J Fitopatol Indones Habazar et al.
55
5 minggu menunjukkan peningkatan intensitas serangan meskipun tetap berada di bawah tanaman kontrol dan tanaman yang diaplikasi dengan formula tapioka tanpa penyimpanan (Tabel 2).
Perkembangan pustul bakteri pada polong kedelai juga menunjukkan kecenderungan yang sama dengan daun, yaitu bahwa semua formula bakteri endofit yang diaplikasi pada kedelai mampu menekan perkembangan penyakit pustul bakteri (Tabel 3). Persentase polong yang terinfeksi X. axonopodis pv. glycines berkisar 33.46–78.65% dibandingkan dengan kontrol 93.14% dengan efektivitas penurunan serangan 9.27–64.16%, dan intensitas pustul bakteri berkisar 11.23–26.77% dibandingkan dengan kontrol 35.58% dengan efektivitas penurunan serangan 23.13–68.43%. Dalam hal ini juga terjadi peningkatan ketahanan tanaman kedelai yang diintroduksi dengan berbagai formula bakteri endofit dari kategori
rentan pada tanaman kontrol (tanpa formula) menjadi agak rentan (2 formula) dan agak tahan (13 formula).
PEMBAHASAN
Viabilitas sel bakteri dipengaruhi oleh medium pembawa, medium alternatif produksi, dan kemampuan bertahan bakteri. Medium pembawa tanah gambut bisa menjadi sumber bahan organik bagi bakteri untuk tumbuh mempertahankan populasi selama penyimpanan (Bai et al. 2003). Hasil penelitian ini hampir sama dengan penelitian Habazar et al. (2009) yang memformulasi isolat rizobakteri dari rizosfer bawang merah dengan bahan pembawa gambut menunjukkan kepadatan populasi bakteri tergolong lebih stabil dalam formula gambut dibandingkan dengan air kelapa, tapioka, dan talkum. Menurut Advinda (2009) P. fluorescens dalam bahan pembawa
Tabel 1 Serangan pustul bakteri pada daun kedelai yang diintroduksi dengan formula isolat bakteri endofit di rumah kaca (%)
FormulaLama penyimpanan formula pada 20 °C
(minggu)
Hari setelah inokulasi
9 12 15 18 21 24 27
Air kelapa + 1 % minyak sawit 7 2.08 5.21 7.29 9.23 11.50 15.60 16.67
Tanah gambut 7 1.90 4.02 6.16 9.21 11.10 14.40 15.34Tepung tapioka 7 4.04 7.11 9.25 11.30 14.36 16.30 19.36Air kelapa + 1 % minyak sawit 5 0.00 5.47 6.45 8.54 8.54 13.90 18.31
Tanah gambut 5 1.96 4.43 6.91 9.08 12.30 15.10 17.26Tepung tapioka 5 5.14 7.10 9.06 9.06 17.10 18.10 20.68Air kelapa + 1 % minyak sawit 3 2.08 5.11 6.87 8.98 10.00 15.00 14.97
Tanah gambut 3 0.00 4.92 6.94 10.8 12.9 16.30 16.32Tepung tapioka 3 1.28 6.73 6.73 11.5 19.7 21.90 21.89Air kelapa + 1 % minyak sawit 1 0.00 3.89 6.81 8.74 10.7 11.70 15.52
Tanah gambut 1 0.00 3.61 6.18 6.18 9.74 13.20 15.77Tepung tapioka 1 4.97 7.35 11.2 14.9 16.00 16.00 22.04Air kelapa + 1 % minyak sawit 0 2.43 5.73 5.73 9.91 13.2 15.30 17.75
Tanah gambut 0 1.11 4.41 4.41 9.98 9.98 16.50 17.59Tepung tapioka 0 2.47 11.00 11.40 16.8 21.3 26.50 32.59Kontrol negatif 9.65 12.30 15.40 23.7 26.5 36.13 40.71

J Fitopatol Indones Habazar et al.
56
Tabel 2 Intensitas serangan pustul bakteri pada daun kedelai yang diintroduksi dengan formula isolat bakteri endofit di rumah kaca (%)
FormulaLama penyimpanan formula pada 20 °C
(minggu)
Hari setelah inokulasi
9 12 15 18 21 24 27
Air kelapa + 1 % minyak sawit 7 0.41 0.83 1.46 2.29 3.33 4.37 6.25
Tanah gambut 7 0.38 0.80 1.23 2.02 2.83 4.53 4.71Tepung tapioka 7 0.80 1.42 2.06 2.67 3.69 5.67 5.66Air kelapa + 1 % minyak sawit 5 0.00 1.09 1.29 1.91 3.21 4.99 6.85
Tanah gambut 5 0.39 0.89 1.38 1.81 3.11 4.47 5.35Tepung tapioka 5 0.39 1.41 2.00 2.71 5.11 6.33 7.56Air kelapa + 1 % minyak sawit 3 0.41 1.02 1.37 1.97 2.18 4.61 4.62
Tanah gambut 3 0.00 0.98 1.38 2.45 2.87 5.14 6.03Tepung tapioka 3 0.26 1.35 2.33 2.55 5.41 7.66 8.55Air kelapa + 1 % minyak sawit 1 0.00 0.79 1.75 2.35 2.92 3.30 4.80
Tanah gambut 1 0.00 0.72 1.23 1.69 2.22 2.00 4.10Tepung tapioka 1 0.99 1.74 2.50 2.97 3.39 5.62 7.10Air kelapa + 1 % minyak sawit 0 0.46 1.15 1.74 2.21 3.60 4.78 6.00
Tanah gambut 0 0.22 0.88 1.56 2.21 2.86 5.07 5.50Tepung tapioka 0 0.49 2.20 2.77 3.89 6.24 9.13 12.00Kontrol negatif 2.23 2.77 4.23 6.74 8.79 10.72 15.72
Formula
Lama penyimpanan formula pada
20 °C (minggu)
Persentase polong terserang Intensitas polong terserang
% Efektivitas (%) % Efektivitas
(%)Reaksi
ketahanan*Tanah gambut 0 45.61 51.19 14.64 efgh 58.85 Agak tahanTanah gambut 1 33.46 64.19 11.23 h 68.43 Agak tahanTanah gambut 3 45.55 51.25 14.36 fgh 59.39 Agak tahanTanah gambut 5 42.96 54.02 12.23 gh 65.65 Agak tahanTanah gambut 7 54.76 34.83 17.53 cdef 50.73 Agak tahanAir kelapa + 1% minyak sawit 0 48.86 47.70 15.7 defgh 55.62 Agak tahanAir kelapa + 1% minyak sawit 1 48.12 48.50 16.49 defg 55.62 Agak tahanAir kelapa + 1% minyak sawit 3 47.64 49.01 15.56 defg 56.27 Agak tahanAir kelapa + 1% minyak sawit 5 52.28 37.49 17.40 cdef 53.65 Agak tahanAir kelapa + 1% minyak sawit 7 57.66 31.73 19.13 cde 46.23 Agak tahanTepung tapioka 0 56.28 33.20 19.65 b 44.77 Agak tahanTepung tapioka 1 49.81 46.69 18.44 cdef 48.17 Agak tahanTepung tapioka 3 48.19 41.87 21.60 c 39.29 Agak rentanTepung tapioka 5 52.28 31.17 17.44 cdef 52.20 Agak tahanTepung tapioka 7 78.65 9.27 26.77 b 23.13 Agak rentanKontrol 93.44 0.00 35.58 a 0.00 RentanKK 17.53 23.82
*Kriterium ketahanan berdasarkan intensitas serangan menurut Dirmawati (2005) Angka yang diikuti dengan huruf yang sama menunjukkan tidak berbeda nyata.
Tabel 3 Perkembangan pustul bakteri pada polong kedelai yang diintroduksi dengan formula isolat bakteri endofit di rumah kaca

J Fitopatol Indones Habazar et al.
57
tepung tapioka lebih tinggi viabilitasnya dibandingkan dengan agar-agar dan alginat, kepadatan populasinya meningkat setelah disimpan 6 minggu dan menurun setelah 8 minggu. Dengan demikian viabilitas bakteri endofit lebih baik dalam formula gambut dibandingkan dengan air kelapa dan tapioka.
Formula bakteri endofit isolat ST1E1.1 dengan 3 macam bahan pembawa (gambut, tepung tapioka, dan air kelapa + 1% minyak sawit) yang disimpan sampai 7 minggu, kemudian diintroduksi pada benih kedelai menunjukkan kemampuan yang lebih baik dalam menghambat perkembangan intensitas serangan pustul bakteri. Penelitian terdahulu dengan perlakuan bakteri endofit isolat ST1E1.1 yang diintroduksi pada benih kedelai menurunkan intensitas serangan pustul bakteri dengan efektivitas 19.6% (Habazar et al. 2012). Habazar et al. (2009) juga melaporkan bahwa tanaman bawang merah yang diberi perlakuan rizobakteri isolat JB2sktE1 dalam formula tepung tapioka menunjukkan intensitas serangan hawar daun bakteri (X. axonopodis pv. alii) yang rendah (7.80%) dengan efektivitas 71.25%. Sebagian besar formula bakteri endofit mampu meningkatkan ketahanan kedelai yang tergolong agak tahan (1 formula tepung tapioka tanpa disimpan) dan selebihnya tahan (14 formula yang disimpan 1, 3, 5 dan 7 minggu), sedangkan tanaman kontrol tergolong agak rentan seperti yang dilaporkan Dirmawati (2005).
Umumnya intensitas pustul pada polong dari tanaman yang diinokulasi dengan formula bakteri endofit lebih rendah dibandingkan dengan kontrol. Seperti halnya intensitas serangan pustul bakteri pada daun kedelai, terjadi peningkatan ketahanan kedelai ter-hadap pustul bakteri. Kondisi ini juga telah dilaporkan oleh Habazar et al. (2012) bahwa 2 isolat bakteri endofit yang diinokulasi pada kedelai dapat meningkatkan ketahanan kedelai terhadap pustul bakteri. Artinya bahwa isolat bakteri endofit yang diformulasi menunjukkan pengaruh yang stabil dalam pengendalian pustul bakteri pada kedelai.
Formula bakteri antagonis telah digunakan dalam mengendalikan X. axonopodis pv.
malvacearum penyebab penyakit hawar daun bakteri pada kapas dalam bentuk formulasi cair dan tepung talkum (Rajendran et al. 2006). Aktivitas biokontrol B. megaterium B1301 yang diformulasi dalam tepung jagung baik untuk pengendalian penyakit layu bakteri oleh Ralstonia solanacearum ras 4 pada jahe (Yang et al. 2012). Dari 8 formula P. fluorescens yang diperlakukan pada benih kapas efektif mengendalikan penyakit rebah kecambah sampai 62.5% dibandingkan dengan kontrol,hanya formula TAL-B2 (suspensi P. fluorescens galur CKK-3) mengandung 9 × 108 CFU mL-1 dicampur dengan tepung talkum (1 kg) dan CMC (10 g) yang kurang efektif (Ardakani et al. 2010). Penggunaan tepung tapioka sebagai bahan pembawa telah dilaporkan Habazar et al. (2009) yang menunjukkan kemampuan yang sama dengan tepung talkum. Farlina et al. (2009) melaporkan bahwa formulasi bakteri rizoplan dalam tepung tapioka yang disimpan 2 minggu menunjukkan intensitas daun bawang terserang X. axonopodis pv. alii paling rendah.
Berdasarkan hasil penelitian dapat disimpulkan bahwa hampir semua formula isolat bakteri endofit (ST1E1.1) yang disimpan 1, 3, 5, dan 7 minggu yang diinokulasi pada benih kedelai menunjukkan kemampuan menekan intensitas pustul bakteri. Formula yang terbaik ialah gambut yang disimpan 1 dan 7 minggu dan formula air kelapa + 1% minyak sawit yang disimpan 3 minggu.
UCAPAN TERIMA KASIH
Penelitian ini dibiayai oleh DP2M DIKTI DEPDIKNAS melalui Hibah Kompetensi Tahun 2012 dengan Kontrak No. 063/UN.16/PL/Hikom/2012.
DAFTAR PUSTAKA
Advinda L. 2009. Tanggap fisiologis tanaman pisang yang diintroduksi dengan formula Pseudomonad fluoresen terhadap blood disease bacteria (BDB) [disertasi]. Padang (ID): Universitas Andalas.

J Fitopatol Indones Habazar et al.
Araujo WL, Marcon J, Maccheroni WJr, Van Elsas JD, Van Vuurde JWL, and Azevedo JL. 2002. Diversity of endophytic bacterial populations and their interaction with Xylella fastidiosa in citrus plants. Appl. Environ. Microbiol. 68:4906-4914. DOI: http://dx.doi.org/10.1128/AEM.68.10.4906-4914.2002.
Ardakani SS, Heydari A, Khorasani N, Arjmandi R. 2010. Development of new bioformulations of Pseudomonas fluorescens and evaluation of these products against damping-off of cotton seedlings. J Plant Pathol. 92(1):83–88.
Bai Y, Zhuo X, Smith DL. 2003. Enhanced soybean plant growth resulting from co inoculation of Bacillus strains with Bradyrhizobium japonicum. Crop Sci. 43:1774–1781. DOI: http://dx.doi.org/10.2135/cropsci2003.1774.
Dirmawati SR. 2005. Penurunan intensitas penyakit pustul bakteri kedelai melalui strategi cara tanam tumpangsari dan penggunaan agensia hayati. J Agrijati. 1(1):1–11.
Farlina R, Habazar T, Yanti Y. 2009. Stabilitas beberapa formula isolat bakteri rizoplan dalam penyimpanan dan kemampuannya menekan penyakit hawar daun bakteri (Xanthomonas axonopodis pv.alii) pada tanaman bawang merah. Manggaro. 10(2):49–54.
Habazar T, Nasrun, Jamsari, Rusli I, Ernita M, Irfandri, Resti Z, Yanti Y. 2009. Introduction of rhizobacteria indigenous strains from healthy onion rhizosphere to control Xanthomonas leaf blight disease on onion. International Seminar and Workshop Biodivesity, Biotechnology,
and Crop Production. Padang (ID): PBPI Komisariat Sumatera Barat.
Habazar T, Resti Z, Yanti Y, Trisno J, Diana A. 2012. Penapisan bakteri endofit akar kedelai secara in planta untuk mengendalikan penyakit pustul bakteri. J Fitopatol Indones. 8(4):97–103. DOI: http://dx.doi.org/ 10.14692/jfi.8.4.97.
Khaeruni A, Suwanto A, Tjahjono B, Sinaga SM. 2007. Deteksi cepat penyakit pustul bakteri pada kedelai menggunakan teknik PCR dengan primer spesifik. Hayati. 14(2):76–80.
Klement Z, Rudolph K, Sand DC. 1990. Methods in phytobacteriology. Budapest (HU): Akademiai Kiado.
Rajendran L. Saravanakumar D, Ragunchander T, Samiyappan R. 2006. Endophytic bacterial induction of defence enzymes againts bacterial blight of cotton. Tamil Nadu (IN): Tamil Nadu.
Sessitsch A, Reiter B, Berg G. 2004. Endophytic bacterial communities of field-grown potato plants and their plant-growth-promoting and antagonistic abilities. Can J Microbiol. 50:239–249. DOI: http://dx.doi.org/10.1139/w03-118.
Sinclair JB, Backman PA. 1989. Compendium of Soybean Diseases. Ed ke-3. Minnesota (US): The American Phytopathological Society.
Yang W, Liu H, Wang Y, Luo Y, Yang H, Guo J.2012. Effects of two different soil amendments on the biocontrol efficacy of biological control agents against ralstonia wilt on ginger. Afr J Biotechnol. 11(39):9383–9390. DOI: http://dx.doi.org/10.5897/AJB11.4317.
58

Volume 11, Nomor 2, April 2015Halaman 59–67
DOI: 10.14692/jfi.11.2.59ISSN: 0215-7950
*Alamat penulis korespondensi: Departemen Proteksi Tanaman, Fakultas Pertanian, Institut Pertanian Bogor, Jalan Kamper, Kampus Darmaga IPB, Bogor 16680Tel: 0251-8629364, Faks: 0251-8629362, Surel: [email protected]
59
Deteksi Virus yang Menginfeksi Kedelai di Jawa
Detection of Viruses Infecting Soybean in Java
Yunita Fauziah Rahim*, Tri Asmira Damayanti, Munif GhulamahdiInstitut Pertanian Bogor, Bogor 16680
ABSTRAK
Infeksi virus merupakan salah satu kendala produksi kedelai. Penelitian dilakukan untuk mendeteksi infeksi virus pada beberapa pertanaman kedelai di Jawa. Sampel daun diambil secara acak sebanyak 50 tanaman dari tiap lokasi pertanaman kedelai di Bogor, Cirebon, Bantul, dan Ponorogo. Pengamatan dilakukan terhadap gejala yang ditemukan di lapangan dan insidensi penyakit ditentukan secara serologi menggunakan antibodi Cucumber mosaic virus (CMV), Soybean mosaic virus (SMV), Cowpea mild mottle virus (CPMMV), dan Bean pod mottle virus (BPMV). Insidensi CMV, SMV, dan CPMMV berturut-turut berkisar 72–84%, 14–24%, dan 6–8%; sedangkan infeksi BPMV tidak ditemukan. Pita DNA spesifik CMV, Potyvirus, dan Geminivirus berhasil diamplifikasi berturut-turut menggunakan primer spesifik gen protein selubung CMV, primer universal Potyvirus dan Geminivirus. Hasil sikuensing menunjukkan bahwa homologi CMV tertinggi adalah terhadap CMV galur S asal Bogor (99%), homologi Potyvirus tertinggi terhadap BCMV isolat kacang hijau asal Cina dan galur Blackeye asal Vietnam (90%), dan homologi Geminivirus tertinggi terhadap Pepper yellow leaf curl virus (PYLCV) asal Bogor dan Jawa (96%). Analisis filogenetika menunjukkan bahwa CMV-S berada dalam satu kelompok dan terpisah dari CMV galur lainnya. BCMV pada kedelai dalam penelitian ini membentuk kelompok terpisah dari BCMV asal negara lain, sedangkan PYLCV isolat kedelai membentuk satu kelompok dengan PYLCV Bogor dan Jawa.
Kata kunci: Cucumber mosaic virus, Soybean mosaic virus, Cowpea mild mottle virus,Pepper yellow leaf curl virus
ABSTRACT
Virus infection is an important production constraint for soybean. Research was conducted to detect virus infection from soybean samples collected from several locations in Java. Leave samples from 50 plants was taken randomly from each location in Bogor, Cirebon, Bantul, and Ponorogo. Field symptoms was observed and disease incidence was determined based on serological assay using specific antibodies to Cucumber mosaic virus (CMV), Soybean mosaic virus (SMV), Cowpea mild mottle virus (CPMMV), and Bean pod mottle virus (BPMV). Incidence of CMV, SMV, and CPMMV was 72–84%, 14–24%, and 6–8%, respectively; whereas infection of BPMV was not found. Specific viral DNA of CMV, Potyvirus, and Geminivirus was successfully amplified using specific primer for CMV coat protein, universal primer for Potyvirus and Geminivirus, respectively. Nucleotide sequence analysis showed that isolate CMV from soybean has the highest homology (99%) to CMV strain S, Potyvirus isolates has the highest homology (90%) to BCMV isolate Mungbean from China and BCMV strain Blackeye from Vietnam, and Geminivirus isolates has the highest homology (96%) to Pepper yellow leaf curl virus (PYLCV) from Bogor and Java. Phyllogenetic analysis showed that CMV strain S formed

J Fitopatol Indones Rahim et al.
60
a distinct group from other strains of CMV, BCMV from soybean in Java was in separate group from BCMV from other countries, while PYLCV isolate soybean was in the same group with PYLCV from Bogor and Java.
Key words: Cucumber mosaic virus, Soybean mosaic virus, Cowpea mild mottle virus,Pepper yellow leaf curl virus
PENDAHULUAN
Kedelai (Glycine max) merupakan salah satu komoditas penting di Indonesia, selain dapat dijadikan sebagai bahan konsumsi pangan juga dimanfaatkan sebagai bahan baku industri olahan. Pemanfaatan kedelai yang cukup tinggi belum diikuti dengan produksi yang mencukupi, bahkan cenderung terjadi penurunan dari tahun ke tahun. BPS (2013) melaporkan terjadinya penurunan produksi kedelai sejak tahun 2009–2012, yaitu berturut-turut sebesar 974.51 ton, 907.03 ton, 851.29 ton, dan 843.15 ton.
Kedelai rentan terhadap infeksi virus sehingga dapat memengaruhi hasil panen secara kualitas dan kuantitas. Beberapa virus yang dilaporkan menyebabkan penyakit penting pada pertanaman kedelai ialah Soybean mosaic virus (SMV) (Andayanie 2012), Soybean stunt virus (SSV)/Cucumber mosaic virus galur soybean (CMV-S) (Asadi et al. 2003), Cowpea mild mottle virus (CPMMV) (Akin 2003). Berdasarkan hasil penelitan Sulandari et al. (2006) di rumah kaca, ditemukan juga bahwa Geminivirus dapat menginfeksi kedelai.
Gejala infeksi virus di lapangan sangat bervariasi karena ekspresi gejala dipengaruhi oleh jenis virus, kondisi lingkungan, dan kultivar tanaman. Gejala penyakit yang tampak di lapangan tidak dapat diandalkan untuk mendiagnosis virus penyebab penyakit. Deteksi virus secara serologi dan deteksi asam nukleat dengan polymerase chain reaction (PCR) serta perunutan DNA yang berkembang saat ini dapat membantu identifikasi virus secara akurat. Mengingat kedelai merupakan salah satu komoditas pangan penting Indonesia maka perlu diketahui jenis virus yang dapat membatasi produksi kedelai. Oleh karena itu, penelitian ini bertujuan mendeteksi virus yang
menginfeksi kedelai pada beberapa lokasi pertanaman kedelai di Jawa.
BAHAN DAN METODE
Sampel tanaman kedelai diambil dari pertanaman di Kecamatan Cikarawang dan Kecamatan Bogor Barat, Bogor (Jawa Barat); Kecamatan Gempol, Cirebon (Jawa Barat); Kecamatan Kasihan, Bantul (DI Yogyakarta); dan Kecamatan Babadan, Ponorogo (Jawa Timur). Sampel diambil sebanyak masing-masing 50 daun tanaman bergejala dan tidak bergejala dari setiap lokasi.
Uji SerologiSampel tanaman dari lapangan digunakan
untuk mendeteksi virus berdasarkan metode serologi dot blot immunobinding assay (DIBA) menggunakan antibodi spesifik CMV, SMV, CPMMV, dan BPMV mengikuti protokol yang digunakan Anggraini dan Hidayat (2014), namun antibodi yang digunakan hanya antibodi spesifik kedua yang berkonjugasi dengan enzim alkalin fosfatase.
Insidensi infeksi virus (IIV) dihitung menggunakan rumus:
IIV = n
N × 100%, dengan
n, jumlah tanaman positif terdeteksi virus; N, total tanaman yang diuji.
Deteksi dengan Metode PCRVirus target terdiri atas CMV, SMV,
Potyvirus, dan Geminivirus. Tiga virus (CMV, SMV, dan Potyvirus) dideteksi dengan metode reverse transcription (RT)-PCR, sedangkan Geminivirus dengan metode PCR.
RNA total diekstraksi dari daun tanaman menggunakan GeneJET Plant RNA Purification Mini Kit dengan protokol sesuai yang direkomendasikan oleh pembuatnya (Thermo

J Fitopatol Indones Rahim et al.
61
Scientific). Untuk deteksi Geminivirus, DNA total diekstraksi mengikuti protokol yang dikembangkan oleh Doyle dan Doyle (1987). Sintesis complementary DNA (cDNA) dilakukan terhadap RNA total sesuai protokol yang dilaporkan Anjarsari et al. (2013).
Amplifikasi CMV menggunakan pasangan primer CMV-cpF [5’-GACAAATCTGAATCAACCAGTGCC-3’] dan CMV-cpR [5’-ACTGGGAGCACTC-CAGATGTG-3’] dengan target amplikon ~650 pb. Amplifikasi dilakukan sebanyak 1 siklus pada 95 °C selama 3 menit, 45 °C selama 1 menit dan 72°C, selanjutnya 35 siklus dengan tahapan denaturasi pada 94 °C selama 1 menit, aneling pada 50 °C selama 1 menit dan sintesis pada 72 °C selama 1 menit. Khusus untuk siklus terakhir ditambah tahapan sintesis 68 °C selama 5 menit. Amplifikasi SMV menggunakan pasangan primer spesifik gen CI, CI-F [5’-GCATTCAACTGTGCGCTTAAAGAAT-3’ dan CI-R [5’-TTGAGGCTGCAAAAATTAC TCACTT-3’] dengan target amplikon berukuran ~1385 pb dan program PCR sesuai yang dilakukan Kim et al. (2004). Potyvirus diamplifikasi menggunakan pasangan primer universal sebagian gen protein selubung (CP) Potyvirus MJ1 [5’-ATGGTHTGGTGTGYATHGARAAYGG-3’] dan MJ2 [5’TGCTGCKGCYTTCAT-YTG-3’] dengan target amplikon berukuran ~320 pb dan program PCR sesuai yang dilakukan Grisoni et al. (2006). Amplifikasi Geminivirus menggunakan pasangan primer SPG1 [5′-CCCCKGTGCGWRAATCCAT-3′] dan SPG2 [5′-ATCCVAAYWTYCAGGGAGCTAA-3′] dengan target amplikon berukuran ~900 pb
dan program PCR sesuai yang dilakukan Li et al. (2004).
DNA hasil amplifikasi dielektroforesis pada tegangan 50 volt selama 50 menit menggunakan gel agarosa 1% yang dilarutkan dalam bufer 0.5 × Tris-Borate EDTA (TBE) kemudian direndam dalam larutan etidium bromida selama 15 menit. Visualisasi DNA dilakukan menggunakan UV transiluminator dan didokumentasi dengan kamera digital.
Perunutan DNA dan Analisis FilogenetikaPerunutan DNA hasil amplifikasi dilakukan
dengan mengirim sampel DNA ke First Base, Malaysia. Sikuen nukleotida dibandingkan dengan sikuen nukleotida virus asal negara lain yang terdaftar di GenBank menggunakan program basic local alignment search tool (BLAST) pada situs National Center for Biotechnology Information (www.ncbi.nlm.nih.gov).
Analisis filogenetika dilakukan mengguna-kan perangkat lunak molecular evolutionary genetics analysis (MEGA 6.06) dengan model pohon neighbor joining (bootstrap sebanyak 1000 kali).
HASIL
Gejala PenyakitGejala yang beragam ditemukan pada
tanaman kedelai di beberapa lokasi di Jawa (Tabel 1). Gejala yang ditemukan umumnya mosaik, permukaan daun yang tidak merata/melepuh, penebalan tulang daun, malformasi daun, pinggiran daun melengkung ke atas (cupping) dan menguning (Gambar 1).
LokasiJenis gejala*
Mosaik Rugos Vein banding Malformasi daun YellowingCikarawang √ √ √ √ √Bogor Barat √ - √ √ √Cirebon √ √ - √ -Bantul √ √ √ - -Ponorogo √ √ √ - -
Tabel 1 Jenis gejala infeksi virus pada tanaman kedelai di beberapa lokasi di Jawa
*Rugos, permukaan daun tdak rata/melepuh; Vein banding, penebalan tulang daun; √, ditemukan gejala; -,tidak ditemukan gejala.

J Fitopatol Indones Rahim et al.
Gambar 1 Jenis gejala infeksi virus pada tanaman kedelai di beberapa lokasi di Jawa. a, mosaik; b, mosaik, daun mengecil, permukaan daun tidak rata, tepi daun melengkung; c, daun mengecil, penebalan tulang daun; d, malformasi daun, menguning, tepi daun melengkung ke atas; dan e, tepi daun menguning.
a b c
d e
62
Hasil deteksi secara serologi menunjukkan bahwa gejala mosaik dapat disebabkan oleh infeksi tunggal maupun infeksi campuran oleh 2 atau lebih virus. Infeksi campuran dapat menyebabkan gejala mosaik yang lebih berat, malformasi daun dan menguning.
Insidensi Infeksi VirusHasil uji serologi menunjukkan infeksi
CMV paling tinggi dibandingkan dengan infeksi SMV dan CPMMV (Tabel 2). Insidensi penyakit CMV berturut-turut ialah 84, 72,
78, 74, dan 82% untuk daerah Cikarawang, Bogor Barat, Cirebon, Bantul, dan Ponorogo. Beberapa sampel terinfeksi campuran antara CMV dan virus lainnya, yaitu 8 sampel (16%) asal Cikarawang, 12 sampel (24%) asal Cirerbon, 6 sampel (12%) asal Bantul, dan 10 sampel (20%) asal Ponorogo (Tabel 3).
Amplifikasi DNAFragmen DNA CMV berhasil diamplifikasi
dengan primer spesifik gen CP (~650 pb) dari semua sampel (Gambar 2a). Fragmen DNA
LokasiInsidensi infeksi virus (n/N)*
SMV CMV CPMMVCikarawang 16.0 (8/50) 84.0 (42/50) 6.0 (3/50)Bogor Barat 0.0 (0/50) 72.0 (36/50) 0.0 (0/50)Cirebon 24.0 (12/50) 78.0 (39/50) 8.0 (4/50)Bantul 14.0 (7/50) 74.0 (37/50) 0.0 (0/50)Ponorogo 22.0 (11/50) 82.0 (41/50) 0.0 (0/50)Total 15.2 (38/205) 78.0 (195/250) 2.8 (7/250)
*n/N, jumlah tanaman terinfeksi/jumlah total tanaman yang diuji
Tabel 2 Insidensi infeksi (%) virus berdasarkan reaksi serologi

J Fitopatol Indones Rahim et al.
Lokasi Infeksi CampuranSMV-CMV SMV-CPMMV CMV-CPMMV SMV-CMV-CPMMV
Cikarawang 6/50 0/50 1/50 1/50Bogor Barat 0/0 0/50 0/50 0/50Cirebon 8/50 0/50 2/50 2/50Bantul 6/50 0/50 0/50 0/50Ponorogo 10/50 0/50 0/50 0/50
*Frekuensi infeksi ialah jumlah tanaman terinfeksi/jumlah total tanaman yang diamati
Tabel 3 Frekuensi terjadinya infeksi campuran virus berdasarkan reaksi serologi*
SMV tidak berhasil diamplifikasi dengan primer spesifik gen CI SMV dari sampel asal Ponorogo (data tidak ditampilkan) namun dari sampel yang sama berhasil diamplifikasi Potyvirus (~320 pb) (Gambar 2b). Sampel daun yang bergejala melengkung ke atas serta menguning yang hanya ditemukan di lokasi Bogor Barat terdeteksi positif CMV dan Geminivirus (Gambar 2c).
Analisis Sikuen DNAHasil analisis BLAST sikuen DNA gen
CP CMV dari semua lokasi menunjukkan
homologi yang tinggi sebesar 95–99% dengan CMV galur asal kedelai Bogor (FJ177303). Sikuen DNA sebagian gen CP sampel Ponorogo menunjukkan homologi tertinggi sebesar 90% dengan BCMV isolat kacang hijau asal Cina (KC832502) dan BCMV galur Blackeye asal kacang merah dari Vietnam (DQ925421). Sikuen DNA sampel dari Bogor Barat yang diamplifikasi dengan primer universal gen CP Geminivirus menunjukkan homologi tertinggi sebesar 96% dengan Pepper yellow leaf curl virus (PYLCV) isolat cabai asal Bogor (DQ083764) dan asal Jawa (JX416180).
63
Gambar 2 Pita DNA virus hasil amplifikasi dengan primer a, Cucumber mosaic virus (650 pb); b, universal Potyvirus (320 pb); c, universal Geminivirus (900 pb); 1, kontrol positif; 2, sampel Cikarawang; 3, sampel Bogor Barat; 4, sampel Cirebon; 5, sampel Bantul; 6, sampel Ponorogo; M, penanda DNA 1 kb (Thermo).
a
21 543 6
~650 pb
b
21 543 6
320 pb
c
21 543 6
~900 pb

J Fitopatol Indones Rahim et al.
Gambar 3 Pohon filogenetika virus kedelai asal Indonesia (dalam huruf tebal) terhadap virus yang sama dari negara lain. a, CMV; b, BCMV; dan c, PYLCV. PSV, Peanut stunt virus; PStV, Peanut stripe virus; TLCV, Tomato leaf curl virus; MLCV, Malvastrum leaf curl virus; AYV, Ageratum yellow vein virus; SPLCV, Sweet potato leaf curl virus; MYMV, Mungbean yellow mosaic virus; PSV-POL, SMV-C15 CHN, PSV-POL, MYMV MB-VIE sebagai pembanding di luar grup.
CMV-S IDN CkrwngCMV-S IDN Bgr Brt CMV-S IDN Crb CMV-S IDN Btl CMV-S IDN Png CMV-S IDN Bgr CMV-Ts IND (EF153734) CMV-Tfn ITA (Y16926)CMV-113 USA (AF523340)CMV-Lucknow IND (EF187825) CMV-3 BAU MAS (DQ195082) CMV TR15-THA (AJ810264) PSV-POL (JN135294)
83
95
95
a
BCMV-IDN Png BCMV-NL4 COL (DQ666332) BCMV-S KOR (KJ508092) PStV-115-IDN (AJ132158) BCMV-MB CHN (KC832502) BCMV-IRN (KF670145) BCMV-BIC VIE (DQ925421) BCMV-BIC USA (Y17823) BCMV-IND (FJ712783) SMV-C15 CHN (GQ491077)
89
b PYLCV-IDN Jawa PYLCV-IDN Bgr PYLCV-IDN Bogor Barat TLCV-IDN Sul (FJ237615) TLCV-KOM (AM701759) MLCV-CHN (KJ016236) TLCV-PHI (EU487042) AYV-TAI (DQ866133) SPLCSPV-RSA (JQ621844) SPLCV-CHN (NC_024693) MYMV-MB-VIE (JX244181)
100
99
c
64
Analisis Filogenetika Hasil analisis filogenetika berdasarkan
sikuen DNA menunjukkan CMV asal Cikarawang, Bogor Barat, Cirebon, Bantul, dan Ponorogo berada dalam kelompok yang sama dengan CMV-S asal Bogor, Indonesia yang terpisah dari CMV galur lainnya (Gambar 3a). BCMV asal Ponorogo berada pada kelompok yang terpisah dengan BCMV asal negara lain (Gambar 3b), sedangkan PYLCV asal Bogor Barat berada dalam kelompok yang sama dengan PYLCV isolat cabai asal Bogor dan Jawa (Gambar 3c).
PEMBAHASAN
Variasi gejala yang ditemukan di lapangan kemungkinan dapat disebabkan kultivar yang berbeda dan umur tanaman yang bervariasi saat pengambilan sampel dilakukan. Umur tanaman kedelai di lapangan berkisar 45–60 hari setelah tanam (HST). Gejala penyakit virus pada tanaman kedelai setelah melewati awal pertumbuhan (14–28 HST)
sulit dibedakan karena gejalanya menjadi kompleks dan bervariasi (Andayanie 2012).
Infeksi tunggal oleh CMV menghasilkan gejala penyakit yang lebih ringan dibanding-kan dengan gejala oleh infeksi Cowpea mottle virus (CMeV) (Arogundade et al. 2009). Pada infeksi campuran beberapa virus terjadi interaksi 2 virus atau lebih yang menginduksi gejala lebih parah (sinergis) atau sebaliknya (antagonis) (Syller 2012).
BCMV dan CMV dilaporkan menginfeksi kedelai, kacang hijau, dan kacang tanah di Jawa Barat (Green et al. 1988). Kedelai yang diinokulasi dengan BCMV menghasilkan gejala mosaik ringan yang tersebar di permukaan daun (Petrovic 2010). Berdasarkan gejala yang ditemukan di lapangan, sulit membedakan mosaik yang disebabkan CMV dan BCMV. Deteksi serologi atau asam nukleat sangat diperlukan untuk memastikan virus penyebabnya.
Hasil deteksi dan identifikasi molekuler menunjukkan CMV-S adalah virus yang dominan menginfeksi kedelai di semua

J Fitopatol Indones Rahim et al.
lokasi pengambilan sampel. Hal ini mungkin menunjukkan bahwa virus tersebut telah menyebar merata di semua lokasi pengambilan sampel. CMV-S dapat ditularkan melalui benih dan beberapa spesies kutudaun secara nonpersisten. Penularan CMV-S melalui benih cukup tinggi, yaitu 40–100% (Hartman et al. 1999). Sikuen nukleotida gen CP CMV yang berasal dari seluruh lokasi pengambilan sampel memiliki homologi yang tinggi terhadap CMV-S asal Bogor serta membentuk kelompok tersendiri yang terpisah dengan CMV galur nonlegum asal negara lain, namun masih termasuk dalam subgrup IB.
Homologi yang tinggi antarisolat CMV dari semua lokasi pengambilan sampel me-nunjukkan hampir tidak ditemukan keragaman genetika gen CP CMV-S. Perbedaan geografi kurang atau tidak berkontribusi terhadap keragaman genetika CMV-S. Hal ini diduga karena telah terjadi adaptasi inang SSV (CMV-S) terhadap kedelai lokal sehingga SSV (CMV-S) membentuk kelompok sendiri meskipun berbeda asal secara geografi (Hong et al. 2003).
Reaksi silang secara serologi dapat terjadi karena kedekatan homologi gen-gen tertentu suatu virus dalam genus yang sama. Gen CP SMV memiliki homologi yang sangat dekat dengan BCMV, namun terpisah jauh dengan Bean common mosaic necrosis virus (BCMNV) (Chen et al. 2003). Reaksi silang antara SMV dan BCMV diduga terjadi juga dalam penelitian ini. Sampel uji yang positif terdeteksi sebagai SMV dalam deteksi serologi, teramplifikasi hanya dengan primer universal Potyvirus. Analisis BLAST runutan DNA gen CP sampel tersebut menunjukkan homologi tertinggi dengan BCMV asal Cina dan Vietnam. Hal yang sama dilaporkan terjadi juga pada antibodi Henbane mosaic potyvirus (HMV) dan Potato Y potyvirus (PVY) yang bereaksi positif dengan antibodi Eggplant mottle virus (EMoV) (Bhat et al. 1999) dan Tomato spotted wilt virus (TSWV) bereaksi positif dengan Chrysanthemum stem necrosis virus (CSNV) (Boben et al. 2007).
Insidensi BCMV pada kedelai belum pernah dilaporkan sebelumnya di Indonesia,
walaupun kedelai dapat menjadi inang BCMV di Afrika (CABI 2007). BCMV isolat kedelai saat ini dilaporkan menjadi ancaman potensial produksi kedelai pada beberapa wilayah di Cina. Berdasarkan analisis filogenetika BCMV isolat kedelai dari Cina masuk dalam grup I terpisah dengan BCMV isolat dan galur lainnya. Hal ini menunjukkan adanya diferensiasi tetua dan adaptasi inang (Zhou et al. 2014). BCMV isolat kedelai asal Ponorogo juga berada pada kelompok yang terpisah dari galur BCMV atau isolat lainnya dalam filogenetika.
PYLCV (genus Begomovirus) yang terdeteksi pada sampel asal Bogor Barat memiliki hubungan kekerabatan yang sangat dekat dengan PYLCV isolat cabai asal Indonesia. Sebelumnya dilaporkan terdapat 2 Begomovirus yang menginfeksi kedelai secara alami di Nigeria, yaitu Soybean mild mottle virus (SbMMV) dan Soybean chlorotic blotch virus (SbCBV) (Alabi et al. 2010). Penularan PYLCV dengan kutukebul dalam percobaan rumah kaca menunjukkan bahwa tanaman yang termasuk famili Leguminosae (kedelai, kacang panjang, kacang hijau, orok-orok) dapat terinfeksi PYLCV walaupun intensitas serangannya lebih ringan dan masa inkubasinya relatif lebih lama (Sulandari et al. 2006). Di lapangan, pertanaman cabai, tomat, dan kacang-kacangan umumnya ditanam pada lahan yang sama sehingga kemungkinan terjadinya infeksi virus pada tanaman selain inang utama tidak dapat diabaikan. Infeksi alami PYCLV pada kedelai asal Bogor Barat merupakan informasi baru yang menunjukkan ekstensi inang dari virus ini selain Solanaceae.
Berdasarkan pada hasil deteksi tersebut di atas, CMV galur S masih merupakan virus yang dominan menginfeksi kedelai dengan insidensi yang paling tinggi dibandingkan dengan virus lainnya di beberapa pertanaman kedelai di Jawa. Insidensi BCMV dan PYCLV di lapangan merupakan suatu informasi baru.
DAFTAR PUSTAKA
Akin HM. 2003. Respon beberapa genotipe kedelai terhadap infeksi CPMMV
65

J Fitopatol Indones Rahim et al.
(Cowpea mild mottle virus). J HPT Trop. 3(2):40–43.
Alabi OJ, Kumar PL, Mgbechi-ezeri JU, Naidu RA. 2010. Two new ‘legumoviruses’ (genus Begomovirus) naturally infecting soybean in Nigeria. Arch Virol. 155:643–656. DOI: http://dx.doi.org/10.1007/s00705-010-0630-3.
Andayanie WR. 2012. Diagnosis penyakit mosaik (Soybean mosaic virus) terbawa benih kedelai. J HPT Trop. 12(2):185–191.
Anggraini S, Hidayat SH. 2014. Sensivitas metode serologi dan polymerase chain reaction untuk mendeteksi Bean common mosaic Potyvirus pada kacang panjang. J Fitopatol Indones. 10(1):17–22. DOI: http://dx.doi.org/10.14692/jfi.10.1.17.
Anjarsari L, Suastika G, Damayanti TA. 2013. Deteksi dan identifikasi Potyvirus pada ubi jalar di Tana Toraja, Sulawesi Selatan. J Fitopatol Indones. 9(6):193–201. DOI: http://dx.doi.org/10.14692/jfi.9.6.193.
Arogundade O, Balogun SO, Aliyu TH. 2009. Effect of Cowpea mottle virus and Cucumber mosaic virus on six soybean (Glycine max L.) kultivars. J Virol. 220(6):1–5. DOI: http://dx.doi.org/10.1186/1743-422X-6-220.
Asadi, Soemartono, Woerjono M., dan Jumanto, H. 2003. Kendali genetik ketahanan kedelai terhadap penyakit virus kerdil (Soybean stunt virus). Zuriat. 14(2):1–11.
Bhat AI, Varma A, Pappu HR, Rajamannar, Jain RK, Praveen S. 1999. Characterization of a potyvirus from eggplant (Solanum melongena) as a strain of potato virus Y by N-terminal serology and sequence relationships. Plant Pathol. 48:648–654. DOI: http://dx.doi.org/10.1046/j.1365-3059.1999.00384.x.
Boben J, Mehle N, Pirc M, Plesko IM, Ravnikar M. 2007. New molecular diagnostic methods for detection of Chrysanthemum stem necrosis virus (CSNV). ABS. 50(1):41–51.
[BPS] Badan Pusat Statistik. 2013. Data Statistik Indonesia. Badan Pusat Statistik.
Jakarta. www.bps.go.id [diakses 9 April 2013].
[CABI] Centre for Agriculture and Biosciences International. 2007. Crop Protection Compendium. (Serial Online). Wallingford (UK): CAB International.
Chen J, Zheng HY, Shi YH, Adams MJ, Antoniw JF, Zhao MF, Shang YF and Chen JP. 2003. A virus related to soybean mosaic virus from Pinellia ternata in China and its comparison with local soybean SMV isolates. Arch Virol. 149:349–363. DOI: http://dx.doi.org/10.1007/s00705-003-0184-8.
Doyle JJ, Doyle JL. 1987. A rapid DNA isolation procedure for small quantities of fresh leaf tissue. Phytochem Bull. 19:11–15.
Green SK, Lee DR, Horne NM. 1988. Survey for viruses of soyabean, mung bean and peanut in Java, Indonesia. Plant Dis. 72:994. DOI: http://dx.doi.org/10.1094/PD-72-0994A.
Grisoni M, Moles M, Farreyrol K, Rassaby L, Davis R, Pearson M. 2006. Identification of potyviruses infecting vanilla by direct sequencing of a short RT-PCR amplicon. Plant Pathol. 55:523–529. DOI: http://dx.doi.org/10.1111/j.1365-3059.2006.01397.x.
Hartman GL, Sinclair JB, Rupe JC. 1999. Compendium of Soybean Diseases. Ed ke-4. St. Paul, Minnesota (US): The American Phytopathological Society. hlm 182.
Hong JS, Masuta C, Nakano M, Abe J, Uyeda I. 2003. Adaptation of Cucumber mosaic virus soybean strains (SSVs) to cultivated and wild soybeans. Theor Appl Genet. 107:49–53. DOI: http://dx.doi.org/10.1007/s00122-003-1222-3
Kim YH, Kim OS, Roh JH, Moon JK, Sohn SI, Lee C, Lee JY. 2004. Identification of soybean mosaic virus strains by RT-PCR analysis of cylindrical inclusion coding region. Plant Dis. 88(6):641–644. DOI: http://dx.doi.org/10.1094/PDIS.2004.88.6.641.
Li R, Salih S, Hurt S. 2004. Detection of Geminiviruses in sweetpotato by
66

J Fitopatol Indones Rahim et al.
polymerase chain reaction. Plant Dis. 88(12):1347–1351. DOI: http://dx.doi.org/10.1094/PDIS.2004.88.12.1347.
Petrovic D, Ignjatov M, Nicolic Z, Vujakovic M, Vasic M, Milosevic M, Taski-Adjukovic K. 2010. Occurrence and distribution of viruses infecting the bean in Serbia. Arch Biol Sci Belgrade. 62(3):595–601. DOI: http://dx.doi.org/10.2298/ABS1003595P.
Sulandari S, Suseno R, Hidayat SH, Harjosudarmo J, Sosromarso S. 2006. Deteksi dan kajian kisaran inang virus penyebab penyakit daun keriting kuning cabai. Hayati J Biosci.13(1):1–6.
Syller J. 2012. Facilitative and antagonistic interactions between plant viruses in mixed infections. Mol Plant Pathol. 13(2):204–216. DOI: http://dx.doi.org/10.1111/j.1364-3703.2011.00734.x.
Zhou GC, Wu XY, Zhang YM, Wu P, Wu XZ, Liu LW, Wang Q, Hang YY, Yang JY, Shao ZQ, Wang B, Chen JQ. 2014. A genomic survey of thirty soybean-infecting Bean common mosaic virus (BCMV) isolates from China pointed BCMV as a potential threat to soybean production. Virus Res. 191:125–133. DOI: http://dx.doi.org/10.1016/j.virusres.2014.07.029.
67

Volume 11, Nomor 2, April 2015Halaman 68–72
DOI: 10.14692/jfi.11.2.68ISSN: 0215-7950
*Alamat penulis korespondensi: Jurusan Hama dan Penyakit Tumbuhan, Fakultas Pertanian, Universitas Andalas Padang, Kampus Limau Manis, Padang 25163.Tel: 0751-7059087, Faks: 0751-72702, surel: [email protected]
68
Keragaman Cendawan Rizosfer dan Potensinya sebagai Agens Antagonis Fusarium oxysporum
Penyebab Penyakit Layu Tanaman Krisan
Variability of Fungal Rhizosphere and Its Role as Antagonist Agents of Fusarium oxysporum Causing Wilt Disease in Crysanthenum sp.
Engla Yona Liza, Adrinal, Jumsu Trisno*Universitas Andalas, Padang 26163
ABSTRAK
Daerah rizosfer tanaman merupakan daerah yang kaya mikrob, salah satunya ialah kelompok cendawan. Penelitian ini bertujuan mengisolasi cendawan rizosfer dari tanaman krisan sehat dan menguji potensinya sebagai agens hayati untuk pengendalian Fusarium oxysporum penyebab penyakit layu. Isolasi cendawan rizosfer dilakukan dengan metode pengenceran berseri pada medium agar-agar dekstrosa kentang. Ada 6 isolat cendawan yang diperoleh dari rizosfer. Uji patogenisitas menunjukkan bahwa 6 isolat tersebut bukan patogen karena tidak menimbulkan gejala penyakit layu pada tanaman krisan. Hasil seleksi kemampuan antagonisnya menunjukkan bahwa 4 isolat mempunyai kemampuan daya hambat (46.42–51.61% ) dan antibiosis terhadap F. oxysporum. Isolat yang memiliki potensi agens hayati ini diidentifikasi sebagai Trichoderma sp. (2 isolat), Penicillium sp., dan Paecilomyces sp.
Kata kunci: agens hayati, antibiosis, uji patogenisitas
ABSTRACT
Plant rhizosphere is rich of microbes, one of these is a group of fungi. This research aimed to obtain fungi isolates from rhizosphere of healthy Chrysanthemum, followed by evaluation of their potential as biocontrol agents of Fusarium oxysporum. Isolation of fungi rhizosphere was based-on serial dilution method. As many as 6 isolates of rhizosphere fungi was succesfully isolated with diverse characters. Pathogenicity test showed that 6 isolates were not pathogenic because it did not cause wilt symptoms on Chrysanthemum. The selected isolates were subjected to in vitro assay for their potential as biocontrol agents, 4 isolates had antagonist ability (46.42–51.61%) and antibiosis activity against F. oxysporum. Fungi isolates having the potential as biocontrol agents was identified as Trichoderma sp. (2 isolates), Penicillium sp., and Paecillomyces sp.
Key words : antibiosis, biological agents, pathogenicity test
PENDAHULUAN
Krisan (Chrysanthemum sp.) merupakan tanaman hias yang sangat diminati sebagai bunga potong. Salah satu kendala dalam budi
daya tanaman krisan ialah penyakit layu yang disebabkan oleh Fusarium oxysporum.
Sejauh ini petani masih bergantung pada pestisida sintetik dalam mengendalikan penyakit tanaman. Oleh karena itu, penggunaan

J Fitopatol Indones Liza et al.
69
agens hayati lokal perlu dikembangkan, terutama untuk patogen tular tanah. Rizosfer merupakan bagian tanah yang berada di sekitar perakaran tanaman dan berperan sebagai pertahanan luar bagi tanaman terhadap serangan patogen akar. Populasi mikroorganisme di rizosfer biasanya lebih banyak dan beragam dibandingkan pada tanah bukan rizosfer. Purwantisari dan Rini (2009) melaporkan beberapa cendawan rizosfer pada tanaman kentang.
Eksplorasi dan pemanfaatan cendawan rizosfer untuk pengendalian penyakit pada tanaman krisan belum banyak dilaporkan. Penelitian ini bertujuan mendapatkan cendawan rizosfer yang dapat menekan perkembangan F. oxysporum penyebab penyakit layu tanaman krisan.
BAHAN DAN METODE
Sampel Tanah RizosferPengambilan sampel tanah rizosfer di-
lakukan dengan metode acak terpilih. Sampel diambil dari tanaman sehat di lahan endemik penyakit layu fusarium. Tanah diambil dengan menggunakan bor tanah pada 5 cm dari pangkal batang dengan kedalaman 10–15 cm. Pada setiap sampel tanaman diambil 3 titik sampel tanah dengan bobot masing-masing ± 20 g. Sampel tanah digabungkan sebagai contoh komposit.
Isolasi Cendawan RizosferCendawan diisolasi dari tanah rizosfer
tanaman krisan dengan metode pengenceran berseri sampai didapatkan pengenceran 10-5. Sebanyak 1 mL suspensi pada pengenceran 10-3, 10-4, dan 10-5 dituangkan pada medium agar-agar dekstrosa kentang (ADK) dan diinkubasi 48 jam. Setiap koloni yang tumbuh dengan ciri cendawan berbeda, diisolasi ke medium ADK baru sehingga didapatkan biakan murni. Semua cendawan yang telah murni dibuat kultur stok untuk uji lanjut.
Isolasi Cendawan Fusarium oxysporumF. oxysporum diisolasi dengan metode
moist chamber dari tanaman krisan yang
bergejala layu pada lokasi sampel cendawan rizosfir. Cendawan yang tumbuh diisolasi ke medium ADK dan diinkubasi 96 jam. Reisolasi dilakukan sampai didapatkan biakan murni F. oxysporum.
Uji PatogenisitasUji patogenisitas dilakukan untuk
cendawan F. oxysporum dan cendawan rizosfer. Pengujian dilakukan pada tanaman krisan berumur 3 bulan, dengan metode Handoko et al. (2014) dengan modifikasi inokulasi cendawan melalui pelukaan. Pangkal batang tanaman dengan jarak ± 0.5 cm dari permukaan tanah, dilukai membujur dengan panjang ± 1 cm dan kedalaman ± 2 mm. Bagian tanaman yang dilukai ditempeli potongan biakan cendawan rizosfer yang didapatkan, begitu juga dengan cendawan F. oxysporum. Tanaman diinkubasi selama 7 hari pada ruangan dengan suhu 25 °C (suhu kamar), sampai menunjukkan gejala. Apabila selama 7 hari tidak menunjukkan gejala, cendawan rizosfer tersebut tidak patogen.
Uji AntagonisUji kemampuan antagonis cendawan
rizosfer dilakukan dengan metode biakan ganda pada medium ADK. Cendawan rizosfer dan patogen dibiakkan dalam satu cawan petri (Gambar 1). Kemampuan antagonis ditentukan berdasarkan persentase kemampuan daya hambat dan antibiosis dengan menilai ada tidaknya zona hambatan. Persentase hambatan pertumbuhan cendawan dihitung berdasarkan rumus:
R= × 100%, denganr1 – r2
r2
A B
r2 r1
3cm
Gambar 1 Uji biakan ganda. A, Potongan koloni cendawan rizosfer; B, Potongan koloni cendawan Fusarium oxysporum.

J Fitopatol Indones Liza et al.
R, Persentase penghambatan pertumbuhan (%); r1, jari-jari F. oxysporum menjauhi isolat cendawan rizosfer; r2, jari-jari F. oxysporum mendekati isolat cendawan rizosfer.
IdentifikasiCendawan rizosfer yang mempunyai
kemampuan antagonis diidentifikasi sampai tingkat genus berdasarkan ciri morfologi makroskopi dan mikroskopi (Watanabe 2002).
HASIL
Ragam Cendawan Rizosfer Hasil isolasi cendawan dari rizosfer
tanaman krisan didapatkan 6 isolat cendawan dengan morfologi yang beragam (Tabel 1). Isolat tersebut tidak menunjukkan gejala (7 hari setelah inokulasi), sedangkan tanaman yang diinokulasi dengan cendawan F. oxysporum menunjukkan gejala layu.
Hasil identifikasi diketahui bahwa cendawan yang berhasil diisolasi dari rizosfer tanaman krisan ialah Penicillium, Trichoderma, Paecilomyces, dan 2 isolat
(isolat A dan D) tidak diidentifikasi karena tidak mempunyai kemampuan sebagai agens hayati.
Kemampuan AntagonisHasil pengujian kemampuan antagonis
menunjukkan bahwa 4 isolat cendawan rizosfer mempunyai kemampuan antagonis dengan persentase daya hambat dan kemampuan antibiosis yang beragam (Tabel 2; Gambar 2).
PEMBAHASAN
Cendawan rizosfer dari perakaran tanaman krisan sehat di lahan endemik penyakit layu menunjukkan keragaman yang tinggi. Dari satu rizosfer tanaman didapatkan 6 isolat cendawan dari genus Penicillium, Trichoderma, dan Paecilomyces. Purwantisari dan Rini (2009) melaporkan bahwa cendawan yang ada pada rizosfer beragam, penelitian tersebut mendapatkan 8 isolat cendawan antagonis dari rizosfer tanaman kentang, yaitu genus Trichoderma, Penicillium, Phytophthora, dan Mucor.
Karakter morfologi
Ciri-ciri isolat Isolat A Isolat B Isolat C Isolat D Isolat E Isolat F
Warna Koloni Cokelat Abu-abu kehijauan Hijau Oranye
bintik hitamAbu-abu kehijauan
Putih bintik kecokelatan
Bentuk Koloni Melingkar Menyebar Melingkar Melingkar Menyebar MelingkarKetebalan Koloni Tebal Tipis Tebal Tebal Tipis Sangat tebal
Pertumbuhan Koloni Cepat Cepat Cepat Lambat Cepat Lambat
Tabel 1 Karakter morfologi isolat cendawan rizosfer tanaman krisan
70
Cendawan rizosfer Daya hambat (%) Antibiosis*A (tidak diidentifikasi) 0.00 -B (Penicillium sp.) 51.51 +C (Trichoderma sp.) 51.61 -D (Tidak diidentifikasi) 0.00 -E (Penicillium sp.) 46.42 + F (Paecilomyces sp.) 50.00 +
*-, tidak memiliki kemampuan antagonis; +, memiliki kemampuan antagonis
Tabel 2 Kemampuan antagonis cendawan rizosfer tanaman krisan dalam menekan perkembangan cendawan Fusarium oxysporum penyebab penyakit layu tanaman krisan

J Fitopatol Indones Liza et al.
Cendawan rizosfer dari penelitian ini dapat menghambat F. oxysporum. Salah satu mekanisme cendawan antagonis sebagai agens pengendalian hayati penyakit tanaman ialah menghambat dengan cara kompetisi, parasitisme, antibiosis, dan lisis.
Penicillium sp. dapat bersifat antagonis terhadap patogen tular tanah. Cendawan ini telah dicoba sebagai mikrob antagonis terhadap Sclerotium rolfsii yang merupakan patogen tular tanah yang merusak lebih dari 500 spesies tanaman, namun daya hambatnya paling rendah dibandingkan dengan Trichoderma sp. dan Aspergillus sp. yakni sebesar 56.98–46.24% (Bosah et al. 2010). Penicillium sp. bersifat antagonis karena mengeluarkan beberapa senyawa alkaloid seperti agroklavine dan ergometrine yang memiliki sifat anticendawan terhadap Botrytis cinerea, F. solani, dan Alternaria tenius (Haggag dan Mohamed 2007). Penicillium sp.juga telah dicoba untuk mengendalikan penyakit lanas pada tanaman tembakau yang disebabkan oleh P. parasitica var. nicotianae (Roeswitawati 2007).
Genus Trichoderma sp. telah banyak diteliti sebagai agens hayati untuk mengendalikan berbagai penyakit tanaman. T. harzianum dan T. viride telah dikomersialkan untuk mengendalikan penyakit busuk akar dan penyakit rebah kecambah yang disebabkan oleh cendawan Fusarium, Rhizoctonia, dan Pythium (Alabouvette et al. 2006). Purwantisari dan Rini (2009) mengatakan bahwa Trichoderma sp. adalah salah satu cendawan antagonis spesifik lokasi yang menunjukkan kemampuannya dalam uji antagonisme secara in vitro dalam mengendalikan pertumbuhan cendawan patogen P. infestans penyebab penyakit busuk daun tanaman kentang.
DAFTAR PUSTAKA
Alabouvette C, Olivain C, Steinberg C. 2006. Biological control of plant diseases: the European situation. Eur J Plant Pathol. 114:329–341. DOI: http://dx.doi.org/10.1007/s10658-005-0233-0.
Bosah O, Igeleke CA, Omorusi VI. 2010. In vitro microbial control of pathogenic
71
Gambar 2 Daya hambat (antagonis) cendawan rizosfer tanaman krisan dalam menekan pertumbuhan cendawan Fusarium oxysporum (a). A, isolat A (belum diidentifikasi); B, Penicillium sp.; C, Trichoderma sp.; D, isolat D (belum diidentifikasi); E, Penicillium sp.; dan F, Paecilomyces sp.
A B C
a a a
D E F
a a a

J Fitopatol Indones Liza et al.
Sclerotium rolfsii. Int J Agric Biol. 12:474–476. DOI: http:// www.fspublishers.org/ 09–047/ AWB/2010/12–3–474–476.
Haggag WM, Mohamed HALA. 2007.Biotechnological aspects of microorganismsused in plant biological control. Word J Agric Sci. 3(6):771–776.
Handoko A, Abadi AL, Aini LQ. 2014. Karakterisasi penyakit penting pada pembibitan tanaman durian di Desa Plangkrongan Kabupaten Magetan dan pengendalian dengan bakteri antagonis secara in vitro. J HPT. 2(2):15–22.
Purwantisari S, Rini BH. 2009. Isolasi dan identifikasi cendawan indigenous rizosfer
tanaman kentang dari lahan pertanian kentang organik di Desa Pakis, Magelang. J Bioma. 11(2):45–53.
Roeswitawati D. 2007. Penggunaan inokulum antagonis (cendawan dan bakteri) dalam menekan penyakit lanas (Phytophthora parasitica var. nicotiana) pada tembakau. J Ilmu-Ilmu Pert Indones. 3:418–426.
Watanabe T. 2002. Pictorial Atlas of Soil and Seed Fungi: Morphologies of Cultured Fungi and Key to Species. Ed ke-2. Washington DC (US):CRC Press.
72


















