
Aggregation of marine sponge cells induced by Ca pulses, Ca ionophores, and phorbol esters proceeds...
8
Vol. 134, No. 3, 1986 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS February 13, 1986 Pages 1319-1326 AGGREGATION OF MARINESPONGE CELLS INDUCED BY Ca PULSES, Ca IONOPHORES, AND PHORBOL ESTERS PROCEEDS IN THE ABSENCE OF EXTERNAL Ca Philip Dunham1 and Gerald Weissmann 2 Marine Biological Laboratory, WoodsHole, MA 02543 Received December 17, 1985 SUMMARY. Cells of the sponge Microciona prolifera dissociated in Ca,Mg-free sea water reaggregate upon addition of Ca, an observation classically attri- buted to the requirement for Ca of a species-specific aggregation factor. We now report that, unexpectedly, extracellular Ca is not required during aggrega- tion; brief Ca pulses (l-3 set) terminated by excess EDTAsuffice to prepare the cells for aggregation by Ca ionophores (e.g. A23187). We also show that phorbol myristate acetate (PMA) promotes aggregation of pulse-prepared cells. Since PMA and A23187 act synergistically in Microciona, the "twin signal" hypo- thesis, signalling by Ca and protein kinase C in parallel, is validated in this primitive animal. @ 1986 Academic Press, Inc. Cells of the marine sponge Microciona prolifera, depleted of Ca by dis- sociation in Ca,Mg-free sea water, reaggregate when exposed to Ca (1, 2). The dependence on Ca of aggregation has been attributed to the properties of the species-specific Microciona aggregation factor (MAF), a lectin-like 2~10~ Mr proteoglycan (2-7) which requires Ca for its integrity in solution, and which also requires Ca to promote aggregation (7). We have recently suggested (8- 10) that ligand-induced movementsof intracellular Ca play a critical role in sponge cell aggregation, as in other examples of stimulus-response coupling We report here that, unexpectedly, extracellular Ca is not required over the course of aggregation: brief pulses (l-3 set) of Ca (lo-20 mM) terminated by excess EDTA (lo-40 mM) suffice to prepare cells for aggregation by the Ca ionophores A23187 and ionomycin. Ca therefore functions as an extracellular Permanent addresses: lDepartment of Biology, Syracuse University, Syracuse, NY 13210; 2Department of Medicine, NewYork University School of Medicine, New York, NY 10016. Abbreviations: factor; MAFf, CMFSW - Ca,Mg-free sea water; MAF - Microciona aggregation - MAF fragments; T - light transmission; PMA - phorbol myristate acetate; aoCa - external Ca activity. 0006-291X/86 $1.50 1319 Copyright 0 1986 by Academic Press, Inc. All rights qf‘ reproduction in any ,fbrm reserved.
-
Upload
philip-dunham -
Category
Documents
-
view
214 -
download
2
Transcript of Aggregation of marine sponge cells induced by Ca pulses, Ca ionophores, and phorbol esters proceeds...

Vol. 134, No. 3, 1986 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
February 13, 1986 Pages 1319-1326
AGGREGATION OF MARINE SPONGE CELLS INDUCED BY Ca PULSES, Ca IONOPHORES, AND PHORBOL ESTERS PROCEEDS IN THE ABSENCE OF EXTERNAL Ca
Philip Dunham1 and Gerald Weissmann 2
Marine Biological Laboratory, Woods Hole, MA 02543
Received December 17, 1985
SUMMARY. Cells of the sponge Microciona prolifera dissociated in Ca,Mg-free sea water reaggregate upon addition of Ca, an observation classically attri- buted to the requirement for Ca of a species-specific aggregation factor. We now report that, unexpectedly, extracellular Ca is not required during aggrega- tion; brief Ca pulses (l-3 set) terminated by excess EDTA suffice to prepare the cells for aggregation by Ca ionophores (e.g. A23187). We also show that phorbol myristate acetate (PMA) promotes aggregation of pulse-prepared cells. Since PMA and A23187 act synergistically in Microciona, the "twin signal" hypo- thesis, signalling by Ca and protein kinase C in parallel, is validated in this primitive animal. @ 1986 Academic Press, Inc.
Cells of the marine sponge Microciona prolifera, depleted of Ca by dis-
sociation in Ca,Mg-free sea water, reaggregate when exposed to Ca (1, 2). The
dependence on Ca of aggregation has been attributed to the properties of the
species-specific Microciona aggregation factor (MAF), a lectin-like 2~10~ Mr
proteoglycan (2-7) which requires Ca for its integrity in solution, and which
also requires Ca to promote aggregation (7). We have recently suggested (8-
10) that ligand-induced movements of intracellular Ca play a critical role in
sponge cell aggregation, as in other examples of stimulus-response coupling
We report here that, unexpectedly, extracellular Ca is not required over the
course of aggregation: brief pulses (l-3 set) of Ca (lo-20 mM) terminated by
excess EDTA (lo-40 mM) suffice to prepare cells for aggregation by the Ca
ionophores A23187 and ionomycin. Ca therefore functions as an extracellular
Permanent addresses: lDepartment of Biology, Syracuse University, Syracuse, NY 13210; 2Department of Medicine, New York University School of Medicine, New York, NY 10016. Abbreviations: factor; MAFf,
CMFSW - Ca,Mg-free sea water; MAF - Microciona aggregation - MAF fragments; T - light transmission; PMA - phorbol
myristate acetate; aoCa - external Ca activity.
0006-291X/86 $1.50
1319 Copyright 0 1986 by Academic Press, Inc.
All rights qf‘ reproduction in any ,fbrm reserved.

Vol. 134, No. 3, 1986 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
ligand. Moreover, since the aggregation of pulse-prepared cells is inhibited
by fragments of MAF, intact MAF must mediate aggregation. We also show
synergy between A23187 and phorbol myristate acetate (PMA, a tumor promoter
and an activator of protein kinase C) in promoting aggregation, validating the
"twin-signal" hypothesis of Nishizuka (11) in this primitive organism:
signalling in parallel in stimulus-response coupling by protein
kinase C and redistribution of intracellular Ca. A preliminary report of some
of these results has been published (12).
MATERIALS AND METHODS
Microciona prolifera was collected triweekly by the Department of Marine Resources, Marine Biological Laboratory, and kept in running sea water. sus- pensions of cells were prepared by mechanical dissociation in Ca,Mg-Free sea water (cMFSW) containing (mM): NaCl (460), Na2S04 (7), KC1 (lo), HEPES (lo), and EDTA (2.5), pH 7.8, and were kept at 4'C until use. Aggregation was measured as described before (8) from increases in light transmission (AT) in stirred (700 rpm) suspensions of cells (0.1 ml, s2 x lo7 cells) at room tem- perature in a Payton Aggregation Module model 600B (Payton Associates, Buff- alo NY; Model 1242 recorder, Soltec Corp., Sun Valley, CA); T was calibrated as described before (8).
MAF was prepared and its titer determined as described before (4, 5). Stock solutions (1:512 titer) were kept in CMFSW with 2 mM Ca and no EDTA. Ionomycin, PMA, phorbol, and phorbol dibutyrate were from Behring Diagnostics (La Jolla, CA); A23187 was from Sigma (St. Louis, MO).
RESULTS
Aggregation induced by Ca pulses. The extracellular role of Ca was demon-
strated by experiments such as those in Fig. lA, a recording of light trans-
mission (T) through a stirred suspension of sponge cells. At the times
indicated Ca, then EDTA, were added (final concentrations, 10 and 20 mM,
respectively). The time between the additions was 2 set (duration of the
pulse stimulus). No change in T followed. After 7 min the Ca ionophore
A23187 was added (to 10 PM); the marked increased in T is a measure of aggre-
gation, in this experiment proceeding in the presence of [EDTA] more than
double [Cal, and a°Ca <O.l mM.
In about 10% of 288 cell suspensions examined during this study, the cells
were characterized as "hyperresponsive," meaning the cells aggregated to the
same extent and at the same rate upon addition of Ca (constant exposure as did
most cells after addition of both Ca and A23187. Figure 1B shows aggregometer
recordings for hyperresponsive cells exposed to Ca pulses of four durations,
1320

Vol. 134, No. 3. 1986 8lOCHEMlCAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
A
t
plhr: 2 SW
20% Co e EDTA AT
H 1 min
I pulse: 5sac
Figure 1. Aggregation in response to calcium pulses of dissociated sponge cells, both normal and "hyperresponsive." Recordings are ahown of light transmission through stirred suspensions of cells. Fig. 1A: pulse-induced aggregation of normal cells. Ca was added to cells (final concentration, 10 mM), followed in 2 set by EDTA (to 2OmM). Seven minutes later A23187 was added (10 JIM). Stock solutions of A23187 were in CMSO. Here and elsewhere control cells received DMSO alone (final DMSO <1X). Fig. 1B: aggregation of "hyperresponsive" cells (defined in text). Recordings are shown for Ca pulses (Ca, 10 mM, followed by EDTA, 20 mM) of varying durations and also exposure to constant Ca (10 mM; stippled records). The 0 set pulse means simultaneous addition of Ca and EDTA.
along with recordings made at the same time of the same cells exposed to con-
stant Ca (stippled records). A 5 set pulse of Cs evoked aggregation indistin-
guishable from that induced by constant Ca. Pulses of two and one set caused
progressively less aggregation, and simultaneous addition of Ca and EDTA
caused none.
The results of a number of experiments like those in Fig. 1 are summarized
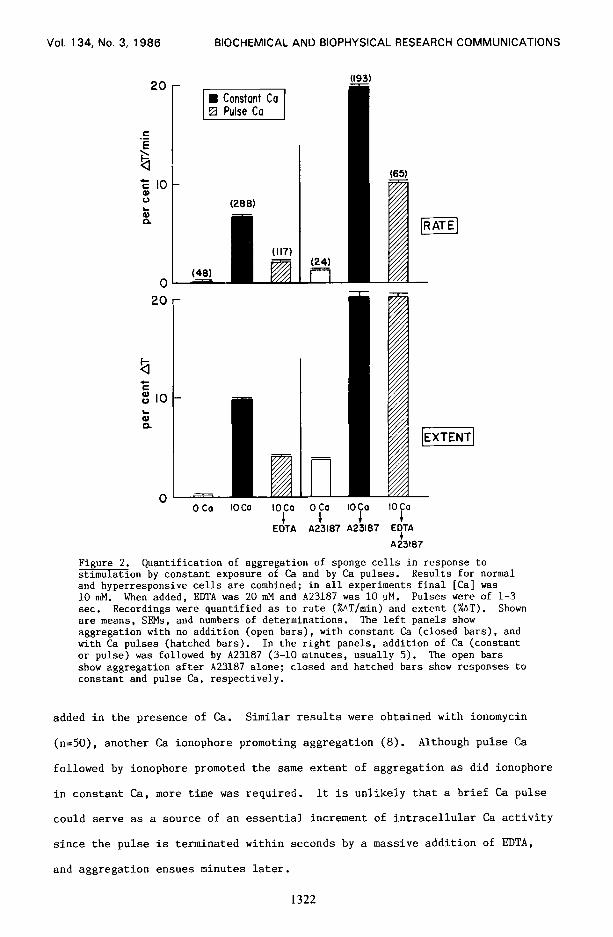
in Fig. 2. Aggregation was quantified as to rate (%AT/min) and extent (%AT).
The effect of pulses of Ca (l-3 set) can be compared with constant exposure to
Ca. In all of these experiments Ca was 10 mM and EDTA, when added, was 20 mM.
Pulses caused significant aggregation, but less than constant Ca did. A23187
added after a pulse was an effective aggregant, but not as effective as when
1321
Vol. 134, No. 3, 1986 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
0 20
0
(288)
Is: 0 co IO co loco
4
(24)
11 EOTA A23187 A23187 ‘4jTA
A23187
Figure 2. Quantification of aggregation of sponge cells in response to stimulation by constant exposure of Ca and by Ca pulses. Results for normal and hyperresponsive cells are combined; in all experiments final [Cal was 10 mM. When added, EDNA was 20 mM and A23187 was 10 uM. Pulses were of l-3 sec. Recordings were quantified as to rate (%AT/min) and extent (%AT). Shown are means, SEMs, and numbers of determinations. The left panels show aggregation with no addition (open bars), with constant Ca (closed bars), and with Ca pulses (hatched bars). In the right panels, addition of Ca (constant or pulse) was followed by A23187 (3-10 minutes, usually 5). The open bars show aggregation after A23187 alone; closed and hatched bars show responses to constant and pulse Ca, respectively.
added in the presence of Ca. Similar results were obtained with ionomycin
(n=50), another Ca ionophore promoting aggregation (8). Although pulse Ca
followed by ionophore promoted the same extent of aggregation as did ionophore
in constant Ca, more time was required. It is unlikely that a brief Ca pulse
could serve as a source of an essential increment of intracellular Ca activity
since the pulse is terminated within seconds by a massive addition of EDTA,
and aggregation ensues minutes later.
1322

Vol. 134, No. 3, 1986 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
Aggregation induced by A23187 alone. A23187 with no Ca caused much less
aggregation than Ca + A23187. However the mean aggregation in Fig. 2 caused
by A23187 alone was comparable to that caused by Ca pulses without A23187. We
examined this further in twelve paired experiments, comparing aggregation
after a Ca pulse and after A23187 with no Ca. The rate and extent of aggre-
gation were greater in two experiments with Ca pulses, the same in five, and
greater with ionophore in five experiments. The aggregation with ionophore at
10 uM without added Ca was probably due to redistribution of intracellular Ca.
In other experiments, A23187 at 1 UM enhanced Ca-induced aggregation (see
below), but caused none when added alone.
Effects of MAF fragments on pulse-induced aggregation. Since MAF requires
>l mM a°Ca for its integrity in solution, our results suggest that if MAF is
involved, its state or locus render it insensitive to EDTA. On the other
hand, it is possible that pulse-induced aggregation is anomalous in the sense
of being independent of MAF. We therefore exploited the inhibitory effect
of fragments of MAF, which form in EDTA, on MAF-promoted aggregation (3,
13). Fig. 3 shows the results of an experiment in which cells in paired
cuvettes were given a pulse of Ca (20 mM, then 40 mM EDTA, higher concentra-
tions than in the experiments in Fig. 2). MAF was added to one cuvette,
then A23187 to both. The normal pulse-induced aggregation was greatly
inhibited by the MAF fragments (MAFf,) formed in EDTA. Assuming specificity
of binding of the fragments [either to the cells (3) or to MAF (13)], the
results of this experiment show that pulse-promoted aggregation requires
MAF. Results of further experiments (legend, Fig. 3) confirm and extend
those presented graphically. In four experiments (row l), MAFf, inhibited
by more than 50% aggregation induced by Ca pulses and A23187. In three
experiments (row 2), MAFfr also inhibited aggregation promoted by a constant
Ca stimulus. It was important to show that the preparation of MAF employed
was active in promoting aggregation with Ca present. In seven experiments
the rate of aggregation was enhanced 1.9-fold, and extent 1.5-fold, by MAF
(26 units/ml) relative to Ca alone when added 1 min after Ca (20 mM).
1323

Vol. 134, No. 3, 1986 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
A
40% AT
CO PULSE
1 I CMFSW
H I min
Inhibition of sponge cell aggregation by Microciona aggregation magmenta (MAFfr). Two cell suspensions were given pulse Ca stimuli as indicated (Ca to 20 mM followed in 5 set by EDTA to 40' r&f). After 1 min, control cells (upper trace) were given CMFSW, and cells represented by lower trace were given MAF (final MAF, 26 units/ml). MAFfr form quickly with EDTA in excess of Ca (ref. 3). After 2 min. A23187 (to 10 uM) was added to both cuvettes. Confirmatory results (same [Cal and [A23187]):
aggregation:
protocol % inhibition by MAFfr
rate extent (n>
1. MAFf, inhibit after f
Ca- EDTA- 76 f 11 55 f 9 (4)
pulse stimulus 1 NFfr -A23187
2. MAFfr inhibit after I
EDTA- fMAFfr- 38 f 8 25 f 3 (3)
constant stimulus 1 Ca-MAF
Protocol indicates orders of additions. Row 1: EDTA (to 40 mM) 5 set after -- Ca; MAF 1 min later (forming MAFfr); A23187 2.5 min after MAF. Row 2: -- Initial EDTA 10 mM, then MAF (forming MAFf,); after 10 min, CZa, and then second MAF addition (remaining intact) 1 min. later.
Effects of phorbol esters on aggregation. Protein kinase C often has a role
in stimulus-coupling (11). Since it acts synergistically with Ca to promote
cellular functions, and is activated directly by tumor promoters such as PMA
(11). we tested PMA on aggregation. In cell suspensions preincubated with
0.1 to 5 uM PMA, aggregation was greatly enhanced either with a constant Ca
stimulus OF with pulse Ca + A23187 (Fig. 4). PMA alone had little effect;
Ca + A23187 (1 M) caused measureable but slight aggregation. In contrast
A23187 following PMA and either pulse or constant Ca provoked massive aggre-
gation, suggesting synergy of action between protein kinase C and intra-
cellular Ca. In four additional experiments, phorbol itself was without
effect, whereas phorbol dibutyrate was as effective as PMA.
1324

Vol. 134, No. 3. 1986 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
PMA co A23187 i
Figure 4. Promotion of sponge cell aggregation by phorbol myristate acetate (PMA). In each of these two experiments (constant and pulse Ca), PMA (to 100 nM, DMSO stock solution) was added to two of three cuvettes of cells (traces 1 & 3). After 2 min Ce was added to all 3 cuvettes (to 10 mM). In the pulse Ca experiment, EDTA (to 20 mM) was added 4 set later. After 3 more min A23187 (to 1 PM) was added to cuvettes 1 & 2 in both experiments. Thus aggregation caused by PMA alone (traces 3), A23187 alone (traces 2), and PMA + A23187 (traces 1) was observed with constant or pulse stimuli. Similar results were obtained in 10 such experiments; PMA was active at concentrations as low as 10 nM.
DISCUSSION
That aggregation proceeds without Ca is unexpected. MAF readily, and
irreversibly, dissociates in EDTA (5, 7), and MAF added in solution aggrega-
gates cells only in the presence of Ca (2, 7). In consequence, if MAF is
the physiological stimulus to which aggregation is a response (2, 8), MAF
after its secretion must reside at sites on the cell insensitive to EDTA, at
which sites it can promote aggregation. Indeed the natural aggregation of
sponge cells undoubtedly depends on surface bound MAF, and not MAF in solu-
tion. Is MAF the mediator of aggregation in the presence of excess EDTA?
Fragments of MAF are known to inhibit MAF-induced aggregation. MAFfr pro-
duced by heating with urea appear to inhibit by binding to the cells (3),
while a purified 10,000 M, subunit of MAF inhibits by binding to MAF (13;
this binding seems to require Ca). In either case, our results (Fig. 3)
1325

Vol. 134, No. 3, 1986 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
support the view that secreted MAF is the mediator of aggregation. Further-
more it follows that Ca, briefly presented extracellularly, acts as a
ligand, indirectly promoting secretion of MAF inaccessible to EDTA.
The synergy between Ca ionophore and PMA was not unexpected. Indeed the
data at once reinforce our hypothesis that aggregation of sponge cells is an
example of stimulus-response coupling and also the "twin-signal" hypothesis of
Nishizuka (II), which invokes the synergistic effects of intracellular Ca and
protein kinase C to explain cellular responses to external ligands. The
aggregation of sponge cells is the most primitive example, with respect to
phylogeny, yet advanced of the twin signal effect. Our observation of an
extracellular role for Ca as a ligand in stimulating sponge cell aggregation
may have broader significance in stimulus-secretion coupling of higher organ-
isms, particularly in cases where roles for cellular and external Ca have not
been distinguished.
ACKNOWLEDGEMENTS. We thank Leslie Vosshall, Cynthia Bayer, Lenore Azaroff, and Sarah Davidson for their competent technical assistance, and Dr. Tom Humphreys for useful discussions and for the MAF. This work was supported by NIH grants (AM-29290 & AM-33640 to PD; AM-11949 & HL-19721 to GW).
REFERENCES
1.
::
4.
5.
6.
7. 8.
9.
10.
11. 12.
13.
Galtsoff, P. S. (1925) J. Exp. Zool. 42, 183-221. Humphreys, T. (1963) Dev. Biol. 8, 27-47. Misevic, G. N., Jumblatt, J. E., and Burger, M. M. (1982) J. Biol. Chem. 257, 6931-6936. Humphreys, S., Humphreys, T., and Sano, J. (1977) J. Supramolec. Strut. 7, 339-351. Cauldwell, C. B., Henkart, P., and Humphreys, T. (1973) Biochem. 12, 3051-3055. Jumblatt, J. E., Schlup, V., and Burger, M. M. (1980) Biochem. 19, 1038-1042. Rice, D. J. and Humphreys, T. (1983) J. Biol. Chem. 258, 6394-6399. Dunham, P., Anderson, C., Rich, A. M., and Weissmann, G. (1983) Proc. Nat. Acad. Sci. 80, 4756-4760. Rich, A. M., Weissmann, G., Anderson, C., Vosshall, L., Haines, K. A., Humphreys, T., and Dunham, P. (1984) Biochem. Biophys. Res.Comm. 121, 863-870. Weissmann, G., Rich, A. M., Anderson, C., Haines, K. A., Korchak; H. M., and Dunham, P. (1984) Trans. Assoc. Am. Phys. 97, 369-383. Nishizuka, Y. (1984) Nature (London) 308, 693-695. Dunham, P. B., Vosshall, L. B., Bayer, C. A., Rich, A. M., and Weissmann, G. (1985) Fed. Proc. 44, 2914-2924. Verakalasa, P. and Humphreys, T. (1982) Biol. Bull. 163, 378 (abstr.).
1326


















