
Natural Sciences 360 Legacy of Life Lecture 08 Dr. Stuart S. Sumida Finishing Protostomes.
Upload
trinhxuyenCategory
view
212download
0
4387DEVELOPMENT AND STEM CELLS RESEARCH ARTICLE
INTRODUCTIONThe planarian Schmidtea mediterranea can regenerate from smallbody fragments (Reddien and Sánchez Alvarado, 2004). This robustregenerative capacity, together with emerging molecular tools forplanarian gene function studies, make these animals a powerfulsystem for studying the regeneration of cell types, tissues and evenentire organ systems. In addition to uncovering regenerativemechanisms, studying the regeneration of planarian cell types couldalso identify broadly used developmental mechanisms.
Here, we investigate the genetic mechanisms underlyingregeneration of the planarian protonephridial system, which isinvolved in waste excretion and osmoregulation. Excretorysystems consist of a filtration surface and a tubule, whichmodifies the ultrafiltrate by reabsorption and secretion andconnects to the animal exterior (Ruppert and Smith, 1988;Bartolomaeus and Ax, 1992; Ruppert, 1994). Protonephridiaconsist of blind tubules ending in a terminal cell (Ruppert andSmith, 1988; Bartolomaeus and Ax, 1992), in which beating ciliagenerate negative pressure allowing filtration from theextracellular space into the tubule lumen through membranefenestrations (Ruppert and Smith, 1988; Bartolomaeus and Ax,1992). Metanephridia involve specialized epithelial cells calledpodocytes, which filter pressurized fluid from circulatory systemsinto a tubule (Quaggin and Kreidberg, 2008). Within the Bilateria,most deuterostomes have metanephridia, whereas bothprotonephridia and metanephridia are found within protostomes(Bartolomaeus and Ax, 1992) (see Fig. S1 in the supplementarymaterial). In addition to morphological differences at the filtrationsite, the two nephridial systems can also have different germ layerorigins; most invertebrate nephridia (protonephridia and
metanephridia) have an ectoderm origin, whereas metanephridiain vertebrates have a mesoderm origin. These characteristics,together with the presence of both systems in differentdevelopmental stages of some invertebrates, have generateddisparate conclusions regarding the homology of nephridia(Wilson and Webster, 1974; Bartolomaeus and Ax, 1992;Ruppert, 1994). Therefore, despite the importance and ubiquityof excretory systems, their evolutionary origin remains unclear.
The vertebrate kidney is the most extensively studiedmetanephridial system (Dressler, 2006; Dressler, 2009; Costantiniand Kopan, 2010). In the mouse, nephrons are the basic functionalunits of kidneys and develop from the intermediate mesoderm, andnumerous genes involved in their development have beencharacterized (Dressler, 2009). Protonephridia are mostly found insmall adult animals or larvae of several phyla, includingNermertea, Gastrotricha, Platyhelminthes, Mollusca and Annelida(Ruppert and Smith, 1988; Bartolomaeus and Ax, 1992). The twoclassical protostome model organisms, Caenorhabditis elegans andDrosophila melanogaster, have specialized excretory systems withonly one excretory cell in the former (Nelson et al., 1983) and aseparation of ultrafiltration from excretion in the latter (Denholmand Skaer, 2009). Therefore, despite morphological andultrastructural studies of protonephridial systems in severalorganisms, our understanding of the regulatory genes required forprotonephridia formation is limited.
Most animals share a discrete and relatively small number oforgan systems, such as eyes, muscle, gut, heart and an excretorysystem. Characterization of regulatory programs required for theformation of these major tissues/organs is a central challenge indevelopmental biology. The identity or existence of a commongenetic regulatory program for excretory systems has not beenestablished. Here, we identify a regeneration program for theplanarian tubule-based filtration excretory system. Our resultssuggest that there existed a genetic program for nephridiaformation in the common ancestor of the Bilateria withcomponents still responsible for the formation and function ofvaried excretory systems in extant bilaterians.
Development 138, 4387-4398 (2011) doi:10.1242/dev.068098© 2011. Published by The Company of Biologists Ltd
Howard Hughes Medical Institute, Whitehead Institute, and Department of Biology,Massachusetts Institute of Technology, 9 Cambridge Center, Cambridge, MA 02142,USA.
*Author for correspondence ([email protected])
Accepted 15 August 2011
SUMMARYPlanarians can regenerate any missing body part, requiring mechanisms for the production of organ systems in the adult,including their prominent tubule-based filtration excretory system called protonephridia. Here, we identify a set of genes, Six1/2-2, POU2/3, hunchback, Eya and Sall, that encode transcription regulatory proteins that are required for planarian protonephridiaregeneration. During regeneration, planarian stem cells are induced to form a cell population in regeneration blastemasexpressing Six1/2-2, POU2/3, Eya, Sall and Osr that is required for excretory system formation. POU2/3 and Six1/2-2 are essentialfor these precursor cells to form. Eya, Six1/2-2, Sall, Osr and POU2/3-related genes are required for vertebrate kidneydevelopment. We determined that planarian and vertebrate excretory cells express homologous proteins involved in reabsorptionand waste modification. Furthermore, we identified novel nephridia genes. Our results identify a transcriptional program andcellular mechanisms for the regeneration of an excretory organ and suggest that metazoan excretory systems are regulated bygenetic programs that share a common evolutionary origin.
KEY WORDS: Kidney, Neoblast, Planarian, Protonephridia, Regeneration, Stem cell
A regulatory program for excretory system regeneration inplanariansM. Lucila Scimone, Mansi Srivastava, George W. Bell and Peter W. Reddien*
DEVELO
PMENT

4388
MATERIALS AND METHODSRNAi experimentsRT-PCR and RACE were used to amplify and determine genesequences. Genes were cloned into pPR244 for RNA interference(RNAi) as described (Reddien et al., 2005a). cDNAs used to generateRNA probes were: SAAH-aaa15g04 (carbonic anhydrase VII, CA),H.85.1h (alkaline phosphatase), H.2.8b (rootletin), H14.9d (EF-handdomain) and SAAH-aaa96a06 (ring finger). Six1/2-2, POU2/3, Sall andhunchback RNAi was performed by feeding. Bacterial culture (10 ml)was pelleted and resuspended in 30 l of liver. Animals were fed ondays 0, 4 and 7. For regeneration, animals were amputated at day 8 andscored at day 15 (Six1/2-2 RNAi), or fed and cut again at day 16 andscored at day 23 (POU2/3 and hunchback RNAi). For intact animalsundergoing tissue turnover, feedings were on days 0, 4 and 7 for Six1/2-2 with an additional feeding on day 14 for POU2/3 and hunchback,seven additional feedings for Eya, and four extra feedings for Sall. ForRNAi by injection, dsRNA from in vitro transcription reactions(Promega) was injected on days 0, 1 and 2, with animals amputated onday 2. Six days later, animals were injected and amputated again, withfixation six days thereafter.
In situ hybridization and immunolabelingIn situ hybridization and immunolabeling were performed as described(Pearson et al., 2009; Glazer et al., 2010). Numbers of carbonic anhydrase(CA)-expressing cells and tubule branches in clusters were counted in tailsof intact animals and in the anterior and posterior blastemas of regeneratingtrunk pieces. X1 cells were isolated as described (Scimone et al., 2010),adhered to poly-L-lysine-coated coverslips and fixed in 4%paraformaldehyde for 20 minutes. Images were obtained using a ZeissAxioImager with Apotome or a Zeiss LSM710 confocal microscope.
Transmission electron microscopyAnimals were fixed in 2.5% glutaraldehyde, 3% paraformaldehyde with5% sucrose in 0.1 M sodium cacodylate buffer (pH 7.4) for one hour, thenpost-fixed in 1% OsO4 in veronal-acetate buffer. Animals were stainedovernight with 0.5% uranyl acetate in veronal-acetate buffer (pH 6.0),dehydrated, and embedded in Spurr’s resin. Sections were cut on a ReichertUltracut E microtome with a Diatome diamond knife at a thickness settingof 50 nm then stained with 2% uranyl acetate and lead citrate. The sectionswere examined using a FEI Tecnai spirit at 80 KV and photographed withan AMT CCD camera.
Microarray experimentsRNA from Six1/2-2(RNAi) and POU2/3(RNAi) animals was harvestedwith Trizol at days 8 and 15 after RNAi initiation for the former anddays 15 and 21 for the latter. Three biological replicates were used. Cy3-and Cy5-labeled cRNA was prepared using a QuickAmp Labeling Kit(Agilent) starting with 1 g total RNA. Agilent custom planarian4�44,000 expression arrays were hybridized according themanufacturer’s instructions and scanned using an Agilent DNAmicroarray scanner. Array images were quantified and statisticalsignificance of differential expression was calculated using Agilent’sFeature Extraction Image Analysis software with the default two-colorgene expression protocol. Agilent two-color arrays were within-arraynormalized by loess, followed by between-array quantile normalizationof average intensities across channels (Aquantile). Differentialexpression analysis was performed using a moderated t-test, asimplemented in the limma package of Bioconductor, with P-valuecorrection by false discovery rate. Microarray data are available athttp://www.ncbi.nlm.nih.gov with accession GSE31618.
RESULTSSix1/2-2, POU2/3 and hunchback are required forexcretory system functionTo study organ system regeneration we sought genes in theplanarian S. mediterranea that are required for organ functionsusing RNAi screening. Inhibition of three genes resulted in asimilar phenotype that suggested a defect in excretion/
osmoregulation [this work and the work of Reddien et al. (Reddienet al., 2005a)]. Smed-Six1/2-2 (abbreviated to Six1/2-2) encodes ahomeodomain-containing protein homologous to the Drosophilasine oculis gene (see Fig. S2A in the supplementary material),Smed-POU2/3 (abbreviated to POU2/3) encodes a representativeof the POU family of homeobox genes (see Fig. S2B in thesupplementary material) and Smed-hunchback (abbreviated tohunchback) encodes a zinc finger protein homologous to theDrosophila hunchback gene (see Fig. S2C in the supplementarymaterial). Following RNAi of Six1/2-2, POU2/3 or hunchback,animals regenerated but displayed blisters, were bloated with fluid,and lysed (Fig. 1A, upper row) (Reddien et al., 2005a). Planariansmaintain their adult tissues through the action of neoblasts,proliferative cells that replace aged adult cells (Reddien andSánchez Alvarado, 2004). Inhibition of Six1/2-2, POU2/3 orhunchback during normal tissue turnover in intact animals alsocaused bloating, blistering and lysis (Fig. 1A, lower row).Protonephridia contain cilia (Ruppert and Smith, 1988; Glazer etal., 2010) and inhibition of genes required for cilia biology cancause similar defects to those observed in Six1/2-2, POU2/3 andhunchback RNAi animals, indicating that protonephridiadysfunction might underlie this phenotype (Reddien et al., 2005a;Glazer et al., 2010).
POU2/3 is expressed in tubule cells and Six1/2-2 isexpressed in tubule-associated cellsFlatworm excretory systems have been described in many speciesusing vital dye labeling, and light and electron microscopy(Reisinger, 1922; McKanna, 1968a; McKanna, 1968b). To imageS. mediterranea protonephridia, we combined fluorescence in situhybridization and immunolabeling using Smed-carbonic anhydraseVII (Smed-CA; abbreviated to CA) and Smed-rootletin (abbreviatedto rootletin) RNA probes, which were previously shown to labelthe excretory system (Sánchez Alvarado et al., 2002; Glazer et al.,2010), and an antibody that recognizes acetylated tubulin and labelsplanarian protonephridia cilia (Glazer et al., 2010). CA encodes anenzyme required for the interconversion of carbon dioxide andbicarbonate to maintain acid-base balance (Purkerson andSchwartz, 2007) and rootletin encodes a ciliary rootlet component(Yang et al., 2002). The S. mediterranea excretory system includesat least three cell types: a ciliated terminal cell (anti-acetylatedtubulin+ and often called a ‘flame cell’ because its beating ciliaresemble a flame), a ciliated tubule cell type (labeled with the anti-acetylated tubulin antibody and the rootletin RNA probe), andclusters of CA-expressing cells located in close proximity to thetubules (Fig. 1B,C; see Fig. S3A in the supplementary material)(see note added in proof). In some flatworms, there exist tubule-associated cells, with a suggested excretory role, calledparanephrocytes (Hertel, 1993). Because of the uncertainrelationship between paranephrocytes and the CA-expressing cells,we refer to the CA-expressing cells as tubule-associated cells.
In transmission electron micrographs, S. mediterranea flamecells were subepidermal (see Fig. S3B in the supplementarymaterial) with numerous cilia, membrane fenestrations andperipheral microvilli (Fig. 1D; see Fig. S3B,C in the supplementarymaterial). Cilia beating generates negative pressure, allowingfiltration from the extracellular space into the flame cell throughthe fenestrations (McKanna, 1968a; Ruppert and Smith, 1988). Thefiltrate travels into the tubule lumen (see Movie 1 in thesupplementary material) to the animal exterior. The tubule lumenwas formed by two interdigitating, ciliated cells that lackfenestrations or microvilli (Fig. 1E).
RESEARCH ARTICLE Development 138 (20)
DEVELO
PMENT
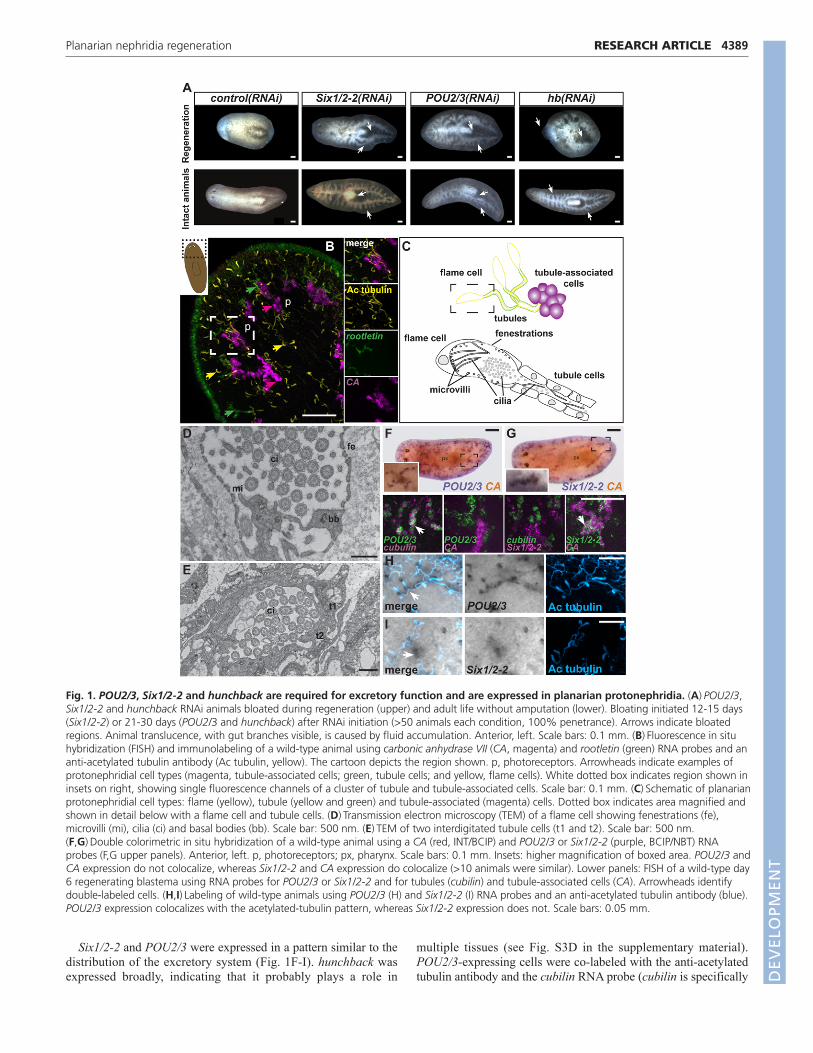
Six1/2-2 and POU2/3 were expressed in a pattern similar to thedistribution of the excretory system (Fig. 1F-I). hunchback wasexpressed broadly, indicating that it probably plays a role in
multiple tissues (see Fig. S3D in the supplementary material).POU2/3-expressing cells were co-labeled with the anti-acetylatedtubulin antibody and the cubilin RNA probe (cubilin is specifically
4389RESEARCH ARTICLEPlanarian nephridia regeneration
Fig. 1. POU2/3, Six1/2-2 and hunchback are required for excretory function and are expressed in planarian protonephridia. (A)POU2/3,Six1/2-2 and hunchback RNAi animals bloated during regeneration (upper) and adult life without amputation (lower). Bloating initiated 12-15 days(Six1/2-2) or 21-30 days (POU2/3 and hunchback) after RNAi initiation (>50 animals each condition, 100% penetrance). Arrows indicate bloatedregions. Animal translucence, with gut branches visible, is caused by fluid accumulation. Anterior, left. Scale bars: 0.1 mm. (B)Fluorescence in situhybridization (FISH) and immunolabeling of a wild-type animal using carbonic anhydrase VII (CA, magenta) and rootletin (green) RNA probes and ananti-acetylated tubulin antibody (Ac tubulin, yellow). The cartoon depicts the region shown. p, photoreceptors. Arrowheads indicate examples ofprotonephridial cell types (magenta, tubule-associated cells; green, tubule cells; and yellow, flame cells). White dotted box indicates region shown ininsets on right, showing single fluorescence channels of a cluster of tubule and tubule-associated cells. Scale bar: 0.1 mm. (C)Schematic of planarianprotonephridial cell types: flame (yellow), tubule (yellow and green) and tubule-associated (magenta) cells. Dotted box indicates area magnified andshown in detail below with a flame cell and tubule cells. (D)Transmission electron microscopy (TEM) of a flame cell showing fenestrations (fe),microvilli (mi), cilia (ci) and basal bodies (bb). Scale bar: 500 nm. (E)TEM of two interdigitated tubule cells (t1 and t2). Scale bar: 500 nm.(F,G)Double colorimetric in situ hybridization of a wild-type animal using a CA (red, INT/BCIP) and POU2/3 or Six1/2-2 (purple, BCIP/NBT) RNAprobes (F,G upper panels). Anterior, left. p, photoreceptors; px, pharynx. Scale bars: 0.1 mm. Insets: higher magnification of boxed area. POU2/3 andCA expression do not colocalize, whereas Six1/2-2 and CA expression do colocalize (>10 animals were similar). Lower panels: FISH of a wild-type day6 regenerating blastema using RNA probes for POU2/3 or Six1/2-2 and for tubules (cubilin) and tubule-associated cells (CA). Arrowheads identifydouble-labeled cells. (H,I)Labeling of wild-type animals using POU2/3 (H) and Six1/2-2 (I) RNA probes and an anti-acetylated tubulin antibody (blue).POU2/3 expression colocalizes with the acetylated-tubulin pattern, whereas Six1/2-2 expression does not. Scale bars: 0.05 mm.
DEVELO
PMENT

4390
expressed in tubule cells, see below) but not with the CA RNAprobe (Fig. 1F,H). By contrast, Six1/2-2-expressing cells co-expressed CA but were not labeled with the anti-acetylated tubulinantibody or the cubilin RNA probe (Fig. 1G,I). These expressionpatterns were observed in intact animals and in day 6 regeneratingblastemas (Fig. 1F,G lower panels; see Fig. S3E in thesupplementary material). RNAi reduced detectable correspondingmRNAs (see Fig. S3F,G in the supplementary material). Weconclude that Six1/2-2 is expressed in tubule-associated cells andPOU2/3 is expressed in tubule cells of the planarianprotonephridia.
Six1/2-2, POU2/3 and hunchback are required forformation and maintenance of protonephridiaTo assess the roles of Six1/2-2, POU2/3 and hunchback inplanarian protonephridia regeneration, RNAi animals wereamputated to remove heads and tails, and regenerating blastemas(unpigmented outgrowths at wound sites) were analyzed. Six1/2-2,POU2/3 and hunchback were essential for formation of the entireprotonephridial system during regeneration; too few flame(acetylated tubulin+), tubule (acetylated tubulin+) or tubule-associated (CA+) cells were regenerated when any of these geneswas inactivated (Fig. 2A). Inhibition of Six1/2-2, POU2/3 andhunchback specifically affected the nephridial system, with otherciliated cells remaining unaffected (see Fig. S4 in thesupplementary material). The requirement for Six1/2-2, POU2/3
and hunchback for protonephridia regeneration provides anexplanation for the animal bloating, blistering and lysis phenotypeobserved following RNAi of these genes.
RNAi animals undergoing tissue turnover (without amputation)were examined to assess gene requirements in protonephridiamaintenance. Tubule cells (acetylated tubulin+), which normallyexpress POU2/3, were gradually lost following POU2/3 RNAi;tubule-associated cells (CA+) were also lost, but at a slower rate(Fig. 2B). hunchback RNAi resulted in a similar pattern of cell loss(Fig. 2B). Conversely, tubule-associated cells, which normallyexpress Six1/2-2, were rapidly lost in Six1/2-2(RNAi) animals, withloss of tubule cells occurring more slowly (Fig. 2B). Six1/2-2,POU2/3 and hunchback are, therefore, required for protonephridiamaintenance. Requirements for mature cell viability and/orproduction of new differentiated cells during natural tissue turnovercould contribute to these phenotypes. In the case of Six1/2-2 RNAi,tubule-associated cells are lost faster than their normal tissue-turnover rate [e.g. irradiated animals lacking neoblasts do not showsigns of rapid protonephridia loss (Reddien et al., 2005b)],indicating a requirement for maintenance of existing cells for (atleast) this gene. In POU2/3 and hunchback RNAi animals at earlytime points, flame cells aggregated (Fig. 2B); lack of acetylatedtubulin+ signal for tubules in these RNAi animals is, therefore,suggestive of tubule loss rather than simply cilia loss. Additionalmarkers (see below) indicate further that it is not only cilia that arelost from protonephridia in these RNAi animals. Together, these
RESEARCH ARTICLE Development 138 (20)
Fig. 2. POU2/3, hunchback and Six1/2-2 are required for protonephridia regeneration and maintenance. (A)Six1/2-2, POU2/3 andhunchback RNAi animals failed to regenerate protonephridia. Left, cartoon of amputation and regeneration; blastemas are white. Dotted boxindicates the area shown in images. Numbers of tubule branches and tubule-associated cells per cluster were determined in regenerating blastemas(day 7 shown). (B)Fluorescence in situ hybridization and immunolabeling of control, POU2/3, hunchback or Six1/2-2 intact RNAi animals using a CA(magenta) RNA probe and an anti-acetylated tubulin antibody (yellow). Animals were analyzed at early (day 10 for Six1/2-2 and day 18 for POU2/3and hunchback) and late (day 14 for Six1/2-2 and day 26 for POU2/3 and hunchback) time points following RNAi initiation. Cartoon indicates theregion displayed in images and quantified in graphs. Anterior, left. Scale bars: 0.05 mm. Graphs display numbers of CA+ cells and tubule branches(Ac tubulin+) per cluster in tail regions. Data are mean ± s.e.m. (>6 animals per group). One-way ANOVA tests were performed, followed by aDunnet post-hoc test, comparing different groups relative to the control. ***P<0.0001. D
EVELO
PMENT

RNAi data demonstrate a requirement for Six1/2-2, POU2/3 andhunchback in both regeneration and maintenance of planarianprotonephridia.
Genes associated with protein clearance and iontransport are expressed in planarianprotonephridiaVertebrate homologs of Smed-Six1/2-2 and Smed-POU2/3 areessential for normal kidney development in mice and zebrafish(Hauptmann and Gerster, 2000; Nakai et al., 2003; Xu et al., 2003;Self et al., 2006; Kobayashi et al., 2008; Dressler, 2009). This raisesthe possibility that animal excretory systems utilize regulatoryprograms with a common evolutionary origin for theirdevelopment. If the similarities between planarian protonephridiaand the vertebrate kidney reflect homology, then homologs of othervertebrate genes involved in kidney function should be expressed inthe planarian nephridia. In order to investigate this possibility,microarray experiments with Six1/2-2 and POU2/3 RNAi animalswere performed (Fig. 3A). Because protonephridial cells becamedepleted over time in intact Six1/2-2 and POU2/3 RNAi animals, wereasoned that RNA from these animals should be depleted ofmRNAs expressed specifically in protonephridia. Numerous genesdisplayed decreased expression in RNAi animals compared with thecontrol. For example, expression of 736 and 351 genes was reducedto less than 50% of the control expression level in Six1/2-2(RNAi)(day 15 following RNAi) and POU2/3(RNAi) (day 21 post-RNAi)animals, respectively, with 208 of these genes in both lists.
Several genes with significant loss of expression in Six1/2-2 orPOU2/3 RNAi animals encode proteins associated with ion transportand protein clearance, and other genes encode hydrolytic enzymesand enzymes involved in the maintenance of acid-base balance (seeTables S1 and S2 in the supplementary material). These genesencode alkaline phosphatase (Smed-alkaline phosphatase-1),carbonic anhydrase (Smed-carbonic anhydrase, described above),cubilin (Smed-cubilin), plastin (Smed-plastin) and solute carrierproteins (Smed-HCO3– transporter, Smed-Na+/H+ exchanger andSmed-Na+/Ca2+ exchanger; see below for more information onSmed-Na+/Ca2+ exchanger), and were expressed specifically intubule or tubule-associated cells (Fig. 3B). These predicted proteinsare homologous to proteins present in the vertebrate kidney(Christensen and Birn, 2002; Delanote et al., 2005; Purkerson andSchwartz, 2007; El-Sheikh et al., 2008; Lisowska-Myjak, 2010). Inconclusion, planarian protonephridia and the vertebrate kidney sharesimilarities in addition to developmental roles for Six1/2 and POU2/3genes: they both possess a cell type for fluid filtration (flame cellsand podocytes), and the planarian tubule/tubule-associated cells andthe vertebrate kidney tubule cells utilize a similar set of excretoryfluid-processing proteins.
Identification of novel excretory system genesand a second tubule-associated cell typeTwo genes identified in the microarray experiments, Smed-Na+/Ca2+
exchanger (encoding a solute carrier protein) and Smed-ring finger-1 (abbreviated to Smed-RNF-1; encoding a novel ring finger protein),
4391RESEARCH ARTICLEPlanarian nephridia regeneration
Fig. 3. Microarray experiments identify homologs of vertebrate kidney genes expressed in S. mediterranea protonephridia.(A)Schematic of microarray experiments. RNA was collected as tubule-associated cells (purple) and tubule cells (green) were lost from Six1/2-2 orPOU2/3 RNAi animals. (B-D)Cartoon depicts region imaged. Fluorescence in situ hybridization and immunolabeling of a wild-type animal (tailregion shown) using different candidate genes, carbonic anhydrase (CA) and cubilin RNA probes, and an anti-acetylated tubulin antibody (Actubulin). Anterior, up. Scale bars: 0.05 mm. >10 animals per group were similar. Insets with higher magnification show colocalization (arrowheads). D
EVELO
PMENT

4392
were co-expressed in cells associated spatially with protonephridia.Expression of the Smed-Na+/Ca2+ exchanger gene was absent inregenerating Six1/2-2 and POU2/3 RNAi animals (see Fig. S5A inthe supplementary material). Some CA+ tubule-associated cells alsoexpressed Smed-Na+/Ca2+ exchanger; however, most did not (Fig.3C). These data suggest that a second tubule-associated cell typeexists in the planarian protonephridia (Fig. 3C).
Additional genes identified from the microarray experiments havehomologs in vertebrates, but do not have well-characterized roles inexcretory systems. A myelodysplasia/myeloid leukemia factorhomologous gene, Smed-myeloid leukemia factor (Smed-MLF), wasexpressed in tubule cells (Fig. 3D). Human MLF1 was identifiedfrom a translocation associated with myodysplastic syndrome andacute myeloid leukemia (Hitzler et al., 1999) and its biochemicalfunction is poorly understood. Smed-receptor protein tyrosinephosphatase-1 (Smed-RPTP-1; encoding a receptor tyrosinephosphatase) and Smed-EF-hand domain-containing 1 (Smed-EFHC1; encoding an EF-hand containing-1-like protein) were alsoexpressed in tubule cells (Fig. 3D). Mutations in vertebrate EFHC1have been demonstrated in some forms of human epilepsy and mightbe associated with cilia biology (Suzuki et al., 2004; Ikeda et al.,2005). The identification of Smed-RNF-1, Smed-MLF, Smed-RPTP-1 and Smed-EFHC-1 highlights how the planarian protonephridiacan be utilized to identify novel excretory organ genes.
Eya and Sall are required for protonephridiaformation and maintenance and Osr is expressedin tubule cellsThe microarray experiments described above also identifiedhomologs of transcriptional regulators. RNAi of an eyes absent gene,Smed-Eya (abbreviated to Eya; see Fig. S2D in the supplementary
material), decreased numbers of tubule and tubule-associated cells inblastemas (Fig. 4A). Moreover, intact animals under Eya RNAiconditions lost flame, tubule and tubule-associated cells (see Fig.S5B in the supplementary material). No Eya expression in matureprotonephridia cells was detected; however, Eya is expressed in eyeand parenchymal cells in planarians (Lapan and Reddien, 2011;Mannini et al., 2004) and Eya expression was detected duringprotonephridia regeneration (see below). Eya knockout mice lackkidneys (Xu et al., 1999). Eya acts together with sine oculis duringeye formation in D. melanogaster as well as in the organogenesis ofseveral other tissues (Bonini et al., 1993; Cheyette et al., 1994; Li etal., 2003). Our results suggest that Smed-Six1/2-2 and Smed-Eyamight act together in planarian protonephridia regeneration.
A second candidate regulatory gene, Smed-Sall (abbreviated toSall) (see Fig. S2E in the supplementary material), which encodesa transcription factor similar to Drosophila Spalt (Salm – FlyBase),was expressed in tubule cells in day six regenerating blastemas(Fig. 4B). Sall(RNAi) intact and regenerating animals bloated (Fig.4C; see Fig. S5C in the supplementary material). BloatedSall(RNAi) animals showed a severe reduction of tubule cells withan attendant increased number of CA-expressing cells (Fig. 4C).One possible explanation for this phenotype is that Sall might actin tubule cell formation/maintenance to repress the tubule-associated cell fate. Sall1-deficient mice do not form kidneys(Nishinakamura et al., 2001).
Smed-Osr (abbreviated to Osr) is homologous to the Drosophilaodd skipped gene (see Fig. S2F in the supplementary material) andwas also expressed in the planarian protonephridial tubule cells(Fig. 4B). A vertebrate Osr family gene, Osr1, is essential fornephron progenitor development (Mugford et al., 2008) and osr1inactivation causes loss of kidney progenitors in zebrafish and a
RESEARCH ARTICLE Development 138 (20)
Fig. 4. Eya and Sall are required for protonephridia regeneration and Osr is expressed in tubule cells. (A)Eya(RNAi) animals failed toregenerate protonephridia. Cartoon depicts region analyzed. Left: numbers of tubule branches (Ac tubulin+) and tubule-associated cells (CA+) percluster in control and Eya RNAi animals six days following amputation, 15 days following RNAi initiation. Data are mean ± s.e.m. (>12 animals pergroup). ***P<0.0001 determined by Student’s t-test. Right: fluorescence in situ hybridization and immunolabeling of a regenerating control or EyaRNAi animal (tail blastema shown; anterior, left) using the CA (magenta) RNA probe and an anti-acetylated tubulin antibody (yellow). Scale bars:0.05 mm. (B)Sall and Osr are expressed in tubule cells. Cartoon depicts blastema region imaged. Anterior, up. Scale bars: 0.05 mm. Insets on rightshow colocalization of transcription regulatory gene RNA (magenta) and cubilin (green). (C)Top: Sall(RNAi) animals bloated during tissue turnover inintact animals (17/20 animals bloated between days 43 and 46 after initial RNAi). Dotted box indicates the area depicted below. Middle: A bloatedSall(RNAi) animal with reduced tubule (Ac tubulin+, yellow) and increased tubule-associated cell numbers (CA+, magenta) is shown. Arrows indicateexample clusters of ectopic tubule-associated cells at the animal periphery. Bottom: Higher magnification images of control and Sall(RNAi) clusters.Anterior, left. Scale bars: 0.05 mm.
DEVELO
PMENT
subsequent absence of kidney tubules (Tena et al., 2007;Mudumana et al., 2008). Similarly, Osr1-knockout mice do notform the metanephric mesenchyme and, therefore, lack kidneys(Wang et al., 2005; James et al., 2006). We did not detect aphenotype for Smed-Osr(RNAi) animals, but the expression of Osrin protonephridia and roles for vertebrate Osr1 in kidneydevelopment indicate further similarity between transcriptionfactors involved in animal excretory systems. The microarrayresults expand the number of identified transcriptional regulatoryfactors associated with protonephridia regeneration to six: Six1/2-2, POU2/3, hunchback, Eya, Sall and Osr.
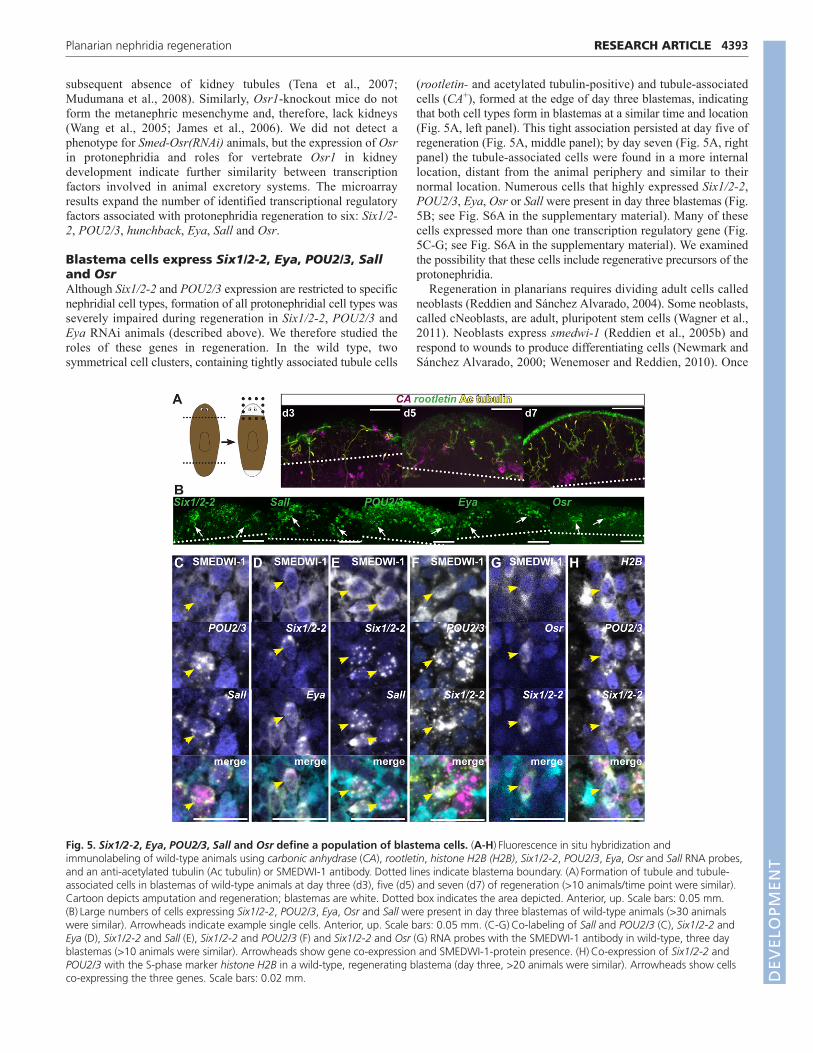
Blastema cells express Six1/2-2, Eya, POU2/3, Salland OsrAlthough Six1/2-2 and POU2/3 expression are restricted to specificnephridial cell types, formation of all protonephridial cell types wasseverely impaired during regeneration in Six1/2-2, POU2/3 andEya RNAi animals (described above). We therefore studied theroles of these genes in regeneration. In the wild type, twosymmetrical cell clusters, containing tightly associated tubule cells
(rootletin- and acetylated tubulin-positive) and tubule-associatedcells (CA+), formed at the edge of day three blastemas, indicatingthat both cell types form in blastemas at a similar time and location(Fig. 5A, left panel). This tight association persisted at day five ofregeneration (Fig. 5A, middle panel); by day seven (Fig. 5A, rightpanel) the tubule-associated cells were found in a more internallocation, distant from the animal periphery and similar to theirnormal location. Numerous cells that highly expressed Six1/2-2,POU2/3, Eya, Osr or Sall were present in day three blastemas (Fig.5B; see Fig. S6A in the supplementary material). Many of thesecells expressed more than one transcription regulatory gene (Fig.5C-G; see Fig. S6A in the supplementary material). We examinedthe possibility that these cells include regenerative precursors of theprotonephridia.
Regeneration in planarians requires dividing adult cells calledneoblasts (Reddien and Sánchez Alvarado, 2004). Some neoblasts,called cNeoblasts, are adult, pluripotent stem cells (Wagner et al.,2011). Neoblasts express smedwi-1 (Reddien et al., 2005b) andrespond to wounds to produce differentiating cells (Newmark andSánchez Alvarado, 2000; Wenemoser and Reddien, 2010). Once
4393RESEARCH ARTICLEPlanarian nephridia regeneration
Fig. 5. Six1/2-2, Eya, POU2/3, Sall and Osr define a population of blastema cells. (A-H)Fluorescence in situ hybridization andimmunolabeling of wild-type animals using carbonic anhydrase (CA), rootletin, histone H2B (H2B), Six1/2-2, POU2/3, Eya, Osr and Sall RNA probes,and an anti-acetylated tubulin (Ac tubulin) or SMEDWI-1 antibody. Dotted lines indicate blastema boundary. (A)Formation of tubule and tubule-associated cells in blastemas of wild-type animals at day three (d3), five (d5) and seven (d7) of regeneration (>10 animals/time point were similar).Cartoon depicts amputation and regeneration; blastemas are white. Dotted box indicates the area depicted. Anterior, up. Scale bars: 0.05 mm.(B)Large numbers of cells expressing Six1/2-2, POU2/3, Eya, Osr and Sall were present in day three blastemas of wild-type animals (>30 animalswere similar). Arrowheads indicate example single cells. Anterior, up. Scale bars: 0.05 mm. (C-G)Co-labeling of Sall and POU2/3 (C), Six1/2-2 andEya (D), Six1/2-2 and Sall (E), Six1/2-2 and POU2/3 (F) and Six1/2-2 and Osr (G) RNA probes with the SMEDWI-1 antibody in wild-type, three dayblastemas (>10 animals were similar). Arrowheads show gene co-expression and SMEDWI-1-protein presence. (H)Co-expression of Six1/2-2 andPOU2/3 with the S-phase marker histone H2B in a wild-type, regenerating blastema (day three, >20 animals were similar). Arrowheads show cellsco-expressing the three genes. Scale bars: 0.02 mm. D
EVELO
PMENT

4394
neoblasts stop dividing and initiate differentiation, smedwi-1mRNA disappears; SMEDWI-1 protein, however, persists indifferentiating neoblast progeny cells for several days (Guo et al.,2006; Scimone et al., 2010). Therefore, an antibody recognizingSMEDWI-1 labels neoblasts and newly made neoblast progeny,including for differentiating cells of the nervous system andintestine (Wagner et al., 2011). We used this SMEDWI-1 antibodyto describe a population of blastema cells with extensive co-expression of the protonephridia transcription factors. MostPOU2/3+ cells in day three blastemas co-expressed Sall (107/120POU2/3+ cells were Sall+) (see Fig. S6A in the supplementarymaterial) and frequently had detectable SMEDWI-1 protein (55/70POU2/3+/Sall+ cells were SMEDWI-1+), indicating that these cellsare newly made neoblast progeny cells (Fig. 5C). In day sixblastemas, both POU2/3 and Sall were expressed in tubule cells(Fig. 4C; see Fig. S3E in the supplementary material); therefore,day three blastema cells co-expressing POU2/3 and Sall couldrepresent precursor cells of protonephridial tubules (see below).Several Six1/2-2+ cells co-expressed Eya (146/301 Six1/2-2+ cellswere Eya+) and these cells frequently had SMEDWI-1 (97/104Six1/2-2+/Eya+ cells were SMEDWI-1+) (Fig. 5D). Surprisingly,considering the different expression sites for Six1/2-2 and Sall inday six blastemas (Fig. 4C; see Fig. S3E in the supplementarymaterial), many Six1/2-2+/Sall+ double-positive cells existed in daythree blastemas (85/91 Six1/2-2+ cells were Sall+) (see Fig. S6A inthe supplementary material). Most of these cells also possessedSMEDWI-1 protein (52/65 Six1/2-2+/Sall+ cells were SMEDWI-1+) (Fig. 5E). Furthermore, we found colocalization of Six1/2-2 andPOU2/3 mRNAs themselves in day three blastemas (187/275Six1/2-2+ cells were POU2/3+) (see Fig. S6A in the supplementarymaterial). Many Six1/2-2+/POU2/3+ cells also possessedSMEDWI-1 (36/57 Six1/2-2+/POU2/3+ cells were SMEDWI-1+)(Fig. 5F). Cells also co-expressed Six1/2-2 and Osr in day threeblastemas (see Fig. S6A in the supplementary material) and someof these cells were SMEDWI-1+ (Fig. 5G). We also found cells thatco-express Osr and POU2/3 or Osr and Sall, together withSMEDWI-1 (see Fig. S6B in the supplementary material).
The significant co-expression of these transcription factor genes(pair-wise tests) indicates the existence of blastema cells that co-express Six1/2-2, Eya, POU2/3 and Sall, with some cells alsoexpressing Osr. For example, the likelihood that Sall+/POU2/3+
and Six1/2-2+/POU2/3+ populations overlap in day three blastemasis significant (Fisher’s test, P<0.0001); similarly, the likelihood thatSix1/2-2+/POU2/3+ and Six1/2-2+/Eya+ cell populations overlap issignificant (Fisher’s test, P0.0029). These cells are newlyproduced (SMEDWI+) and present in early blastemas followingamputation. Cells co-expressing the transcription factors POU2/3and Six1/2-2 or Six1/2-2 and Sall were not detected in day threeblastemas of animals that were lethally irradiated the day afteramputation (see Fig. S6C in the supplementary material), furtherindicating that these cells represent a transient cell populationdetected prominently during regeneration.
We next assessed whether expression of nephridia transcriptionfactors initiates within neoblasts. Cells that co-express eachtranscription factor tested and the neoblast marker smedwi-1 wereindeed present (see Fig. S6D in the supplementary material).Moreover, cells co-expressing POU2/3, Six1/2-2 and the S-phasemarker histone H2B (H2B) (which labels neoblasts, as the onlydividing somatic cell population) were observed in day threeblastemas (Fig. 5H). These results suggest that initial specificationof candidate nephridial precursor cells occurs within neoblasts.Similarly, eye precursor cells originate within neoblasts during
regeneration (Lapan and Reddien, 2011). Examination ofindividual neoblasts isolated by flow cytometry (neoblasts can beisolated by DNA labeling because of their greater than 2N DNAcontent and are called X1 cells), has previously revealed geneexpression heterogeneity in dividing planarian cells (Hayashi et al.,2010), and we detected some X1 cells expressing POU2/3 fromday three blastemas (see Fig. S6E in the supplementary material).Whereas some neoblasts are pluripotent (Wagner et al., 2011), theseeye data (Lapan and Reddien, 2011) and nephridia data togetherdemonstrate that gene expression heterogeneity of neoblasts existsin vivo with some neoblasts expressing lineage-specific genes. It isunknown whether these nephridia transcription factor+ neoblastswill divide to expand in number or directly differentiate.
Next, we reasoned that if blastema cells expressing Sall, Six1/2-2, Eya, Osr and POU2/3 are indeed precursor cells forprotonephridia, there might exist cells in intermediate stagesbetween precursors and differentiated cells that co-express both thetranscription factors and the differentiated markers. We found thatsome of the cells co-expressing POU2/3 and SMEDWI-1, Six1/2-2 and SMEDWI-1, or Sall and SMEDWI-1 also expressed lowlevels of cubilin (Fig. 6A), which is specifically expressed inprotonephridial tubule cells (Fig. 3B). Similarly, some cells co-expressing Six1/2-2 and SMEDWI-1 also expressed low levels ofCA, the tubule-associated cell marker (Fig. 6A). These resultsfurther support the hypothesis that Sall, Six1/2-2, POU2/3, and byassociation (because of co-expression data above) Eya and Osr,label blastema cells that, as a population, will produce tubule andtubule-associated cells.
Six1/2-2 and POU2/3 RNAi animals fail to generateprecursor cellsIf the population of Six1/2-2+, POU2/3+, Eya+, Osr+ and Sall+ day3 blastema cells does indeed include precursors for tubule andtubule-associated cells, ablation of this cell population should blockboth differentiation pathways. We therefore assessed the presenceof the candidate nephridial precursors in Six1/2-2 and POU2/3RNAi animals. As expected, no tubule (rootletin+) or tubule-associated cells (CA+) were found in Six1/2-2 and POU2/3 RNAiregeneration blastemas (Fig. 6B). Furthermore, no POU2/3+/Sall+
blastema cells were found in Six1/2-2(RNAi) animals and noSix1/2-2+/Sall+ blastema cells were found in POU2/3(RNAi)animals (Fig. 6B). In fact, few blastema cells expressing Six1/2-2or POU2/3 were present at all following RNAi of either gene.These results support a model in which Six1/2-2 and POU2/3functions are essential for the generation of a population ofnephridial precursor cells, with some cells of this populationdifferentiating into tubule cells and others differentiating intotubule-associated cells. The absence of such a population ofprecursors would explain the failure to regenerate both tubule andtubule-associated cells in these RNAi animals.
DISCUSSIONSix1/2-2, POU2/3, hunchback, Eya, Sall and Osr areinvolved in regeneration of an excretory systemThe ability to regenerate missing body parts is a phenomenon thatis broadly distributed in multicellular organisms. However, themechanisms underlying this phenomenon are not well understood.Because planarians can regenerate all adult organs and their genescan be efficiently inhibited by RNAi, planarians can be utilized toidentify mechanisms controlling regeneration of numerous celltypes. We investigated genes consist of in regeneration of theplanarian excretory system, called protonephridia. The S.
RESEARCH ARTICLE Development 138 (20)
DEVELO
PMENT

mediterranea protonephridia consist of numerous tubules, twotypes of tubule-associated cells, and ciliated terminal cells. Becausethis organ system is dispersed throughout planarian bodies, mostinjuries will remove or damage regions of the protonephridia.Adult planarians must, therefore, possess mechanisms for makingeach protonephridial cell type and for integrating new cells with theremaining tubule system.
We identified multiple genes (Six1/2-2, POU2/3, hunchback,Eya, Sall and Osr) encoding conserved transcription regulatoryproteins that are involved in protonephridia regeneration. Six1/2-2,POU2/3, hunchback and Eya were required for regeneration of allprotonephridial cells, and Sall was required for regeneration of theproper ratio of tubule to tubule-associated cells. Duringregeneration, a population of blastema cells co-expressed many ofthese transcription regulatory genes. These blastema cells werenewly produced neoblast progeny (SMEDWI protein+ andirradiation-sensitive). Some transcription regulatorygene+/SMEDWI-1+ blastema cells co-expressed differentiatednephridial markers. Finally, RNAi of Six1/2-2 or POU2/3eliminated this population of blastema cells and blockedregeneration of differentiated protonephridial cells. Based on theseobservations, we propose a regenerative model for protonephridia(Fig. 7). First, protonephridia transcription factors are expressedtogether in neoblasts (some Six1/2-2 and POU2/3 double-positivecells were co-labeled with smedwi or H2B mRNA). Next, neoblastprogeny (SMEDWI+) that co-express Six1/2-2, POU2/3, Eya, Salland Osr in the blastema are produced (Fig. 7A), requiring Six1/2-2 and POU2/3 for formation. Cells from within this populationproduce terminally differentiated cells, either a tubule cellexpressing POU2/3, Osr and Sall and the markers rootletin and
cubilin, or a tubule-associated cell expressing Six1/2-2 and CA (Fig.7A). These precursor cells are described as a population and could,therefore, include multipotent nephridial precursors or couldpossess distinct lineage-restricted cells with one subset of cellsmaking tubules and a different subset making tubule-associatedcells. Because of the lack of a planarian flame cell marker, there isno direct evidence that the described blastema cells can producecells that differentiate into flame cells. However, flame cells are notregenerated in either Six1/2-2 or POU2/3 RNAi animals. Duringregeneration, newly formed cells in the blastema combine withremaining pre-existing protonephridia cells in the remainder of thebody to reconstitute the excretory system.
In vertebrates, the existence of adult renal progenitors iscontroversial (Anglani et al., 2010; Guo and Cantley, 2010), and noadult, multipotent renal stem cell has been isolated (Guo and Cantley,2010). However, cells with long-term self-renewal capacity in vitroand expression of renal transcription factors such as Cited1, Wt1,Osr1, Six1, Six2, Sall, Eya and Hoxa11 have been identified in theembryonic metanephric mesenchyme (Osafune et al., 2006; Guo andCantley, 2010; Lusis et al., 2010). In addition, single cells expressingSall isolated from the embryonic metanephric mesenchyme in miceformed colonies in vitro and differentiated into cells expressingpodocyte, proximal tubule, distal tubule and Henle’s loop markers(Osafune et al., 2006). Moreover, in vivo studies found that most ofthe mouse kidney cells arise from an Osr1-expressing population ofcells (James et al., 2006; Mugford et al., 2008) and that a capmesenchyme Six2+ population represents a multipotent progenitorcell population with the ability to differentiate into an epithelialglomerular and tubular cell (Boyle et al., 2008; Kobayashi et al.,2008). Therefore, dividing mesenchymal cells can promote nephron
4395RESEARCH ARTICLEPlanarian nephridia regeneration
Fig. 6. Differentiation and formation ofnephridia precursors. (A)Fluorescence in situhybridization (FISH) and immunolabeling usingcarbonic anhydrase (CA), cubilin, Six1/2-2, POU2/3and Sall RNA probes and the anti-SMEDWI-1antibody in wild-type, three-day regeneratingblastemas. Arrowheads show cells co-expressing atranscription factor, a differentiated marker and theSMEDWI-1 protein (>10 animals were similar). Scalebars: 0.02 mm. (B)Six1/2-2 and POU2/3 RNAianimals failed to generate nephridia precursors. FISHwith control, Six1/2-2 or POU2/3 RNAi animals (>20animals were similar). Dotted lines indicate blastemaboundary. Anterior, up. Scale bars: 0.05 mm.
DEVELO
PMENT

4396
development in vertebrates and parenchymal proliferating cells(neoblasts) promote protonephridia regeneration in planarians, withthese developmental and regenerative processes involving severalsimilar transcription factors.
A common transcriptional regulatory program fornephridia formation in bilateriansDiverse animals utilize organs for similar tasks. The discovery thatsimilar developmental genes are utilized in organisms across theBilateria (the bilaterally symmetrical animals) for the developmentof similar organs, such as Pax6/eyeless for the eye (Quiring et al.,1994), MyoD for muscle (Weintraub et al., 1991; Chen et al.,1992), Gata and Forkhead/HNF3 for the gut (Ang et al., 1993;Rehorn et al., 1996; Zhu et al., 1997) and tinman/Nkx2-5 for theheart (Evans et al., 1995), has raised the possibility that certainorgan regulatory programs existed in the common ancestor of theBilateria. However, no conserved gene regulatory paradigm has yetbeen established for animal excretory systems.
In vertebrates, the POU3-family member Brn-1 (Pou3f3) isexpressed in the developing kidney and is required for kidneytubule formation (Hauptmann and Gerster, 2000; Nakai et al.,2003). Six1, Eya1, Osr1 and Sall are expressed in the metanephricmesenchyme and are essential for early mammalian kidneydevelopment. Mice deficient for each of these genes are similar;they die prenatally or at birth and lack kidneys owing to a failureof ureteric bud outgrowth and metanephric induction resulting inmesenchyme apoptosis (Abdelhak et al., 1997; Xu et al., 1999;Nishinakamura et al., 2001; Xu et al., 2003; Sajithlal et al., 2005;Wang et al., 2005). Moreover, Six2-deficient mice also show severekidney hypoplasia because Six2 is required for function of anephron-progenitor population (Self et al., 2006; Kobayashi et al.,2008). The identification of a molecular program includingPOU2/3, Six1/2, Eya, Sall and Osr family genes involved in theformation of excretory systems in both planarians (protostomes)
and vertebrates (deuterostomes) suggests that a similar set of genesmight have specified excretory system cell development in thecommon ancestor of bilaterally symmetrical animals.
Similarity between characters of different animal groups couldreflect homology or could occur by independent evolution(Abouheif et al., 1997). If the roles for POU2/3, Six1/2, Eya, Salland Osr family genes in planarian and vertebrate excretory systemsreflect homology, cell types expressing or requiring these genes fortheir development should utilize similar proteins for theirdifferentiated functions. We found several genes encoding proteinsinvolved in protein clearance, acid-base balance and ion transportexpressed in the planarian protonephridia that are indeed similar tovertebrate kidney genes (Fig. 7B). A further prediction of thehomology hypothesis is that additional similarities would exist forexcretory systems in other organisms. Metanephridial andprotonephridial excretory systems are found in the three superphylacomprising bilaterians: deuterostomes, and the two groups amongprotostomes, the Ecdysozoa and the Lophotrochozoa (see Fig. S1in the supplementary material). Future molecular analyses inadditional organisms will, therefore, be an important direction forassessing potential homologous roles for the gene familiesdescribed here in excretory system development. The specializedecdysozoan excretory systems studied in C. elegans andDrosophila might reflect evolutionarily derived characters from theancestral condition for the Ecdysozoa. Nonetheless, the recentdiscovery that orthologous genes are involved in nephrocyte slitformation in Drosophila and mouse podocytes points to potentialevolutionary conservation of the functional morphology betweenwaste filtration cells in ecdysozoans and deuterostomes (Weaverset al., 2009). Furthermore, a C. elegans POU2/3 family gene, ceh-6, is essential for the function of the only excretory cell in thisanimal (Burglin and Ruvkun, 2001). Therefore, the hypothesis ofa homologous role in excretory systems for POU2/3 genes hassupport from all three bilaterian superphyla.
RESEARCH ARTICLE Development 138 (20)
Fig. 7. A genetic and cellular program for regeneration of the planarian protonephridia. (A)Model for protonephridia regeneration:neoblasts (dark blue, dividing cell expressing smedwi-1 and histone H2B) produce a transient population of nephridia precursor cells (light blue) thatco-expresses Six1/2-2, Eya, Osr, POU2/3, Sall and SMEDWI-1. Some cells expressing Six1/2-2, POU2/3 and Sall were found to express neoblast-specific H2B (S-phase marker) and/or smedwi-1 mRNA. Dotted box indicates that data cannot distinguish whether this precursor population ishomogeneous or heterogeneous in potential. Precursor cells differentiate either as a tubule cell (green), maintaining expression of POU2/3, Sall andOsr and expressing rootletin and cubilin, or as a tubule-associated cell (magenta), maintaining Six1/2-2 expression and expressing CA. (B)Schematicof the planarian protonephridial system. Genes expressed in either tubule (green and yellow), tubule-associated cells (magenta) or tubule-associatedcells-2 (orange) are listed.
DEVELO
PMENT

Some gene classes that we identified to be expressed in or to beimportant for formation of planarian nephridia have no known rolesin the vertebrate kidney, raising the possibility that a similar rolefor orthologous genes might exist in other organisms. Theseinclude a hunchback gene, an MLF gene, a conserved Ring finger-encoding gene, an EF-hand domain-containing-1 gene, and areceptor protein tyrosine phosphatase-1 gene. Study of planarianscan thus identify previously uncharacterized excretory systemproteins.
Our analysis of planarian protonephridia identifies genespromoting regeneration of a filtration-based tubule excretorysystem. Identification of central regulatory programs for cell types,and distinguishing the core factors from those with species-specificfunction, is facilitated by comparing functional data from differentorganisms. Comparing data from planarians and vertebratesindicates that Six1/2-2, Eya, POU2/3, Sall and Osr, among themany factors possible, are candidate central regulators of animalexcretory cell types. This could indicate candidate genes to utilizefor future attempts at reprogramming-based regenerative medicineapplications for kidney diseases. We propose that nephridia werepresent in the bilaterian ancestor with molecular mechanismsunderlying nephridia formation conserved over millions of years ofbilaterian evolution.
Note added in proofWhile this manuscript was in production, Rink et al. (Rink et al.,2011) published work demonstrating a role for an EGFR orthologin tubule branching in planarian nephridia. This manuscript is alsoa resource for anatomical markers and features of protonephridia.
AcknowledgementsWe thank E. Alvarez-Saavedra and members of the Reddien laboratory fordiscussions; Sylvain Lapan for sharing methods and advice for detectingprecursor cells; Danielle Wenemoser for help with the X1 in situ experiment;Irving Wang for illustrations; and N. Watson at the Whitehead, W. M. KeckMicroscopy Facility for electron microscopy.
FundingM.L.S. and M.S. were supported by Jane Coffin Childs Fellowships. P.W.R. is anearly career scientist of the Howard Hughes Medical Institute. Weacknowledge support by NIH R01GM080639 and Keck Foundation support,and a Cabot career development professorship. Deposited in PMC for releaseafter 6 months.
Competing interests statementThe authors declare no competing financial interests.
Supplementary materialSupplementary material for this article is available athttp://dev.biologists.org/lookup/suppl/doi:10.1242/dev.068098/-/DC1
ReferencesAbdelhak, S., Kalatzis, V., Heilig, R., Compain, S., Samson, D., Vincent, C.,
Weil, D., Cruaud, C., Sahly, I., Leibovici, M. et al. (1997). A humanhomologue of the Drosophila eyes absent gene underlies branchio-oto-renal(BOR) syndrome and identifies a novel gene family. Nat. Genet. 15, 157-164.
Abouheif, E., Akam, M., Dickinson, W. J., Holland, P. W., Meyer, A., Patel, N.H., Raff, R. A., Roth, V. L. and Wray, G. A. (1997). Homology anddevelopmental genes. Trends Genet. 13, 432-433.
Ang, S. L., Wierda, A., Wong, D., Stevens, K. A., Cascio, S., Rossant, J. andZaret, K. S. (1993). The formation and maintenance of the definitive endodermlineage in the mouse: involvement of HNF3/forkhead proteins. Development119, 1301-1315.
Anglani, F., Mezzabotta, F., Ceol, M., Cristofaro, R., Del Prete, D. andD’Angelo, A. (2010). The regenerative potential of the kidney: what can welearn from developmental biology? Stem Cell Rev. 6, 650-657.
Bartolomaeus, T. and Ax, P. (1992). Protonephridia and Metanephridia – theirrelation within the Bilateria. J. Zool. Syst. Evol. Res. 30, 21-45.
Bonini, N. M., Leiserson, W. M. and Benzer, S. (1993). The eyes absent gene:genetic control of cell survival and differentiation in the developing Drosophilaeye. Cell 72, 379-395.
Boyle, S., Misfeldt, A., Chandler, K. J., Deal, K. K., Southard-Smith, E. M.,Mortlock, D. P., Baldwin, H. S. and de Caestecker, M. (2008). Fate mappingusing Cited1-CreERT2 mice demonstrates that the cap mesenchyme containsself-renewing progenitor cells and gives rise exclusively to nephronic epithelia.Dev. Biol. 313, 234-245.
Burglin, T. R. and Ruvkun, G. (2001). Regulation of ectodermal and excretoryfunction by the C. elegans POU homeobox gene ceh-6. Development 128, 779-790.
Chen, L., Krause, M., Draper, B., Weintraub, H. and Fire, A. (1992). Body-wallmuscle formation in Caenorhabditis elegans embryos that lack the MyoDhomolog hlh-1. Science 256, 240-243.
Cheyette, B. N., Green, P. J., Martin, K., Garren, H., Hartenstein, V. andZipursky, S. L. (1994). The Drosophila sine oculis locus encodes ahomeodomain-containing protein required for the development of the entirevisual system. Neuron 12, 977-996.
Christensen, E. I. and Birn, H. (2002). Megalin and cubilin: multifunctionalendocytic receptors. Nat. Rev. Mol. Cell Biol. 3, 256-266.
Costantini, F. and Kopan, R. (2010). Patterning a complex organ: branchingmorphogenesis and nephron segmentation in kidney development. Dev. Cell 18,698-712.
Delanote, V., Vandekerckhove, J. and Gettemans, J. (2005). Plastins: versatilemodulators of actin organization in (patho)physiological cellular processes. ActaPharmacol. Sinica 26, 769-779.
Denholm, B. and Skaer, H. (2009). Bringing together components of the fly renalsystem. Curr. Opin. Genet. Dev. 19, 526-532.
Dressler, G. R. (2006). The cellular basis of kidney development. Annu. Rev. CellDev. Biol. 22, 509-529.
Dressler, G. R. (2009). Advances in early kidney specification, development andpatterning. Development 136, 3863-3874.
El-Sheikh, A. A., Masereeuw, R. and Russel, F. G. (2008). Mechanisms of renalanionic drug transport. Eur. J. Pharmacol. 585, 245-255.
Evans, S. M., Yan, W., Murillo, M. P., Ponce, J. and Papalopulu, N. (1995).tinman, a Drosophila homeobox gene required for heart and visceral mesodermspecification, may be represented by a family of genes in vertebrates: XNkx-2.3,a second vertebrate homologue of tinman. Development 121, 3889-3899.
Glazer, A. M., Wilkinson, A. W., Backer, C. B., Lapan, S. W., Gutzman, J. H.,Cheeseman, I. M. and Reddien, P. W. (2010). The Zn finger protein Iguanaimpacts Hedgehog signaling by promoting ciliogenesis. Dev. Biol. 337, 148-156.
Guo, J. K. and Cantley, L. G. (2010). Cellular maintenance and repair of thekidney. Annu. Rev. Physiol. 72, 357-376.
Guo, T., Peters, A. H. and Newmark, P. A. (2006). A Bruno-like gene is requiredfor stem cell maintenance in planarians. Dev. Cell 11, 159-169.
Hauptmann, G. and Gerster, T. (2000). Combinatorial expression of zebrafishBrn-1- and Brn-2-related POU genes in the embryonic brain, pronephricprimordium, and pharyngeal arches. Dev. Dyn. 218, 345-358.
Hayashi, T., Shibata, N., Okumura, R., Kudome, T., Nishimura, O., Tarui, H.and Agata, K. (2010). Single-cell gene profiling of planarian stem cells usingfluorescent activated cell sorting and its ‘index sorting’ function for stem cellresearch. Dev. Growth Differ. 52, 131-144.
Hertel, L. A. (1993). Excretion and osmoregulation in the flatworms. Trans. Am.Microsc. Soc. 112, 10-17.
Hitzler, J. K., Witte, D. P., Jenkins, N. A., Copeland, N. G., Gilbert, D. J.,Naeve, C. W., Look, A. T. and Morris, S. W. (1999). cDNA cloning, expressionpattern, and chromosomal localization of Mlf1, murine homologue of a geneinvolved in myelodysplasia and acute myeloid leukemia. Am. J. Pathol. 155, 53-59.
Ikeda, T., Ikeda, K., Enomoto, M., Park, M. K., Hirono, M. and Kamiya, R.(2005). The mouse ortholog of EFHC1 implicated in juvenile myoclonic epilepsyis an axonemal protein widely conserved among organisms with motile cilia andflagella. FEBS Lett. 579, 819-822.
James, R. G., Kamei, C. N., Wang, Q., Jiang, R. and Schultheiss, T. M. (2006).Odd-skipped related 1 is required for development of the metanephric kidneyand regulates formation and differentiation of kidney precursor cells.Development 133, 2995-3004.
Kobayashi, A., Valerius, M. T., Mugford, J. W., Carroll, T. J., Self, M., Oliver,G. and McMahon, A. P. (2008). Six2 defines and regulates a multipotent self-renewing nephron progenitor population throughout mammalian kidneydevelopment. Cell Stem Cell 3, 169-181.
Lapan, S. W. and Reddien, P. W. (2011). dlx and sp6-9 control optic cupregeneration in a prototypic eye. PLoS Genetics 7, e1002226.
Li, X., Oghi, K. A., Zhang, J., Krones, A., Bush, K. T., Glass, C. K., Nigam, S.K., Aggarwal, A. K., Maas, R., Rose, D. W. et al. (2003). Eya proteinphosphatase activity regulates Six1-Dach-Eya transcriptional effects inmammalian organogenesis. Nature 426, 247-254.
Lisowska-Myjak, B. (2010). Serum and urinary biomarkers of acute kidney injury.Blood Purif. 29, 357-365.
4397RESEARCH ARTICLEPlanarian nephridia regeneration
DEVELO
PMENT

4398
Lusis, M., Li, J., Ineson, J., Christensen, M. E., Rice, A. and Little, M. H.(2010). Isolation of clonogenic, long-term self renewing embryonic renal stemcells. Stem Cell Res. 5, 23-39.
Mannini, L., Rossi, L., Deri, P., Gremigni, V., Salvetti, A., Salo, E. andBatistoni, R. (2004). Djeyes absent (Djeya) controls prototypic planarian eyeregeneration by cooperating with the transcription factor Djsix-1. Dev. Biol. 269,346-359.
McKanna, J. A. (1968a). Fine structure of the protonephridial system in Planaria. I.Flame cells. Z. Zellforsch. Mikrosk. Anat. 92, 509-523.
McKanna, J. A. (1968b). Fine structure of the protonephridial system in Planaria.II. Ductules, collecting ducts, and osmoregulatory cells. Z. Zellforsch. Mikrosk.Anat. 92, 524-535.
Mudumana, S. P., Hentschel, D., Liu, Y., Vasilyev, A. and Drummond, I. A.(2008). Odd skipped related1 reveals a novel role for endoderm in regulatingkidney versus vascular cell fate. Development 135, 3355-3367.
Mugford, J. W., Sipila, P., McMahon, J. A. and McMahon, A. P. (2008). Osr1expression demarcates a multi-potent population of intermediate mesodermthat undergoes progressive restriction to an Osr1-dependent nephron progenitorcompartment within the mammalian kidney. Dev. Biol. 324, 88-98.
Nakai, S., Sugitani, Y., Sato, H., Ito, S., Miura, Y., Ogawa, M., Nishi, M.,Jishage, K., Minowa, O. and Noda, T. (2003). Crucial roles of Brn1 in distaltubule formation and function in mouse kidney. Development 130, 4751-4759.
Nelson, F. K., Albert, P. S. and Riddle, D. L. (1983). Fine structure of theCaenorhabditis elegans secretory-excretory system. J. Ultrastruct. Res. 82, 156-171.
Newmark, P. A. and Sánchez Alvarado, A. (2000). Bromodeoxyuridinespecifically labels the regenerative stem cells of planarians. Dev. Biol. 220, 142-153.
Nishinakamura, R., Matsumoto, Y., Nakao, K., Nakamura, K., Sato, A.,Copeland, N. G., Gilbert, D. J., Jenkins, N. A., Scully, S., Lacey, D. L. et al.(2001). Murine homolog of SALL1 is essential for ureteric bud invasion in kidneydevelopment. Development 128, 3105-3115.
Osafune, K., Takasato, M., Kispert, A., Asashima, M. and Nishinakamura, R.(2006). Identification of multipotent progenitors in the embryonic mouse kidneyby a novel colony-forming assay. Development 133, 151-161.
Pearson, B. J., Eisenhoffer, G. T., Gurley, K. A., Rink, J. C., Miller, D. E. andSánchez Alvarado, A. (2009). Formaldehyde-based whole-mount in situhybridization method for planarians. Dev. Dyn. 238, 443-450.
Purkerson, J. M. and Schwartz, G. J. (2007). The role of carbonic anhydrases inrenal physiology. Kidney Int. 71, 103-115.
Quaggin, S. E. and Kreidberg, J. A. (2008). Development of the renalglomerulus: good neighbors and good fences. Development 135, 609-620.
Quiring, R., Walldorf, U., Kloter, U. and Gehring, W. J. (1994). Homology ofthe eyeless gene of Drosophila to the Small eye gene in mice and Aniridia inhumans. Science 265, 785-789.
Reddien, P. W. and Sánchez Alvarado, A. (2004). Fundamentals of planarianregeneration. Annu. Rev. Cell Dev. Biol. 20, 725-757.
Reddien, P. W., Bermange, A. L., Murfitt, K. J., Jennings, J. R. and SánchezAlvarado, A. (2005a). Identification of genes needed for regeneration, stem cellfunction, and tissue homeostasis by systematic gene perturbation in planaria.Dev. Cell 8, 635-649.
Reddien, P. W., Oviedo, N. J., Jennings, J. R., Jenkin, J. C. and SánchezAlvarado, A. (2005b). SMEDWI-2 is a PIWI-like protein that regulates planarianstem cells. Science 310, 1327-1330.
Rehorn, K. P., Thelen, H., Michelson, A. M. and Reuter, R. (1996). A molecularaspect of hematopoiesis and endoderm development common to vertebratesand Drosophila. Development 122, 4023-4031.
Reisinger, E. (1922). Uber die Terminalorgane and das Kanalsystem einigerbekannter Typholoplaniden. Zoologischer Anzeiger 56, 205-224.
Rink, J. C., Vu, H. T. and Sánchez Alvarado, A. (2011). The maintenance andregeneration of the planarian excretory system are regulated by EGFR signaling.Development 138, 3769-3780.
Ruppert, E. E. (1994). Evolutionary origin of the vertebrate nephron. Am. Zool.34, 542-553.
Ruppert, E. E. and Smith, P. R. (1988). The functional organization of filtrationnephridia. Biol. Rev. 63, 231-258.
Sajithlal, G., Zou, D., Silvius, D. and Xu, P. X. (2005). Eya 1 acts as a criticalregulator for specifying the metanephric mesenchyme. Dev. Biol. 284, 323-336.
Sánchez Alvarado, A., Newmark, P. A., Robb, S. M. and Juste, R. (2002). TheSchmidtea mediterranea database as a molecular resource for studyingplatyhelminthes, stem cells and regeneration. Development 129, 5659-5665.
Scimone, M. L., Meisel, J. and Reddien, P. W. (2010). The Mi-2-like Smed-CHD4gene is required for stem cell differentiation in the planarian Schmidteamediterranea. Development 137, 1231-1241.
Self, M., Lagutin, O. V., Bowling, B., Hendrix, J., Cai, Y., Dressler, G. R. andOliver, G. (2006). Six2 is required for suppression of nephrogenesis andprogenitor renewal in the developing kidney. EMBO J. 25, 5214-5228.
Suzuki, T., Delgado-Escueta, A. V., Aguan, K., Alonso, M. E., Shi, J., Hara, Y.,Nishida, M., Numata, T., Medina, M. T., Takeuchi, T. et al. (2004). Mutations in EFHC1 cause juvenile myoclonic epilepsy. Nat. Genet. 36, 842-849.
Tena, J. J., Neto, A., de la Calle-Mustienes, E., Bras-Pereira, C., Casares, F.and Gomez-Skarmeta, J. L. (2007). Odd-skipped genes encode repressors thatcontrol kidney development. Dev. Biol. 301, 518-531.
van der Flier, L. G. and Clevers, H. (2009). Stem cells, self-renewal, anddifferentiation in the intestinal epithelium. Annu. Rev. Physiol. 71, 241-260.
Wagner, D. E., Wang, I. E. and Reddien, P. W. (2011). Clonogenic neoblasts arepluripotent adult stem cells that underlie planarian regeneration. Science 332,811-816.
Wang, Q., Lan, Y., Cho, E. S., Maltby, K. M. and Jiang, R. (2005). Odd-skippedrelated 1 (Odd 1) is an essential regulator of heart and urogenital development.Dev. Biol. 288, 582-594.
Weavers, H., Prieto-Sanchez, S., Grawe, F., Garcia-Lopez, A., Artero, R.,Wilsch-Brauninger, M., Ruiz-Gomez, M., Skaer, H. and Denholm, B. (2009).The insect nephrocyte is a podocyte-like cell with a filtration slit diaphragm.Nature 457, 322-326.
Weintraub, H., Davis, R., Tapscott, S., Thayer, M., Krause, M., Benezra, R.,Blackwell, T. K., Turner, D., Rupp, R., Hollenberg, S. et al. (1991). The myoDgene family: nodal point during specification of the muscle cell lineage. Science251, 761-766.
Wenemoser, D. and Reddien, P. W. (2010). Planarian regeneration involvesdistinct stem cell responses to wounds and tissue absence. Dev. Biol. 344, 979-991.
Wilson, R. A. and Webster, L. A. (1974). Protonephridia. Biol. Rev. Camb. Philos.Soc. 49, 127-160.
Xu, P. X., Adams, J., Peters, H., Brown, M. C., Heaney, S. and Maas, R. (1999).Eya1-deficient mice lack ears and kidneys and show abnormal apoptosis oforgan primordia. Nat. Genet. 23, 113-117.
Xu, P. X., Zheng, W., Huang, L., Maire, P., Laclef, C. and Silvius, D. (2003). Six1is required for the early organogenesis of mammalian kidney. Development 130,3085-3094.
Yang, J., Liu, X., Yue, G., Adamian, M., Bulgakov, O. and Li, T. (2002).Rootletin, a novel coiled-coil protein, is a structural component of the ciliaryrootlet. J. Cell Biol. 159, 431-440.
Zhu, J., Hill, R. J., Heid, P. J., Fukuyama, M., Sugimoto, A., Priess, J. R. andRothman, J. H. (1997). end-1 encodes an apparent GATA factor that specifiesthe endoderm precursor in Caenorhabditis elegans embryos. Genes Dev. 11,2883-2896.
RESEARCH ARTICLE Development 138 (20)
DEVELO
PMENT


















