
Action of electrolytes on lateral body wall movements of grasshopper embryos (Acrididae, Orthoptera)
28
ACTION OF ELECTROLYTES ON LATERAL BODY WALL MOVEMENTS OF GRASSHOPPER EM- BRYOS (ACRIDIDAE, 0RTHOPTERA)l VINCENT THOMPSON ZooTogical Laboratory, Slate University of Iowa SIXTEEN FIBURES INTRODUCTION Since the demonstration by Ringer (1882) of the ions neces- sary for maintenance of normal contractions in the frog's heart, investigations of the effect of electrolytes on various types of movement have been widely extended. Reviews of the earlier literature on this subject have been prepared by Loeb ( '12), Falk ( '23), Bayliss ( '24) and Hober ( '26). Relatively few studies have been reported concerning the effect of ions on embryonic movements. Lewis ( '29) investi- gated the effect of various salts on the pulsation rate of isolated hearts from young chick embryos. Koller ( '32) transplanted embryonic frog hearts, and noted effects of alteration in salt contents of the medium. Yamamoto ('34) and his students have examined the movements of the embryo of the fish, Oryzias latipes, and varied the ionic content of the normal medium by use of artificial sea water. Embryos of the grasshopper, Melanoplus differentialis, during the process of yolk engulfment, exhibit vigorous, rhythmic lateral body wall movements, and Slifer ('34) has shown that these con- tractions in isolated embryos are satisfactorily maintained in a modified Ringer solution. 'Aided by a grant from the Rockefeller Foundation for research in cellular physiology. 19
-
Upload
vincent-thompson -
Category
Documents
-
view
213 -
download
0
Transcript of Action of electrolytes on lateral body wall movements of grasshopper embryos (Acrididae, Orthoptera)

ACTION O F ELECTROLYTES ON LATERAL BODY WALL MOVEMENTS OF GRASSHOPPER EM-
BRYOS (ACRIDIDAE, 0RTHOPTERA)l
VINCENT THOMPSON ZooTogical Laboratory, Slate University of Iowa
SIXTEEN FIBURES
INTRODUCTION
Since the demonstration by Ringer (1882) of the ions neces- sary for maintenance of normal contractions in the frog's heart, investigations of the effect of electrolytes on various types of movement have been widely extended. Reviews of the earlier literature on this subject have been prepared by Loeb ( '12), Falk ( '23), Bayliss ( '24) and Hober ( '26).
Relatively few studies have been reported concerning the effect of ions on embryonic movements. Lewis ( '29) investi- gated the effect of various salts on the pulsation rate of isolated hearts from young chick embryos. Koller ( '32) transplanted embryonic frog hearts, and noted effects of alteration in salt contents of the medium. Yamamoto ('34) and his students have examined the movements of the embryo of the fish, Oryzias latipes, and varied the ionic content of the normal medium by use of artificial sea water. Embryos of the grasshopper, Melanoplus differentialis, during the process of yolk engulfment, exhibit vigorous, rhythmic lateral body wall movements, and Slifer ('34) has shown that these con- tractions in isolated embryos are satisfactorily maintained in a modified Ringer solution.
'Aided by a grant from the Rockefeller Foundation for research in cellular physiology.
19

20 VINCENT THOMPSON
These movements in the grasshopper embryo were observed by' Nelsen ( '31) who suggested that they are resolved into the beatings of the heart when that organ is formed. This has been confirmed by Slifer ( '33 a ) and Thompson ( '37). Fur- ther, such activity is an unusually interesting type, as no completely differentiated striated muscle cells are present at the time movement begins (Slifer, '34).
This investigation of the effect of ions on lateral body wall movements was initiated since comparatively little work has been done on the action of electrolytes on embryonic move- ments, and, as observed above, grasshopper embryos are of special interest for such an inquiry.
MATERIALS AND METHODS
Embryos of the grasshopper, Melanoplus differentialis, were used throughout this study. Observations concerning the adaptability of this material for physiological experi- mentation and directions regarding collection and care of in- sects and eggs may be found in the papers of Carothers ('23, '37), Bodine ('25) and Slifer ('32 b). The stage used was that reached about 2 days after blastokinesis or 5 to 6 days after diapause. This was approximately the same age em- ployed by Slifer ( '34) and Walker ( '35), who made use of the lateral wall beatings for physiological indicators of solution isotonicity and carbon dioxide effects respectively. The darkly pigmented eyes, which may be seen through the egg shells, have reached a point midway between the two ends of the egg, making selection of eggs with embryos at the cotrect stage an easy task.
The embryos were dissected out of the eggs in a Ringer solution, as modified by Belar ('29), which consisted of 0.9% NaCl, and 0.02% each of potassium and calcium chlorides. A buffer was added, either 0.027. NaHCO, or phosphate buffer mixture, M/150 phosphate concentration, to hold the saline at about pH 6.3. Belar ('29) demonstrated the solu- tion to be isotonic with the germ cells of the grasshopper, and Slifer ('34) showed it to be satisfactory for the isolated embryos.

ELECTROLYTES AND EMBRYONIC MOVEMENT 21
The embryos were removed from the eggs by cutting away the end of the egg opposite the micropyle, and then, by gentle pressure, were forced out of the egg shells into the Ringer solution. Excess yolk, projecting beyond the eyes, was care- fully dissected away. Transference of the embryos from one solution to another was accomplished with a medicine dropper. In each group from ten to twenty-five embryos were used; these were passed through two washings in the solution to be tested and then arranged in order in a castor dish. The tem- perature was kept at 25 t 0.5"C. Observations were made with a binocular microscope, and a stop watch employed to determine the rate of movement.
The solutions were prepared from Bakers ' analyzed chemi- cals in doubly distilled water. The amounts of various salts required were calculated from freezing point data in the International Critical Tables. All solutions, which could be heated without loss of dissolved materials, were sterilized for 30 minutes in a steam autoclave. Glassware was washed with soap and water, rinsed free of soap, washed with dichromate- sulphuric acid cleaning solution, with tap water, and finally with distilled water.
The method of presentation of results is shown graphically in figure 1. The solid lines (fig. la) represent the per cent of embryos which continue movement after the indicated hours in the Ringer solution. Movement is easily maintained for 10 hours, and, with special care to avoid bacterial contamina- tion, this time may be extended several days. The results obtained with a solution which inhibits movement are also shown (fig. lb). This MgC1,-KC1-dextrose mixture will be discussed later, but is shown here because of the extreme changes which occur (i.e., a marked initial arrest of movement during the first 2 hours, followed by a slower recovery which varied from 100% in group (a) to about 70% in group (d), and, finally, a gradual decline in the number of active indi- viduals). The general uniformity of the behavior of the dif- ferent groups should be noted.

22 VINCENT THOMPSON
I I
Fig. 1 Behavior of embryos in Ringer and in a KCl-MgCI,-dextrose mixture. l a , effect of Ringer solution on lateral wall movements. - , per cent of embryns which continue moving at the indicated time. ---- , the average rate of move- ment in contractions/minute. lb, effect of a solution containing 4 parts isotonic KCl plus 4 parts MgCl, plus 2 parts dextrose on duration of movement. a'-d', curves from different groups of twenty-five embryos each. lc, effect of the same mixture on ra te of movement. a'-d', the average rate of contraction of those embryos in each group which are in motion at the designated times. Id, as Zc, except showing the average rate of movement of only those embryos which con- tinue to beat throughout the entire experiment.
TABLE 1 Statistical treatment of rate of lateral wall movement
Tima 0 1 hour 2 hours 3 hours 4 hours 6 hours
10 hours
Ringer Y P.E.
12.920.19 12.2fO.l.i 12.7f0.33 12.0+0.25 12.820.25 14.1f0.28 14.420.27
KCl .YgCl~~duztroae Timu Ai P.E. 0 13.220.23
30 minutes 22.820.33 1 hour 27.820.33 2 hours 27.720.57 3 hours 25.820.66 4 hours 24.420.64 5 hours 20.9k0.55 6 hours 19.520.55 7 hours 17.1+0.50 9 hours 15.7+0.34
The mean ra te of rontraction ( M ) and the probable error of the mean (P.E.) for movements of embryos immersed in Ringer solution and a KCI-MgCl,-dextroae mixture fo r the indicated times: 200 embryos were used in the Ringer; 100 in the latter solution.

ELECTROLYTES AND EMBRYONIC MOVEMENT 23
The rate of movement remains practically unchanged in Ringer (fig. la , dotted line), except for a slight increase toward the end of the 10-hour period, which is barely signifi- cant (table 1). I n the MgC1,-KC1-dextrose mixture, the rate varies greatly (fig. l c ) . Each curve represents the average rate of the embryos of that group which are still moving at the indicated times. These changes in rate are statistically significant (table 1). In general appearance, the curves are the same if one determines the average rate of only those embryos which remain in motion throughout the entire course of the experiment (fig. la).
To avoid confusion, a single line has been plotted in all figures, representing the mean of at least four groups of em- bryos (or a minimal total of sixty individuals) whieh were tested in the solution in question, and treated as above.
RESULTS
Non-el ectrolytes I n order to eliminate the effects of the presence of extra
ions, when testing an otherwise hypotonic concentration of a particular salt, it was necessary to find some non-electrolyte which could be present in considerable amounts without harm- ful results. Sucrose, dextrose, glycerol and urea were tested. Isotonic solutions of non-electrolytes are approximately 0.3 11, and the inhibitory effect of all these may be seen from exami- nation of figure 2a. The rapidity of stoppage is in the order:
Urea > glycerol > dextrose > sucrose
Carlson ('06) reported the same order for rate of inhibition of Limulus hearts, while Pantin ('31) obtained a reversed order for arrest of amoeboid movement. Certain amounts of salts in the fluid environment are essential for maintenance of proper functioning. This has been demonstrated in a wide variety of rhythmic systems, as turtle hearts and heart strips (Baetjer and McDonald, '31 ; Greene, 1899) ; Fundulus hearts (Carlson, '06) ; amoebae (Pantin, '31) ; frog skeletal muscle (Hober, '26) ; cilia (Gray, '28), and others.

24 VINCENT THOMFSON
Urea and glycerol were ineffective osmotically. I n these solutioiis the embryos behaved as if in distilled water, be- coming greatly swollen and whitish in appearance. Urea itself is not toxic to the lateral wall movements, since, with sufficient salt to maintain the osmotic pressure, movement con- tinues in the presence of the penetrating urea (fig. 2b). Hill
l t 3 4 S C - r HRS.
I- s I b
Fig. 2 Effect of non-electrolytea on lateral wall movement. - , per cent of embryos maintaining movement. - - - - , average rate in contractions/minute. 2a, 1-isotonic urea; 2-isotonic glycerol ; 3-isotonic dextrose; 4 isotonic sucrose ; 5-Ringer. 2b, 1-isotonic urea ; %urea plus Ringer. For further explanations see text.
( '32) found the same result with luminescence of Bacillus fischeri. On the other hand, Pantin ('31) found the most satisfactory non-electrolyte for amoeboid movement was urea, and Lussana ('11) employed it for replacing a large part of NaCl in Ringer for the frog's heart.
Of the two sugar solutions, dextrose proved the more satis- 'factory. The arrest of movement and decrease in rate dis- appeared with a mixture of isotonic solutions contaiiiing 20

ELECTROLYTES AND EMBRYONIC MOVEMENT 25
parts (or more) of Ringer with enough dextrose to make 100 parts. With sucrose, embryos were shrunken and movement was stopped in 10 parts (or more) of isotonic sugar plus Ringer to make 100 parts. Accordingly dextrose was selected as the non-electrolyte most suitable for use as a substitute for salts.
Cations 1. Essential cations in Ringer. Three main salts are pres-
ent in Ringer solutions: sodium, calcium and potassium chlo- rides. From the above experiments it is evident that a large
l . . . . . , . . . I I I I , + 3 C 1 8 ,
**>. % L J * 1 L T D . ) H.5
Fig.3 Effect of the components of Ringer on movement. - , per cent of embryos maintaining movement. - - - - , average rate in contraetions/minute. 3a, isotonic NaCI. 3b, NaCl plus KCI in the same proportions as in Ringer. 3e, NaCl plus CaCI, in the same proportions as in Ringer. 3d, normal Ringer (Belar) solution.
part of the NaCl serves merely to keep the osmotic pressure high enough: it may be replaced by a non-penetrating non- electrolyte as dextrose. The following experiment was de- vised to determine whether all three of these cations are essential for maintenance of contraction and normal rhythm rate. Embryos were placed 1) in isotonic NaC1, 2) in isotonic NaCl plus CaCl,, and 3) in isotonic NaCl plus KC1, with the CaCl, or KC1 at the same concentration as in normal Ringer (i.e., 0.0018 &I CaC1, or 0.0026 M KCI). Pure NaCl (fig. 3a)

36 VINCENT THOMPSON
does not permit movement in all the embryos, and about the same effect is seen when KC1 is added (fig. 3b). I n both there is some acceleration of rate, with the latter exhibiting about 100% increase. Embryos in NaCl plus CaC1, and in Ringer (fig. 3c and 3d) behave almost identically. Apparently the sodium and calcium chlorides are necessary, and the potas- sium chloride may be omitted from the solution. In general, pure NaCl solutions are toxic (Loeb, 'OO), and addition of calcium antagonizes this action. Potassium ions, it has been
"R3.
Fig.4 Effect of variation of pH. Per cent movement (solid lines) and aver- age rate in contractions/minute (broken lines) plotted against the length of time (hours) in Ringer a t the indicated pH. For further details see text.
suggested, are added to antagonize the possible effects of the calcium or part of the sodium ions (Chao, '33), but they are apparently not needed here.
The desired hydrogen ion concentration was attained by adding HC1 or NaOH to un- buffered Ringer, and then adding 1 part of glycine buffer mixture to 9 parts of the prepared Ringer., Regardless of the hydrogen ion concentration, no change was observed in rate of lateral body wall contractions (fig. 4). Movement con-
2. Efect o f variat,iolz o f p H .
' Glycine, in this concentration, is not harmful to movements of the grasshopper The pH of all solutions was checked by the quinhydrone or glass elec- embryo.
trode method. Calcium chloride was omitted from solutions above pH 8.

ELECTROLYTES AND EMBRYONIC MOVEMENT 27
tinued without arrest from pH 3.8 to 10.6. Above pH 10.6 movement was quickly inhibited. The embryos became soft and ‘mushy,’ sticking to any object they touched, and rapidly cytolyzing. This sudden effect at pH 10.6 is probably due to a complete break-down of cellular permeability : the embryos are greatly swollen and may be stretched to many times their original length. The swelling and ‘mushy’ appearance com- mences to be apparent at pH 9.0 to 9.5, but without harmful effects on the beatings of the lateral body walls.
In acid solutions no such sudden arrest of movement oc- curred. At pH 3.0 activity was completely depressed in about 6 hours, and at pH 2.0 an hour was required for 100% stop- page. I n these solutions the surface of the embryo turned whitish, giving the appearance of fixation, long before move- ment was discontinued.
This wide range of pH (3.0 to 3.7 to 10.6) over which grass- hopper embryos maintain movement is matched only by fig- ures fromwhole intact animals (Witschi, ’36). In most cases the alkaline limit for movement has been found to be from pH 9 to 10, while the acid limit varies more widely. For ex- ample, the range for movement of the crayfish heart (Linde- man, ’28) was pH 4 to 9 ; for activity in the isolated rabbit intestine (Fraser, ’25), pH 4.6 on the acid side; the Limulus heart-nerve preparation (Chaa, ’35), pH 5.2 to 10; amoeboid movement (Pantin, ’26), pH 6.0 to 9.0; ciliary movement (Gray, ’28), pH 6.4 to 9.2, and beating of the frog’s heart (Clark, ’27), pH 7 to 9. The relative insensitivity of lateral wall movements to changes in hydrogen ion concentration of the external medium indicates a degree of independence not equalled by other contractile systems. This, therefore, sug- gests difficulties in interpreting the ionic effects on these em- bryonic movements in the light of results from other types of activity. Nevertheless, such an indifference offers possibili- ties in testing certain conditions which ordinarily yield onlp a hydrogen ion effect. It has been possible, for example, to determine the effects of acetate solutions over a wider pH range than would be possible in other rhythmic systems.
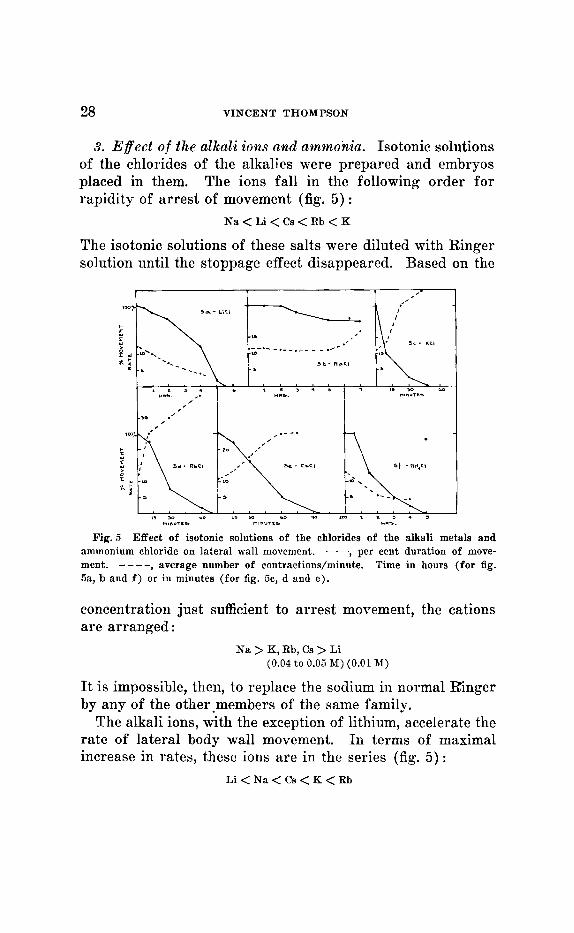
28 VINCENT THOMPSON
3. Efect o f the alkali ions and ammonia. Isotonic solutions of the chlorides of the alkalies were prepared and embryos placed in them. The ions fall in the following order for rapidity of arrest of movement (fig. 5) :
N a < Li < Cs < Rb < K
The isotonic solutions of these salts were diluted with Ringer solution until the stoppage effect disappeared. Based on the
ll,""Tr- rnl""TL3 " -5 .
Fig. 5 Effect of isotonic solutions of the chlorides of the alkali metals and ammonium chloride on lateral wall movement. -, per cent duration of move- ment. ---- , average number of contractions/minute. Time in hours (for fig. 5a, b and f ) or in minutes (for fig. Cic, d and e ) .
concentration just sufficient to arrest movement, the cations are arranged :
Na > K, Rb, Cs > Li (0.04 to 0.09 M ) (0.01 M )
It is impossible, then, to replace the sodium in normal Ringer by any of the other.members of the same family.
The alkali ions, with the exception of lit,hium, accelerate the rate of lateral body wall movement. In terms of maximal increase in rates, these ions are in the series (fig. 5 ) :
Li < N a < Cs < K < Rb

ELECTROLYTES AND EMBRYONIC MOVEMENT 29
If embryos be placed in 4 parts isotonic solutions of KC1 or RbC,l plus 6 parts Ringer (giving 0.06 hI KC1 or RbCl), arrest of movement with marked acceleration occurs (fig. 6a). Two parts of either salt solution plus 8 parts Ringer (i.e., 0.03M KC1 or RbCl) are without effect (fig. 6b). The addi- tive action of these two ions is shown by exposing the em- bryos to a mixture of 2 parts KC1 plus 2 parts RbCl plus enough Ringer to make 10 parts (fig. 6c). Here the salts are present individually in concentrations below that at which any effect is seen. The response of the lateral wall movements
n(R5.
Fig.6 Additive actions of K and Rb. - , per cent maintenance of move- ment. ---- , rate of movement in eontractions/minute. The following mixtures of isotonic solutions were used: 6a, 4 parts KC1 (or RbCl) plus 6 parts Ringer. 6b, 2 parts KC1 (or RbCl) plus 8 parts Ringer. 6c, 2 parts KCI plus 2 parts RbCl plus 6 parts Ringer.
is, however, as to the total concentration of the two ions in question. Further evidence concerning the similarity of the action of potassium and rubidium was found in the antago- nism of the effects of these ions by calcium (see below).
I n general the action of potassium and rubidium place them is a group apart from the other elements in this family. This has been reported for such different systems as frog muscle (Hober, '26), sperm (Gellhorn, '27), hearts (Clark, '27), and fish embryos (Yamamoto, '34). I n amoeboid (Pantin, '26) and ciliary (Gray, '28) movements the distinction between the effects of potassium and those of sodium are not so striking.
THE JOURNAL OF EXPERIMENTAL ZOOLOaY, YOL. 78, NO. 1

30 VINCENT THOMPSON
r
-
7 1
% 2 > * 3 H B .
man[ KCI I D C I T R O I L I loo*- ; -
I
I-
g ;; b t -
I I I I I
I-;-*
-
30 VINCENT THOMPSON
Lateral body wall contractions of grasshopper embryos fall in the first group.
Recovery in Ringer solution after exposure to excess KC1 was complete (fig. 7a). The rate of movement, at resumption of contraction by the lateral walls of the embryo, is not rapid as it was when arrest occurred. A non-electrolyte solution (dextrose) is less satisfactory than Ringer (fig. 7b). This
Fig. 7 Recovery of activity after arrest of movement in KCI. -, per cent of total embryos in motion. ---- , rate in contraetions/mjnute. The sequence of solutions is indicated on the graph. 7a, recovery in Ringer (Belar). 7b, recovery in dextrose.
recovery from potassium treatment is consistent with the re- sults of other workers (Martin, 04, on the terrapin heart; Thorner, '29, on the Fundulus heart; Matsura, '34, on move- ments in Oryzias embryos).
No alteration in the time of maintenance of movement in KCl or of recovery time after KC1 treatment could be found in the grasshopper embryos by variation in pH (from pH 4
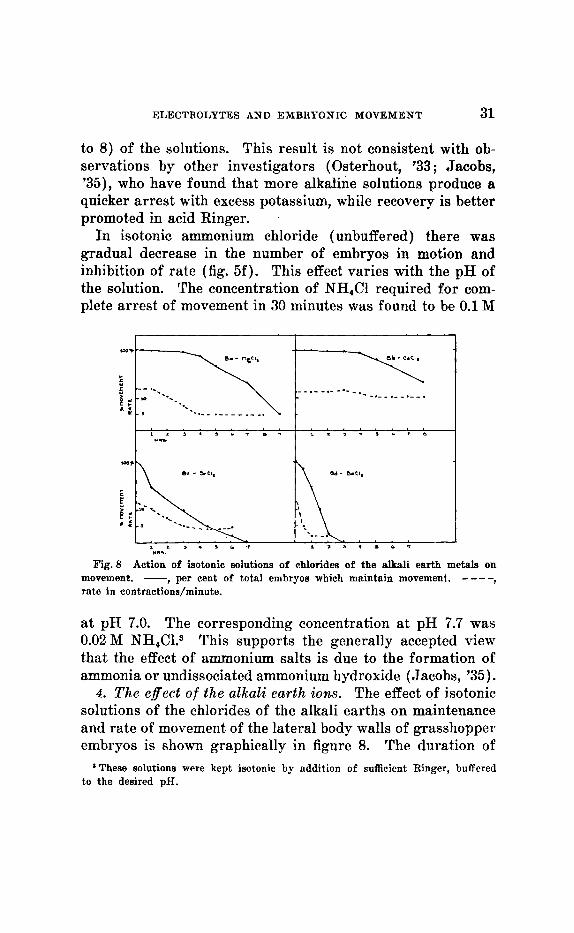
ELECTROLYTES AND EMBRYONIC MOVEMENT 31
Lm*.
5 e g : ,
to 8) of the solutions. This result is not consistent with ob- servations by other investigators (Osterhout, '33 ; Jacobs, '35), who have found that more alkaline solutions produce a quicker arrest with excess potassium, while recovery is better promoted in acid Ringer.
In isotonic ammonium chloride (unbuffered) there was gradual decrease in the number of embryos in motion and inhibition of rate (fig. 5f). This effect varies with the pH of the solution. The concentration of NH,C1 required for com- plete arrest of movement in 30 minutes was found to be 0.1 M
- - .-_.__.--. ,-- ..* I _ - ._ -.- - .- -.
lo -*..
L z J . 3 * 7 L . % = , . , - ? * "CI
a t pH 7.0. The corresponding concentration at pH 7.7 was 0.02 M NH,C1.3 This supports the generally accepted view that the effect of ammonium salts is due to the formation of ammonia or undissociated ammonium hydroxide (Jacobs, '35).
4 . The effect of the alkali earth ions. The effect of isotonic solutions of the chlorides of the alkali earths on maintenance and rate of movement of the lateral body walls of grasshopper embryos is shown graphically in figure 8. The duration of
'These solutions were kept isotonic by addition of sufficient Ringer, buffered to the desired pH.
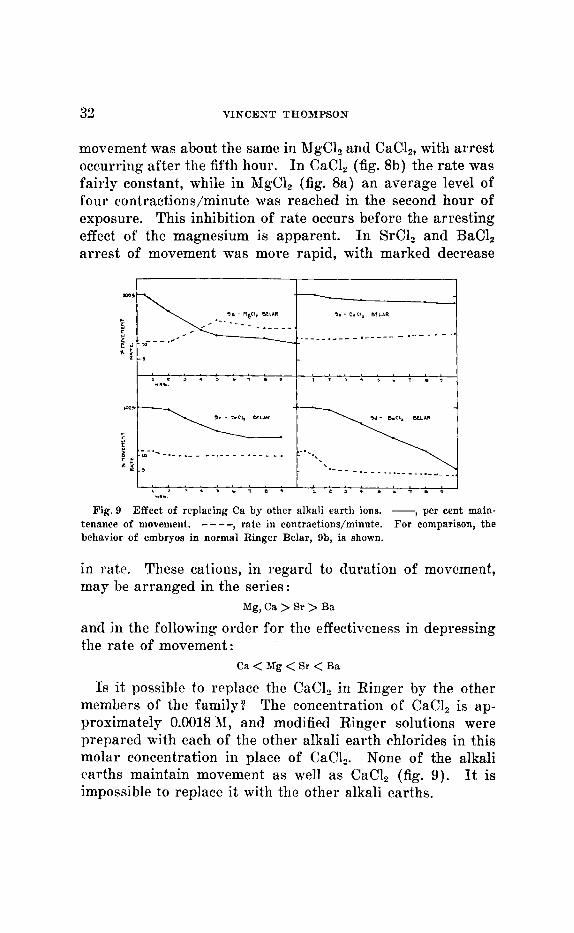
33 VINCENT THOMPSON
movement was about the same in RZgC1, and CaCI,, with arrest occurring after the fifth hour. In CaC1, (fig. 8b) the rate was fairly constant, while in RIgC1, (fig. 8a) an average level of four contractions/minute was reached in the second hour of exposure. This inhibition of rate occurs before the arresting effect of the magnesium is apparent. I n SrCl, and BaCl, arrest of movement was more rapid, with marked decrease
Fig. 9 Effect of replacing Ca by other alkali earth ions. - , per cent main- tenance of movement. ---- , rate in contractions/minute. For comparison, the behavior of embryos in nornial Ringer Belar, 9b, is shown.
in rate. These cations, in regard to duration of movement, may be arranged in the series:
and in the following order for the effectiveness in depressing the rate of movement:
Mg, Ca > Sr > Ba
Ca < Mg < Sr < Ba
Is it possible to replace the CaCl, in Ringer by the other members of the family? The conceiitratioii of CaCI, is ap- proximately 0.0018 11, and modified Ringer solutions were prepared with each of the other alkali earth chlorides in this molar concentration in place of CaC1,. None of the alkali earths maintain movement as well as CaCI, (fig. 9) . It is impossible to replace it with the other alkali earths.

ELECTROLYTES AND EMBRYONIC MOVEMENT 33
In general these ions depress the rate of rhythmic move- ments, although an initial stimulation with barium is some- times observed (Chao, '33). Lindeman ('28), with the crayfish heart, observed a decrease in rate with excess magnesium or calcium, and Chao ( '33), using the Limulus heart-nerve prepa- ration, reported that calcium, magnesium and strontium were effective in depressing the rate of movement. Magnesium has the same effect on embryonic amphibian hearts (Koller, '32). Calcium, strontium and barium show a gradation in effect on physiological activity of skeletal and cardiac muscles of the frog (Mines, 'll), with magnesium behaving as calcium in respect to the former and differently in action on cardiac muscle. This difference, between response to excess calcium and magnesium, has also been observed for amoeboid (Pantin, '26) and ciliary (Gray, '28) movements.
5. Ion antagonism. This section is concerned with antago- nism of the action of potassium on lateral body wall contrac- tions by the alkali earth ions. Various mixtures of isotonic solutions of potassium and calcium chlorides were used. No antagonism was apparent if more than 5 parts KC1 in 10 of solution were tested. The series with 50% (0.08 M) KC1 is shown (fig. 10). The KCl was kept constant ( 5 parts) and the CaC1, varied, with sufficient isotonic dextrose solution present to make a total of 10 parts of solution. The following points should be noted. 1) In excess potassium, two results are seen : movement was quickly arrested and the rate acceler- ated (fig. 10a). 2) The effect of potassium on rate of move- ment (acceleration) was antagonized first by the calcium. No stimulation was evident with 20% (0.024M) or more CaCl,. 3) The stoppage action of potassium was imperfectly antago- nized by calcium. In the series with 50% KCl (fig. 10, a to f ) , the first part of the maintenance (per cent movement) curves does not vary greatly. In all cases about 80% of the embryos had ceased movement at the end of 2 hours, regardless of the amount of recovery later. Even with 4 parts KCl plus 6 parts CaC1, (fig. log) movement was arrested in 50% of the cases. The only possible method found to prevent this initial arrest

34 VINCENT THOMPSON
of movement in excess potassium (i.e., to completely antago- nize the potassium effects) was to give the embryos a bath in CaCl, solution before exposing them to the potassium. The time required for this treatment varied with the amount of calcium in the ‘conditioning’ solution, and with the potassium
. ?D ,.-*-. 10 L
l l , . H”3
“C, - 3 c 1 c q .* DLLIIOSL - ,
l i 3 . 5 . 7 l l , . , * L 1 3 . S “ w 3
Fig. 10 Antagonism of action of K by Ca. - , pet cent maintenance of activity. The proportions by volume of isotonic solutions of KCl, CaCl, and dextrose are shown on each graph.
- - - - , rate of movement in contractions/minute.
% L 1 4 5 L YRL.
Fig.11 Complete antagonism of the initial stoppage effect of K. - 7 per cent of total embryos in motion. Embryos treated 3 hours in mixture consisting of equal parts isotonic CaCI, plus NaCI, and then transferred to a solution with equal parts CaCI, plus KCI. This figure should be compared with figure 10f.
---- , rate i n contractions/minute.

ELECTROLYTES AND EMBRYONIC MOVEMENT 35
concentration in the final medium. The behavior of embryos treated in this way is illustrated graphically in figure 11.
It has already been shown that magnesium and calcium do not influence lateral body wall movement in the same manner. Further, magnesium does not act the same in antagonism (fig.. 12). In all KCI-hlgC1, mixtures the accelerating effect of the potassium ion was seen, although some antagonism of the stoppage action was observed (fig. 12, c to e). These re- sults may be contrasted with those obtained with embryos in
I ,.- -.- _. I ._ - -. , -.--._
1c1 -. n,ct, - 5 DI.1RU011 - ,
L . . . = C T b 7 “.b,
Fig. 12 KCI-MgCI, antagonism. As figure 10, except isotonic MgCI, in place of CaCl,.
employed
KCI-CaCl, mixtures, where the acceleration effect disappeared with relatively low calcium concentrations.
The effects of rubidium and caesium on lateral body wall contractions may be antagonized by calcium (fig. 13). Another similarity, in addition to those mentioned above, between potassium and rubidium is evident from figures 13a and 10e.
Anions 1. Common aizions. Figure 14 represents graphically the
results of treating the grasshopper embryos with isotonic

36 VINCENT THOMPSON
L L J t S b T ~ nu,.
Fig. 13 Antagonism of action of Rb (13s) and Cs (13b) by Ca. -, per cent maintenance of movement. - - - - , rate of movement in contractions/minute. The proportions of isotonic solutions of the salts are shown on the graphs.
' I , . 3 ' 7 1 1 3 . 1 . L I J I S *"5
l i d - nar
I L 3 t = * . r * 9 L L 3 . O C . I D 3 ""3.
Fig. 14 Effects of isotonic solutions of sodium salts of anions. -, per cent maintenance of movement. - - - - , rate of movement in eontraetions/minute.

ELECTROLYTES AND EMBRYONIC MOVEMENT 37
solutions of the sodium salts of certain anions. Calcium chlo- ride was lacking in all cases, so the curves may be compared with those for pure NaCl (fig. 3a). Nitrate, bromide and iodide maintain movement fairly well, with a decided accelera- tion of rate in the bromide. In sulphate there is an initial decrease in rate, followed by a gradual recovery. The main- tenance is not so satisfactory, although about 50% of the embryos continued movement during the entire experimental period. Thiocyanate is the least suitable of any of these anions (fig. 14e).
In general the more common anions may replace the chlo- rides in saline without a great degree of harmful effects. Sakai (quoted by Gray, '22) found that the beat of the frog's heart was well maintained in iodides, bromides or nitrates. Sulphate was also satisfactory, but an initial slowing was observed. Gray ('22) found that cilia of Mytilus continued movement in chlorides, nitrates, bromides, iodides and sul- phates. Scheuring ( '2T) reported that spermatozoa of Ar- bacia retained their motility better in nitrates than in sea water.
Of all the anions studied, fluoride was the most toxic. A Ringer solution, 0.00075 in NaF, caused complete arrest of contractions in about 2 hours with a marked depression in rate.
2. Action. of certain acid-forming anions. It was found that certain concentrations of sodium acetate (buffered to pH 6.6) stopped all movement in a very few minutes after the embryos were immersed in the solutions (fig. 15). In 0.024 M acetate very little recovery occurred. Embryos in 0.018 M solution (fig. 15b) were initially arrested, but recovery of movement was quite rapid. About the same results were obtained with 0.015M, but in 0.012M acetate not all the embryos ceased movement and recovery was 100% (fig. 15c). In 0.006 M the initial inhibition affected only about 50% of the individuals, and in 0.002M acetate this phenomenon has practically dis- appeared. This sudden initial arrest with subsequent recov-
'All solutions were made isotonic by addition of sufficient Ringer. The pH was checked for each solution.

38 VINCENT THOMPSON
L0a.h
2 a $ K
c
ery of movement also occurs in solutions of sodium formate, butyrate and bicarbonate.
It has been suggested (Irwin, '27; Stewart, '31; Becker, '36) that the effectiveness of the fatty acid and bicarbonate salts is the result of the formation of undissociated acid by hydrolysis. If the undissociated acid is the toxic factor in such a solution, and if the resultant toxic effect is merely a question of the Concentration of undissociated acid, then varia- tion in hydrogen ion concentration should produce a corre- sponding variation in the degree of toxicity of the solution.
t,.. oat." C.AC > S b - O O l h n hk
-Y)
- 0
It is possible to calculate the amount of undissociated acid, and this concentration in a salt solution which just produces the complete initial arrest of movement (as does a 0.015 11 acetate solution at pH 6.6) should be constant over a suitable hydrogen ion concentration range.
Accordingly, the concentration of anions, which would just give. the effect observed above, was determined over a wide pH range. Acetate, formate, butyrate and bicarbonate were used. Solutions were prepared by mixing the fatty acid (or HCl for the bicarbonate) and corresponding sodium salt

ELECTROLYTES AND EMBRYONIC MOVEMENT 39
solutions (of equal concentrations) to give the desired pH. Where extra buffers were needed, glycine-HC1 or glycine- NaOH or phosphate systems were used. The hydrogen ion Concentration of all solutions was checked electrometrically. In figure 16 the logarithm of the concentration of anion which just gave the initial arrest of movement is plotted against the pH at which the solution was buffered. From these curves
1 3 4 3 6 7 8
PH
Fig. 16 Variation of effective anion concentration with pH. Logarithm of total anion concentration (ordinate) plotted against the pH of the solution (abscissa). Curves for formate, acetate, butyrate and bicarbonate solutions. For further explanations see text.
the anion concentration for the indicated pH values (table 2) were taken, and the amount of undissociated acid calculated, employing the following expression :
Ct Y [H'l [HA] = [H'l Y + KO
where [HA] is the concentration of undissociated acid ; C,, the total concentration of anion; [H'], the hydrogen ion concen- tration, and y, the ionic activity coefficient. This was deter- mined from the data of Noyes and hfacInnes ('20) on KCl, after calculation of the ionic activity of Ringer by the square

40 VINCENT THOMPSON
of the valence rule (Lewis and Randall, '23). y was found to be about 0.72. This equation is identical with that of Smith ('25) although not in exactly the same form.
The variation in undissociated acid concentration (table 2) is relatively small. For example, while the toxic concentra- tion of total acetate varies from 0.25 M at pH 8 to 0.005 M at pH 4, the undissociated acetic acid changes from 9.7 x 10-j
TABLE 2
Anion and undisaociafed acid concentration
Bcetate
Butyrate
Anion PH Formate 8
7 6 5 4 3
8 7 6 5 4 3
8 7 6 5 4 3
Bicarbonate 8 7
l og . c, - 0.25 - 0.90 - 1.50 - 2.20 - 2.80 - 3.30
- 0.60 - 1.45 - 2.30 - 2.95 - 3.30 - 3.40
- 1.20 - 2.10 - 3.00 - 3.30 - 3.35 - 3.35
- 1.30 - 3.30
ct 0.56 M 0.126 0.032 0.0062 0.0016 0.0005
0.35 0.035 0.005 0.0012 0.0005 0.0004
0.063 0.0079 0.001 0.0005 0.00045 0.00043
0.050 0.005
[ H A ' ]
1.88 x lO-'hl 4.24
10.7 20.5 40.3 38.6
9.7 13.5 16.6 33.5 40.0 39.0
3.07 3.82 4.65
16.5 37.2 43.1
11.7 X 10-'M 9.7
Log. C,, obtained from figure 16. C,, total anion Concentration. lated undissociated acid concentration.
[HA'], ealcu-
to 40 X lop5 M ; a change of 500 times for the anion compared with only four times in undissociated acid concentration. How- ever, it should be noted that there is uniformity in increase in [HA] with a diminishing anion concentration or with de- creasing pH. This reached a limiting value, about 40 x M for all three fatty acids. While the initial stoppage action of these substances may be attributed to the undissociated

ELECTROLYTES AND EMBRYONIC MOVEMENT 41
acid, the buffer r’elationships (hydrogen ion and anion con- centrations) evidently play some role in determining the con- centration of undissociated acid necessary for the initial arrest of movement.
There was some evidence of a different rate of penetration of these acids into the grasshopper embryo. In the formate solutions about 30 minutes were necessary for the maximal stoppage action, while in acetate and butyrate 15 minutes or less were required. Stewart (’31), who studied the penetra- tion of fatty acids into Arbacia eggs, concluded that the rela- tive order of rate of entrance was : formate < acetate < buty- rate, which she related to the order of lipoid solubility.
The poisonous effect of a cyanide solution is also due to the formation of undissociated acid. At pH 10 the toxic amount (to completely arrest movement in 15 minutes) is 0.15 RI NaCN, while a t pH 9, 8 or 7 the corresponding concentration is 0.02 11.
DISCUSSION
The only theory, by which the action of cations on different rhythmic systems has been interpreted, assumes that there are two processes concerned with rhythmic movement which are influenced in various ways by different ions. One of these is an excitation process, which is “concerned with events prior to contraction” (Hogben, ’25)’ and the other is the actual contraction itself, i.e., the mechanical movement. Hogben (’25) suggested this hypothesis in his work on the compara- tive physiology of contractile systems. Pantin ( ’26) applied the idea to amoeboid movement, Clark (’27) to heart con- traction, Gray (’28) to ciliary movement, and recently, Walzl (’37) has shown that the behavior of the oyster heart is con- sistent with the hypothesis.
Calcium supposedly depresses that mechanism (a) con- cerned with excitation and augments the contraction mech- anism (b). Potassium and sodium stimulate (a) and inhibit (b). Now, there may be a great variation in effect of a par- ticular ion on different types of movement. For example, with excess potassium in the perfusion medium, hearts are

42 VINCENT THOMPSON
arrested in systole (invertebrates) or diastole (vertebrates) ; rate of movement is accelerated until stoppage occurs (lateral walls of grasshopper embryos, Oryzias embryonic movements, Yamamoto, ’34, and others) ; or, if the concentration of potas- sium be sub-toxic, merely an increase in rate will be evident. Such variations are interpreted on the assumption that, with a particular ion, mechanisms (a) and (b) in different rhythmic systems are influenced in different proportions.
Excess potassium accelerates the rate of lateral body wall movement of the grasshopper embryo, and also arrests move- ment. In terms of the theory outlines above, potassium ex- cites mechanism (a) and depresses (b). This assumes that a change in rate of lateral wall contractions indicates a dis- turbance in (a), and that it is possible to differentiate between the two mechanisms. The results with calcium are not so simple to consider in terms of the theory, since the effects on lateral wall movement are not very striking. However, from antagonism of the potassium effects it might be surmised that calcium depresses the excitatory mechanism (a), and stimu- lates the contraction mechanism (a).
The value of this theory lies in that it gives a more or less common ground for comparison of effects of cations on dif- ferent rhythmic systems. The assumption of two separable mechanisms involved in contraction, with a possibility of each being influenced in different proportions by various cations, gives a theory with which it would be difficult to imagine any influence of any substance on any type of movement which could not be ‘interpreted.’ The separation of processes in- volved in contraction has been questioned by Fulton (’26, p. 228), who concludes his discussion, “. . . . evidence, there- fore, . . . . appears more and more to point toward the con- clusion that mechanical and electrical responses are in fact inseparable.” Further, it is probable that the cations do not actually come in contact with the contractile elements of the cells. Most animal cells are impermeable to cations, or, at best, penetration occurs very slowly (Jacobs, ’35, or other reviews on permeability). The action of these ions is on the exterior of the cells, and the result (arrest of movement in

ELECTROLYTES AWD EMBRYONIC MOVEMENT 43
diastole or systole, or acceleration or depression of rate of contraction) is the visible end-point of a long series of ‘effects’ of the cation. In other words, there is no evidence that these cations directly affect the contractile process, to say nothing of separately influencing two contractile mechanisms.
Gray (’28) states that the anions which affect (ciliary) movement are those which interfere in a chemical way with the calcium ratio (as precipitation of calcium by sulphate, fluoride, oxalate, etc.). A change in the calcium content in the surrounding medium does not greatly alter the behavior of the lateral body wall movements of the grasshopper embryo. The extreme effect of such an ion as the fluoride is probably due to some specific toxic action. Another class of anions, illustrated by the bicarbonates, fatty acid anions and cyanides, form by hydrolysis penetrating acids which are toxic to the lateral wall movements, and are, themselves, not injurious.
SUMMARY
1. The action of various electrolytes on the lateral body wall movements of grasshopper embryos has been studied. These movements are vigorous, rhythmic contractions which are evi- dent during yolk engulfment, and are maintained satisfac- torily in a modified Ringer solution.
2. It is possible to replace 80% of the salts in Ringer by dextrose. Urea and glycerol cannot be employed for this pur- pose, as they penetrate and are therefore osmotically inactive for the embryos. Urea, itself, is otherwise not harmful.
3. KC1 may be omitted from the Ringer solution without deleterious effects. NaCl and CaCl, are essential for proper functioning of the lateral body wall contractions.
4. Ca cannot be replaced by any of the other alkali earth ions.
5. Movement is maintained in Ringer solutions from pH 3.7 to 10.6, using HCl or NaOH to give the desired hydrogen ion concentrations.
6. Duration of movement in isotonic solutions of the alka- lies is in the series: Na > Li > Cs > Rb > K ; in the alkali earths: Mg, Ca > Sr > Ba.

44 VINCENT THOMPSON
7. Rate of movement (contractions/minute) is accelerated in the alkalies: Li < Na < Cs < I< < Rb. Rate is depressed in the alkali earths: Ca < Mg < Sr < Ba.
8. The effects of potassium may be completely antagonized by calcium. The rate acceleration effect is antagonized by low Ca concentrations, while the stoppage action of K requires special treatment in excess Ca.
9. The sodium salts of common anions, as iodide, bromide and nitrate may replace the chloride in Ringer. Sulpliate is not as satisfactory, and thiocyanate is definitely harmful.
10. Acetate, formate, butyrate ancl bicarbonate ions them- selves do not interfere with maintenance of movement. Their effect is clue to the formation, by hydrolysis, of undissociated acids. These acids produce immediate arrest of movement, from which the embryos are able to recover over a considera- ble range of concentration. Cyanides, except for a more limited recovery, also belong to this group.
LITERATURE CITED
BAETJER, A. A., AND C. H. MCDONAILI 1931 The relation of sodium, potassium Am. J. Physiol., vol. 99,
BAYLISS, W. M. 1924 Principles of General Physiology. 4th ed., pp. 207-235. London.
BECKER, Z. E. A comparison between the action of carbonic acid and other acids upon the living cell. Protoplasma, vol. 25, pp. 161-175.
BELAR, K. 1999 Beitrage zur Kausalanalyse der Mitose. 11. Untersuchungen a n den Spermatocyten von Chorthippus (Stenobothrus) lineatus Panz. Arch. f. Entwick. d. Org., Bd. 118, S. 359486.
1. Effect of temperature on the rate of development of certain Orthoptera. J. Exp. Zool., vol. 42, pp. 91-109.
1906 On the chemical conditions for the heart activity, with special reference to the heart of Liniulus. Am. J. Physiol., vol. 16, pp. 278108.
Notes on taxonomy, development, and l ife history of certain Acrididae. - 1937 Culture methods for grasshoppers. Culture Methods for In- vertebrate Animals, pp. 287-291. Ithaca.
Action of electrolytes on the dorsal median nerve cord of the Limulus heart. Biol. Bull., vol. 61, pp. 358-382.
Hydrogen-ion concentration and the rhythmic activity of the
and calcium ions to the heart rhythmicity. pp. 666-680.
1936
BODINE, J. H.
CARLSON, A. J.
1925
CAROTHERS, E. E. 1923 Trans. Am. Ent. Soe., vol. 49, pp. 7-24.
CHAO. IPING 1933
1935 nerve cells in the ganglion of the Liinulus heart. Ibid., vol. 68, pp. 69-73.

ELECTROLYTES AND EMBRYONIC MOVEMENT 45
CLARK, A. J. 1927 Comparative Physiology of the Heart. Cambridge Univer-
FALK, I. S. 1923 The role of certain ions in bacterial physiology. A review. Abs. Bact., vol. 7, pp. 33-50, 87-105,
A comparison of the effects on the isolated beating in- Am. J. Physiol., vol. 72, pp.
sity Press.
(Studies on salt action. \TI.) 133-147.
FRASER, LOIS M. 1925 testine of CO, and of ‘a mineral acid. 119-124.
FULTON, J. F. 1926 Muscular Contraction. Baltimore. GELLHORN, E R N s r 1927 Ionwirkung und Zelldurchlassigkeit. Protoplasma, Bd.
GRAY, J. 1922 The mechanism of ciliary movement. Proc. Roy. SOC. London,
1928- Ciliary Movement. Cambridge University Press.
1, S. 589-609.
ser. B, vol. 93, pp. 104-131.
GREEND, C. W. 1899 On the relation of the inorganic salts of blood to the auto- Am. J. Physiol., vol.
The effects of ammonia, of the f a t ty acids and of their salts, on the luminescence of Bacillus fischeri. J. Cell. and a m p . Physiol., vol. 1, pp. 145-159.
HOBER, RUDOLF 1926 Physikalische Chemie der Zelle und der Gewebe. S. 615- 705. Leipzig.
HOGBEN, L. T. 1925 Studies on the comparative pliysiology of contractile tissues. 1. The action of electrolytes on invertebrate muscle. Quart. J. Exp. Physiol., vol. 15, pp. 263-312.
The effect of acetate buffer mixtures, acetic acid, and sodium acetate on the protoplasm, as influencing the rate of penetration of cresyl blue into the vacuole of Nitella. J. Gen. Physiol., vol. 11, pp. 111-131.
JACOBS, M. H. 1931 The permeability of the erythrocyte. Erg. d. Biol., vol. 7,
matic activity of a strip of ventricular muscle. 2, pp. 82-126.
HILL, SAMUEL E. 1932
IRWIN, MARIAN 1927
pp. 1-55. 1935 Permeability. Ann. Rev. Biochem., vol. 4, pp. 1-16.
EOLLER, G. 1932 Versuche an nervenfreien embryonnlen Amphienherzen. Zeit.
LEWIS, G. N., AND M. RANDALL 1922 Thermodynamics. New York. LEWIS, W. H.
Wise. Biol. Abt. Vergleich. Physiol., Bd. 18, S. 186-203.
1929 The effect of various solutions and salts on the pulsation rate of isolated hearts from young chick embryos. Carnegie Inst. Wash., vol. 394, pp. 173-192.
LINDENAN, V. F. 1928 The physiology of the crustacean heart. 1. The effect of various ions upon the heart rhythm of the crayfish, Cambarns clarkii. Physiol. Zoo]., vol. 1, pp. 576-592. 1900 On ion-proteid cornpounds and their role in the mechanics of life phenomena. 1. The poisonous character of a pure NaCl solution. Am. J. Physiol., vol. 3, pp. 327336.
LOEB, J.
1912 The Mechanistic Conception of Life. Chicago. LUSSANA, F. 1911 Action des sels inorganiques sur I’irritabilit6 de coeur de
greuouille isol6. Arch. Intern. de Physiol., vol. 11, pp. 1-23.
THE .IOURXAL OF EKPBRIMENTAL z o o m y , VOL. 78, NO. 1

46 VINCENT THOMPSON
MARTIN, E. J. 1904 The inhibitory influence of KCI on the heart, and the effect of variations of temperature upon this inhibition and upon vagus in- hibition.
Influence of temperature upon the action of potassium chloride on the heart beats of Oryzias. J. Fae. Sci. Tokyo, Sect. 4,
MINES, G. R. On the replacement of calcium in certain neuromuscular mechanisms by allied substanres.
NELSEN, 0. E. Life cycle, sex differentiation and testis development in Melanoplus differentialis.
NOYES, A. A., AND D. A. MACINNIB The ionization and activity of largely ionized substances. J. Am. Chem. Soe., vol. 42, pp. 239-245.
OSTERHOUT, W. J. V. 1933 Permeability in large plant cells and in models. Erg. d. Physiol., vol. 35, pp. 967-1021.
PANTIN, C. F. A. 1926 On the phpsiology of amoeboid movement. 111. The action of calcium. J. Exp. Biol., vol. 3, pp. 275-296.
1931 On the physiology of amoeboid movement. VII I . A. The action of certain non-electrolytes. B. A note on the isoelectric point of the proteins of a marine amoeba.
Concerning the influence exerted by each of the constitu- ents of the blood on the contraction of the ventricle. J. Physiol.,
SLIFER, E. H. 1932 a Insect development. 111. Blastokinesis in the living grass- hopper egg.
1932 b Insect development. IV. External morphology of grass- hopper embryos of known age and with a known temperature history. J. Morph., vol. 53, pp. 1-21.
-~ 1934 Insect development. VI. The behavior of grasshopper em- bryos in anisotonic, balanced salt solutions. J. Exp. Zool., vol. 67,
SMITH, HOMER W. 1925 The action of acids on cell division with reference to permeability to anions.
STEWART, DORQTHY R. 1931 The permeability of the Arbacia egg to ammonium salts.
THOMPSON, VINCENT 1937 Effects of temperature on movements of embryos (Acrididae, Orthoptera).
THORNER, M. W. Recovery of the heart beat of Fundulus embryos a f te r stoppage by potassium chloride.
WALKER, J. E. Effect of carbon dioxide tension on beating of lateral body walls of grasshopper embryos. J. Cell. and Comp. Physiol., vol. 6,
WAUL, E. M. Action of ions on the heart of the oyster (Ostrea virginica).
WITSOHI, EMIL 1936 Aufzucht und Haltung der gebraiichlichen Laboratorium- Handbuch d. Biol. Arbeitsmethoden,
YAMAMOTO, TOKI-o 1934 Studies on the rhythmical movements in the early V. The action of electrolytes and osmotic
Am. J. Physiol., vol. 11, pp. 37-93. MATSURA, YOSHIO 1934
V O ~ . 3, pp. 509-516. 1911
J. Physiol., vol. 42, pp. 251-266. 1931
J. Morph., vol. 51, pp. 467-525. 1920
Ibid., vol. 8, pp. 365-378. RINQEIC, SIDNEY 1882
V O ~ . 3, pp. 380-393.
Biol. Zentralb., Bd. 67, S. 824-229.
pp. 137-157.
Am. J. Physiol., vol. 72, pp. 347-371.
Biol. Bull., vol. 60, pp. 171-178.
Physiol. Zool., vol. 10, pp. 21-30. 1929
Biol. Bull., vo1. 56, pp. 157-163. 1935
pp. 317-334. 1937
Physiol. Zool., vol. 10, pp. 125-140.
tiere Amphibian und Reptilien. Bd. 9, S. 611-651.
embryo of Oryzias latipes. pressure. J. Fac. Sci. Tokyo, Seet. 4, vol. 3, pp. 887-300,


















