
A plastid DNA phylogeny of the genus Acacia Miller (Acacieae, Leguminoseae)
28
Bolanicaljournal ofh Linnean S&p (2000), 132: 195-222. With 4 figures doi:10.1006/boj1.1999.0301, available online at http://www.idealibrary.corn on IDEN1 @ * A plastid DNA phylogeny of the genus Acacia Miller (Acacieae, Leguminoseae) JUW ROBINSON* AND STEPHEN A. HARRIS Department o f Plant Sciences, University of Oxfod, South Parks Road, Oxford OX1 2RB Received F e b w v 1999; acceptedfor publication Sejhnber 1999 Past classifications of the tribe Acacieae Rchb. are outlined and the confusion concerning the relationships of the three subgenera of Acacia Mill. are highlighted. A plastid DNA analysis of Acacieae shows that the genus Acacia is not monophyletic. Furthermore subgenera Acacia Vassal and Anrleiferwn Vassal are sister taxa and neither appear closely related to subgenus fiyllodineae (DC.) Ser. Subgenera Acacia and Aculciferum form a clade that is basal to a well-supported clade consisting of tribe Ingeae Benth. taxa, Faidherbia albida @el.) A. Chev. and subgenus Phyllodincuc. The series of relationships suggested by the cpDNA data contradicts previous investigations of the tribe. Possible explanations of this conflict are explored, and the taxonomic implications of the plastid DNA data set are considered. 8 2000 The Linnean Society of London ADDITIONAL, KEY WORDS:--RFLp - Ingeae - Mmosoideae - gene tree. CONTENTS Introduction ........... Material and methods ....... Material .......... Analysis ofplastid DNA ..... Data analysis ......... Results ............ Discussion ........... Relationships within the Tribe Acacieae Relationships within the Ingeae ... Taxonomic implications of the cpDNA data Conclusions ........... Acknowledgements ........ References ........... ............ ............ ............ ............ ............ ............ ............ ............ ............ ............ ............ ............ ............ 195 198 I98 198 20 1 20 1 204 205 209 209 210 210 21 1 INTRODUCTION Acacia Mill. is a widespread genus of tropical-subtropical trees and shrubs ranging from CentralBouth America through Africa to south-east Asia and Australia (Ross, * Corresponding author. E-mail: [email protected] 0024-4074/00/030195 + 28 835.00/0 0 2000 The Linnean Society of London 195
-
Upload
julian-robinson -
Category
Documents
-
view
216 -
download
0
Transcript of A plastid DNA phylogeny of the genus Acacia Miller (Acacieae, Leguminoseae)

Bolanicaljournal o f h Linnean S&p (2000), 132: 195-222. With 4 figures
doi:10.1006/boj1.1999.0301, available online at http://www.idealibrary.corn on IDEN1 @ *
A plastid DNA phylogeny of the genus Acacia Miller (Acacieae, Leguminoseae)
J U W ROBINSON* AND STEPHEN A. HARRIS
Department of Plant Sciences, University of Oxfod, South Parks Road, Oxford OX1 2RB
Received F e b w v 1999; acceptedfor publication Sejhnber 1999
Past classifications of the tribe Acacieae Rchb. are outlined and the confusion concerning the relationships of the three subgenera of Acacia Mill. are highlighted. A plastid DNA analysis of Acacieae shows that the genus Acacia is not monophyletic. Furthermore subgenera Acacia Vassal and Anrleiferwn Vassal are sister taxa and neither appear closely related to subgenus fiyllodineae (DC.) Ser. Subgenera Acacia and Aculciferum form a clade that is basal to a well-supported clade consisting of tribe Ingeae Benth. taxa, Faidherbia albida @el.) A. Chev. and subgenus Phyllodincuc. The series of relationships suggested by the cpDNA data contradicts previous investigations of the tribe. Possible explanations of this conflict are explored, and the taxonomic implications of the plastid DNA data set are considered. 8 2000 The Linnean Society of London
ADDITIONAL, KEY WORDS:--RFLp - Ingeae - Mmosoideae - gene tree.
CONTENTS
Introduction . . . . . . . . . . . Material and methods . . . . . . .
Material . . . . . . . . . . Analysis ofplastid DNA . . . . . Data analysis . . . . . . . . .
Results . . . . . . . . . . . . Discussion . . . . . . . . . . .
Relationships within the Tribe Acacieae Relationships within the Ingeae . . .
Taxonomic implications of the cpDNA data Conclusions . . . . . . . . . . . Acknowledgements . . . . . . . . References . . . . . . . . . . .
. . . . . . . . . . . .
. . . . . . . . . . . .
. . . . . . . . . . . .
. . . . . . . . . . . .
. . . . . . . . . . . .
. . . . . . . . . . . .
. . . . . . . . . . . .
. . . . . . . . . . . .
. . . . . . . . . . . .
. . . . . . . . . . . .
. . . . . . . . . . . .
. . . . . . . . . . . .
. . . . . . . . . . . .
195 198 I98 198 20 1 20 1 204 205 209 209 210 210 21 1
INTRODUCTION
Acacia Mill. is a widespread genus of tropical-subtropical trees and shrubs ranging from CentralBouth America through Africa to south-east Asia and Australia (Ross,
* Corresponding author. E-mail: [email protected]
0024-4074/00/030195 + 28 835.00/0 0 2000 The Linnean Society of London 195

196 J. ROBINSON AND S. A. HARRIS
TABLE I . Outlines of the major classifications within the tribe Acacieae, and the distributions of the taxa recognized
Bentham ( I 875) Vassal (1972) Pedley ( I 986) Distribution
Acacia series Gummijmae Benth.
Ac& subgenus Acacia Acacia
series Vukan-s Benth. subgenus Aculkfmrm Smcgalia series Filicinac Benth. Vassal Raf. series Bottyccphah Benth. subgenus Phyllodineae Racospma series Phyllodineae Benth. (DC.) Ser. (DC.) Mart. series Achellae Benth.
Faidherbia Faidhnbia
America, Africa, Asia and Australia
America, Africa, Asia and Australia
Australia, New Guinea, Hawaii,
Mascarene Islands Africa
1981). Approximately 1200 species of Acacia have been described, the majority of which are endemic to Australia (Mash & Stirton, 1997).
The taxonomy of the genus has been in a state of flux since it was first described by Miller (1 754). Initially it was an ill-defined, heterogeneous assemblage and it was not until Bentham’s series of papers (1 841 , 1842a, by 1845, 1846, 1865, 1875) that the present generic limits were established. Bentham (1841) adopted the tribe Acacieae Rchb. as one of the three Tribes constituting his suborder Mimosoideae, and included the genus Acacia, with the genera Albizia Durazz., Calliandra Benth., Lysiloma Benth., Ggia Walp., Enterolobium Mart., Pithecellobium Mart., Serianthes Benth., Inga Mill. and Afomea A. St.-Hil. (Bentham 1842a, b). Bentham (1865) subsequently made substantial changes to the tribe by restricting it to Acacia, creating the tribe Ingeae Benth. to accommodate the other genera, and dividing Acacia into six series (Table 1; Bentham, 1875). These series were based primarily on vegetative characters (foliage and spinescence), and secondarily on inflorescence characters; fruit was largely ignored (Bentham, 1875). This classification formed the basis for subsequent revision of Acacia.
Vassal (1972) and Guinet & Vassal (1978) attempted to define precisely generic subdivisions and determine their degree of relationship to each other. Vassal (1 972) recognized three subgenera, broadly similar to Bentham’s (1875) series (Table l), and moved the African species Acacia albida Del. to the monotypic genus Faidherbia A. Chev. within the tribe Acacieae. Vassal (1 972) distinguished three subgenera, Acacia, Acuh$mm and Phyllodineae, each defined by a suite of characters. He named this subgenus Hetemphyllum Vassal, but the name Phyllodineae (DC.) Ser. has priority. Guinet & Vassal (1978) then assessed the degree of specialization of each of these characters and divided them into three states, unspecialized, specialized and highly specialized. For example, inflorescence type was divided into spicate (unspecialized), obloid (specialized), and globose (highly specialized). Guinet & Vassal (1978) con- cluded that if the correlation between subgenera and characters was correct then, subgenera Aculeiferum and Phyllodineae were more closely related to each other than either was to subgenus Acacia, despite subgenera Acacia and Aculeiferum sharing a common geographical distribution: the Neotropics, Asia and Africa. That species groups represented in the Americas have a preponderance of ‘unspecialized‘ character states, and character specialization occurred away from the Americas, led Guinet & Vassal (1978) to suggest an origin of the genus in West Gondwana.
Despite its shortcomings, primarily based upon the apparently arbitrary way in

PHYLOGENY OF AC4CL4 197
which character polarity was determined and the way that continuous variation was split into discrete classes, Guinet & Vassal (1 978) produced a workable phylogenetic hypothesis for the genus. This hypothesis formed the basis for Pedley’s (1986) phylogenetic treatment of the genus, which aimed to determine: (i) whether the genus Acacia should be treated as three separate taxa; and (ii) whether these taxa should be treated as genera, rather than subgenera or series.
Pedley (1986) concluded that the genus Acacia, as defined by Guinet & Vassal (1978), was too broadly conceived, and three genera should be recognized, Acacia Mill. ( =Acacia subgenus Acacia), Senegalia RaE ( =Acacia subgenus Acula.mm) and Racospenna (DC.) Mart. (=Acacia subgenus Phyllodineae; Table l), on the basis that proposals to split other large taxa had been made, e.g. Cassieae Bronn (Leguminoseae; Irwin & Barneby, 1981), Thtania R. Br. (Myrtaceae; Wilson & Waterhouse, 1982), Euca&tus L‘Her. (Myrtaceae; Johnson, 1976), and through the application of Williams’ index of diversity (Williams, 1964). Like Guinet & Vassal (1978), Pedley (1 986) concluded that subgenera Aculezzmm and Phyllodineae were closely related, both being distinguished from subgenus Acacia. Pedley (1986) suggested that direct derivation of Senegalia from Acacia sensu Pedley, or the reverse, was unlikely and that Acacia sensu Pedley and Senegalia/Racospenna were derived from different lines within the Ingeae. In addition, Pedley (1986) suggested that Acacia sensu Pedley was affiliated with Pithecellobium (Ingeae) and that Senegalia and Racospemza had affinities with Calliandra (Ingeae). This implies that the tribe Acacieae is polyphyletic, although no data were presented by Pedley (1986) to support Pithecellobiurn and Calliandra as relatives of Acacia sensu Pedley and Senegalia/Racospenna respectively.
Researchers in legume systematics greeted these proposals unenthusiastically (Pedley, 1987), although they stimulated new research into the genus and its relatives (Maslin, 1987). Chappill & Maslin (1995) in a cladistic analysis of the genera of the Mimosoidem concluded that (i) the genus Acacia was not monophyletic and (ii) subgenera Aculafmm and Phyllodineue form a monophyletic group basal to the tribe Ingeae, with subgenus Acacia and Faidherbia nested separately within the tribe Ingeae. Whilst supporting previous studies (Guinet 1969, Pedley 1986), Chappill & M a s h (1 995) considered it inadvisable to split the genus formally. This was the first study to analyse objectively morphological characters in the Acacieae. In addition, Chappill & Maslin (1995) conducted an infrageneric analysis of the genus Acacia the results of which broadly supported the findings of the generic analysis.
Grimes (1995) investigated the generic relationships of the tribe Ingeae, although not explicitly investigating the relationships of the Acacieae, exemplars of the three subgenera (sensu Vassal) of Acacia and Faidherbia were included. Grimes’ (1 995) results indicated that the Acacieae and Ingeae should be amalgamated, although the relationships of the Acacia subgenera to each other were unclear due to an unresolved polytomy. All three Acacia subgenera, and Faidherbia, were basal to a well-supported clade consisting of Pithcellobium and its allies.
These results are in direct contradiction to those of Chappill & Maslin (1995); this is also the case for the many of the Ingeae genera common to both analyses. Thus there is a case for determining the relationships of Acacia subgenera based on alternative data sources. To aid this endeavour Maslin & Stirton (1997) compiled a list of critical species ofdcacia, with which a comparative data set could be constructed. M a s h & Stirton (1997) suggested an informal classification of Acacia (based on Vassal, 1972 and Pedley, 1978). Within this classification they grouped together species according to their presumed taxonomic affinities. In each of the groups

198 J. ROBINSON AND S. A. HARRIS
several taxa were highlighted as being representative of the group, and Maslin & Stirton (1 997) suggested that research on the genus should include as many of these taxa as possible. For example, Acacia subgenus Acuh.$mm section Aculeiferum was divided into two groups; those with prickles in pairs (e.g. A . rnellijira (Vahl) Benth.), and those with prickles solitary or in threes (e.g. A. senegal (L.) Willd.).
The conservative nature of plastid DNA evolution, and the low levels of homoplasy seen in cpDNA restriction site studies (Givnish & Sytsma, 1997) make this approach useful for producing independent data for assessing phylogenetic relationships, for example Pennington (1 995), Wiegreffe, Sytsma & Guries (1 998), Mason-Gamer & Kellogg (1996), Jansen et al. (1992), Olmstead & Palmer (1992).
The aims of the present study are (i) to evaluate whether the tribe Acacieae is monophyletic; and (ii) to determine the relationships between the taxa in the tribe Acacieae.
MATERIAL AND METHODS
Material
The choice of Acacia taxa (Table 2) was based upon Maslin & Stirton’s (1997) list of ‘critical species’ for the preparation of comparative data sets in the genus. Ten of the 15 groups defined by Maslin & Stirton (1997) were represented in the sampling, and additional taxa were analysed to give broad sampling of subgenera Acacia and Acuhfmn. Within subgenus Phyllodineae, with approximately 900 taxa, and over 103 ‘critical taxa’, the taxa included in this study were chosen from a list of ‘suitable’ taxa supplied by Bruce Maslin (Western Australia Herbarium, pers. comm.) as representative of the subgenus. Additional taxa, broadly sampled from the Ingeae, and piptudenia viridjriora (Kunth) Benth. from the Mimoseae were also included.
Anabsis ofplastid DNA
Total DNA was extracted from either fresh or silica-gel dried (Chase & Hillis, 199 1) leaves according to a modified version of Doyle & Doyle (1 987), and purified on caesium chloride density gradients (Maniatis, Fritsch & Sambrook, 1982). DNA from dried material was extracted according to Harris & Robinson (1994). Ap- proximately 500ng of total DNA were digested with each of 15 6bp-cutting restriction endonucleases (Aha I, BamH I, Bcl I, Bgl 11, Ch I, EcoR I, EcoR V, HinD 111, I, Pst I, h 11, Stu I, Smu I, Sst I, Xho I; New England Biolabs Inc.) according to the manufacturer’s instructions. Digested DNA fragments were separated on 1 Yo agarose gels and transferred to nylon membranes using the method of Southern (1 975). Nylon membranes were sequentially probed with non-overlapping 32P- labelled V f a radiatu plastid DNA probes (Feinberg & Vogelstein, 1983; Dowling et al., 1996). Probes MB8 and MB9 were used together, whilst probes MB1, MB2, MB3, MB5, MB7, MB 1 1, MB12 were used individually. Together these probes covered more than 89% of the chloroplast genome. The resulting autoradiographs were then used to construct restriction maps of the taxa (Palmer, 1986), using the

PHYLOGENY OF ACACU 199
I 2 3 4 5 6 7
8 9
10
I I
12
13 14 15
16 17 18
19
20
21 22 23 24 25 26 27 28 29 30 31 32
33 34 35 36
37 38 39
40 41 -
TABLE 2. Taxa used in the plastid DNA analysis of the tribe Acacieae
Taxon Geographic Accession origin" identifier
Group No?
tribe Acacieae Acacia subgenus Acacia Vassal
Acacia abyssinica Hochst. ex Benth. Acacia a& Schinz Acacia dolichoccphala Harms Acacia dwpanolobium Harms ex Sjostedt Acacia erioloba E. Mey Acacia exuvialir Verd. Acacia herbcclada DC. subsp. chb& (0. B. Mill.)
Acacia kanvo Hayne Acacia hahi t z i i Engl. var. hahitzii Acacia niloticn (L.) Willd. ex Del. subsp. kraussiaa
Acacia nilolica (L.) Willd. ex Del. subsp. subalata (Vatke)
Acacia nilohia (L.) Willd. exDel. subsp. tommfosa (Benth.)
Acacia rhemanniana Schinz Acacia sieberana DC. Acacia sieberana DC. var. modii (Burtt Davy) Keay &
Acacia sgal Del. var.Jictula (SchweinE) Oliv. Acacia sgal Del. var. spa1 Acacia fortilk (Forssk.) Hayne subsp. hcteracantha
(Burch.) Brenan Acacia forfili (Forssk.) Hayne subsp. raddiana (Savi)
Brenan Acacia &rfilir (Fonsk.) Hayne subsp. spimcarpa (Hochst.
ex Rich) Brenan Acacia xanthophloea Benth. Acacia ammtucta DC. Acacia c a m (Mol.) Mol. Acacia chori@lplla Benth. Acacia c u q o Barneby & T. A. Zanoni Acacia darmon Ekman & Urb. Acqufamcsiana (L.) Willd. Acacia macracantha Humb. & Bonpl. ex Wdld. Acacia [mnatula (Schltr. & Cham.) Benth. Acacia pringlei Rose Acacia migii Lkon Acacia s c h a w (S. Watson) F. J. Herm.
subgenus Acultifcm Vassal section Acuhifmun Vassal Acacia burkei Benth. Acacia cafia phunb.) Wdld. Acacia galpinii Burtt Davy Ac& mel@ra (Vahl) Benth. subsp. debnm (Burch.)
Acacia n&scens Oliv. Acacia persiq7ora Pax Acacia po!wntha Wdld. subsp. campylacantha (Hochst.
Acacia smcgal (L.) Wdld. var. leiorhachir Brenan Acacia senegal (L.) Willd. var. senegal
Schreib.
(Benth.) Brenan
Brenan
Brenan
Brenan
Brenan
w A. Rich) Brenan
Zimbabwe Zimbabwe
Kenya Tanzania Zimbabwe
(Africa) (Africa)
Zimbabwe (Africa)
Zimbabwe
Kenya
Cape Verde
(Africa) Zimbabwe Zimbabwe
Malawi Kenya
Zimbabwe
Senegal
Zimbabwe
Malawi Mexico
(New World) Cuba Cuba Cuba
Guatemala Cuba
Honduras Mexico Cuba
Mexico
(Africa) (Mica) (Africa) (Africa)
(Africa) Kenya
Tanzania
Botswana Tanzania
39/90 RDB 1908" FAGG 634d
12/92' 23/88' HNBG' HNBG'
RDB 18 17A" HNBG'
RDB 1801Bd
81/90'
70/90'
HNBG' 7 1 / 9 0
1 /89'
77/92' 86/90'
RDB 2131"
I402/84'
RDB 1919"
87/90' HUGHES 1602"
455-65-4501' RICO I 122% RICO 1156y RICO 1O8Og
HUGHES 276" RICO I 124
67/87' HUGHES 1647"
RICO I 152R HUGHES 1632"
HNBG' HNBG HNBG' HNBG'
HNBG' FAGG 640"
11/92'
38/90 17/92'
I
I
I
I 1 I I 1 I I I I I 1 I
2 2 2 2
2 2 2
2 2
continued

200 J. ROBINSON AND S. A. HARRIS
TABLE 2. continued
Ref Taxon Geographic Accession Group origin'' identifier NO.^
42 43 44 45 46 47 48 49
50 51 52 53
54
55
56 57
58 59
60
61 62 63 64
65
66
67
68
section Monocanthea Vassal Acacia ataxacantlra DC. Acacia bmtircapa Harms Acacia charirssa Milne-Redh. Acacia montigma Brenan & Exell Acacia gaumm' S. F. Blake Acacia glomcmsa Benth. Acacia pgii A. Gray Acacia ripark Kunth secfion Filicinae (Benth.) Taub. Acacia anguihsima (MU.) Kuntze Acacia chamlensis L. Rico Acacia msn' Stand. Acacia kquilana S. Wats.
section Bohyccphakze (Benth.) Tauh. Acacia meamrii De Wild. section A l a k (Benth.) Pedley Acacia alab R. Br. section phyllodineae DC. Acacia paradoxa Schldl. Acacia pycnantha Benth. section Plurinms (Benth.) Maiden & Betche Acacia h a A. Gray Acacia melanoglon R. Br.
Faidlmbia albida (Del.) A. Chev.
subgenus phyllodineac (DC.) Seringe
Faidhtrbia
tribe Ingeae AlbizM
AIbirin h a q i Fournet Albizia schimpcrana Oliv. Albkia lomcntma (Micheli) Stand. Albizia ucsicolor Welw. ex Oliv.
&manta saman (Jacq.) Merr.
Enhlobium Gyclocarpum (Jacq.) Griseb.
I5thtcellobium dulce (Roxb.) Benth.
Samanea
Enhlobium
Rthtctllobium
tribe Mimoseae RptUdCtlia
Pipfudmia uidflora (Kunth) Benth.
Kenya Kenya (Africa) Kenya Mexico Mexico
(New World) Mexico
Guatemala Mexico
(New World) Mexico
Kenya
(Australia)
(Australia) (Australia)
(Hawaii) (Australia)
Namibia
(Africa) (Africa) Mexico (Africa)
Honduras
Honduras
Honduras
Mexico
FAGG 622d FAGG 624''
HNBG' FAGG 621'' R E 0 116Iy
RIG0 1005BR
RICO I 152R 161-80-01439K
37/88 RIWI 832BK RICO 8328 RICO 120IK
FAGG 638'
341 -83-04 103'
000-73-18422' 635-88-05356'
178-90-01 142' no. to follow'
84/90'
HNBG' HNBG'
HUGHES 959' HNBG'
69/87'
HUGHES.428'
HUGHES 426'
HUGHES 168Id
5 3 3 3 3 3 3 3
4 4 4 4
6
8
11 I0
13 13
~
'Reference number. See Appendiu, a table of the restriction site changes.
' Alice Holt Seed Bank, Farnham, Surrey, UK. Vouchers have not been assigned to these accessions. 'Herbarium material collected by C. E. Hughes (HUGHES #), C. Fagg (FAGG #) or R. Barnes (RDB #),
Material collected from living collections, Harare National Botanic Gardens, Harare, Zimbabwe. No accession
Country or Continent.
Dept. of Plant Sciences, University of Oxford. Vouchers at FHO.
numbers available. 'Kew Living Collection, Royal Botanic Gardens, Kew, Richmond, Surrey, UK. Field Material either collected or provided by Dr L. Rico-Arce, Royal Botanic Gardens, Kew. Vouchers at K.
hGroup Number from Mash and Stirton (1997). Taxa with numbers in italic are mentioned in Maslin and Stirton (1997), for others we have assigned them to a group.

PHYLOGENY OF AC4CIA 20 I
site occurrence analysis (SOA) of Bremer (1991). The fragments were sized using internal standards (HinD III/h) and the algorithms derived by Schaffer (1993). A complete data matrix is available from the senior author (JR), and a list of the restriction sites, the probe which revealed the change, and the taxa in which the change occurred are presented in the Appendix.
Data anahsis
Restriction sites were scored as either present (l), absent (0) or data missing (?) for each of the 68 taxa studied. Phylogenetic analysis was performed using PAUP* 4.0 b (Swofford, 1993) on an Apple Power Macintosh 730/200. Due to the size of the data matrix (68 taxa x 39lcharacters), exact search methods were impractical, therefore heuristic methods were used; 100 random addition replicates, using the tree bisection-reconnection swapping algorithm (Swofford, 1993). PAUP was also used to construct decay analyses and perform bootstrap and jackknife procedures; both the bootstrap and jackknife analyses used 1000 replicates of a ‘fast heuristic’ search in PAUP.
RESULTS
Restriction maps were constructed for all 68 taxa studied. At least 559 restriction sites were identified. Assuming a genome size of approximately 150 kb (Palmer, 1985), 3354 bp or 2.2% of the genome was sampled. Of the 391 polymorphic sites, 137 (35%) were autapomorphic, and 245 (65%) informative sites, shared between two or more taxa.
The heuristic analysis resulted in 720 minimal length trees of length 675 steps (Fig. lA, B), with a consistency index of 0.55 (including all characters), and a retention index of 0.84. Three major clades were identified from the consensus tree (Fig. 1A). Clude A. This clade consisted of all taxa from the tribe Ingeae, Acacia subgenus Phyllodineae, and Faidher6ia albda (Del.) A. Chev. The clade occurred in trees up to 20 steps longer than the minimal length tree and in 100% of the bootstrap and the jacknife analyses.
Chde B. This clade consisted of all Acacia subgenus Acacia taxa investigated. The clade occurred in trees that were up to nine steps longer than the minimal length tree and in 97% of the bootstrap and the jackknife analyses.
CZude C. This clade consisted of all Acacia subgenus AcuZefmm taxa investigated. This clade was not well-supported, being lost when trees one step longer than the minimal length tree were considered, and had zero bootstrap and jackknife support. Within this clade there were two well-supported clades. One corresponded to section Filicinae, and occurred in all trees 20 steps longer than the minimal length tree. The other clade, corresponded to sections Monocantha and Acuhimm, and occurred in all trees up to six steps longer than the minimal length tree plus, 53% and 51% of the bootstrap and jackknife analyses respectively.
Clades A and B are well supported, with high decay indices and bootstrap/

202 J. ROBINSON AND S. A. HARRIS
I
Foldherbfa albida Albizlo wrsicolw Albizia h a m Albizia schimperana Somonea soman Enterolobfum cyclocorpurn Albizlo tomentoso Acacia mel?piylon f J
Acacia koo Acacia prodoxa $1 Acacia ahta 8 Acacia pycnontho 10 Acocb mearnsii * Pithecelloblum duke Acacia nilotica kraurctrmrr I l Acacia nilotica tomentm Acacia nllotico subdata 1 Acacia leuderitzii leudcr/tzli Acacia twtllis heteracanthp
Acacia abyssinica Acaclo tortllfs spirfKarp0
' f 1
Acach orenaria 1 Acacia eriolo5 1
Acacia xanthophloea 1 Acacia sieberana 1
~cacf" korry
"1" A$; '$1 s-l - Acacia sieberana woodii 1
Acacia ripoffa 3 Acacia mumeri 3
I I A A c a c i o h o n f i g e n a eg il 3 3
Acacia breviscaw 3 b domerm J
I
atitha 5
- Acacieae
lngeae
subgenus Phyllodineae
-I - lngeae
su bgenus Acacia
subgenus Aculelf erum
Acacia angustlssimo 4 Acacia rosef 4 Acacia chamelensis 4 Acocla tequilano 4 Piptadenla virldlflora - Mimoseae
Figure 1. A, strict consensus tree of the 720 trees, length 675, generated by the heuristic search of PAUP. AgtadenM viridjtl~f~ was specified as the outgroup. The numbers above the branches are the bootstrap and jackknife values respectively. Below the branches are the decay indices. There are three clades labelled A, B, and C (see text for discussion of these clades). The numbers associated with the terminal Acacia spp. correspond to their grouping in M a s h & Stirton (1997); along the right of cladogrm are the taxonomic groups to which the taxa belong. B, one of the 720 most parsimonious trees. Numbers above the branches indicate how many character states change (gain or loss of a restriction site) on that particular branch. The lengths of the branch are proportional to the amount of change that occurs on them,

203
Figure 1. continued.

204 J. ROBINSON AND S. A. HARRIS
1-8 Faidherbia albida tribe lngeae subgenus Phyllodineae subgenus Acacia subgenus Aculeiferum tribe Mimoseae
Figure 2. Simplified strict consensus tree (from Fig. 1).
jackknife values. Clade C is less well supported, though within this clade two lineages are well supported. Within clade A the clade corresponding to subgenus Phyllodineae is well supported (Fig. 1). The support for clades B and C, an ‘Acacia ss.’ clade is good. It is well-supported (69 and 70%) in the bootstrap and jackknife analyses, but lost when trees four steps longer than the minimal length tree are considered. Decay indices greater than this produce a trichotomy of clades A, B and C.
DISCUSSION
Three clades were identified in the strict consensus tree; subgenera Acacia (clade B), Acuhijimm (clade C) and Phyllodineae (in clade A) are monophyletic (Fig. 2). The monophyly of each subgenus has not been disputed, since each subgenus is clearly defined by many morphological and chemical characters (Guinet & Vassal, 1978; Pedley, 1986; Maslin, 1988; Chappill & Maslin, 1995). However, the relationships between the subgenera indicates that the genus Acacia is not monophyletic (Fig. 2). Subgenera Acacia and Aculeiferum form a monophyletic group, whilst subgenus Phyllodineae is separated from this clade by members of the tribe Ingeae. Faidherbia albida appears basal to the Ingeae and subgenus Phyllodineae clade.
Statistics and indices for measuring tree ‘robustness’ are varied, but the most commonly used are consistency index (C.I., Kluge & Farris, 1969), decay indices (Bremer, 1988) and bootstrap/jackknife values (Felsenstein, 1985; Farris et al., 1996). Consistency index is viewed as an inverse measure of overall homoplasy for a particular tree (Givnish & Sytsma, 1997), and Sanderson & Donoghue (1 989) showed that the C.I. decreases with increasing numbers of taxa analysed. Givnish & Sytsma (1997) list 40 cpDNA restriction site studies, and within this list there are few studies utilizing as large a number of taxa as in this study, although comparing the C.I. of this study (C.I. =0.554) it appears that a C.I. of 0.554 appears to be standard for 68 taxa. In contrast, retention index (R.I.; Farris, 1989) indicates how well the characters fit the tree that describes them, and R.I. in this study (0.84) is high.
The non-monophyly of Acacia s.1. has been proposed in two recent treatments (Pedley, 1986; Chappill & Maslin, 1995). However, the conclusions of these in- vestigations differ substantially from the cpDNA results reported here. Pedley (1 986) concluded that subgenera Aculeiferum and Phyllodineae were closely related; both clearly distinguished from subgenus Acacia. Furthermore he indicated that subgenus Acacia and Phyllodineae and Aculeiferum, were derived from different lineages within the Ingeae. Chappill & Maslin (1995) presented similar conclusions concerning the non- monophyly of Acacia s.1. and the relationships of the subgenera. Subgenus Acacia was

PHYLOGENY OF A C X U 205
sister to Calliandra, whilst subgenera Acule fmm and Phyllodineae form a monophyletic group basal to the tribe Ingeae and subgenus Acacia (Fig. 3).
Relationships within the tribe Acacieae
Results of the present investigation of cpDNA restriction site variation in the subgenera of the genus Acacia, contradict the results of Chappill & Maslin (1995), and all previous investigations (e.g. Pedley, 1986; Guinet & Vassal, 1978; Guinet, 1990). The cpDNA results show that subgenera Acacia and Aculeijimm are sister taxa and that neither subgenus is closely related to the Ingeae, in contrast to previous suggestions (Pedley, 1986). In contrast, subgenus Phyllodineae, is not closely related to the other Acacia subgenera, rather it appears most closely related to genera within the tribe Ingeae (Fig. 2). A close relationship between subgenera Aculeiferum and Pbllodineae has been suggested in previous classifications and investigations of the genus (Pedley, 1986; Guinet, 1990; Brain, 1987, 1990; Conn et al., 1989). These studies utilized morphological and chemical characters; pollen morphology and free amino acid type have been the main characters used to distinguish subgenera Aculeiferum and Phyllodineae from subgenus Acacia (Pedley, 1986; Guinet, 1990; Chappill & Maslin, 1995; Table 3).
Pollen characters appear to provide the strongest link between subgenera Acule fmm and Phyllodineae (Guinet, 198 1); both subgenera lack columellae with either porate or extraporate apertures. Subgenus Acacia has columellae with colporate apertures. Subgenera Acacia and Aculeifemm however share a lack of exine ornamentation. In the Tribes Ingeae and Mimoseae all the pollen characters found in Acacieae are also present (Guinet, 1990). Free amino acids offer less satisfactory evidence of a close relationship between the subgenera Acule fmm and Phyllodineae, although they illustrate that subgenus Acacia is distinct from the other subgenera. Again all of the free amino acids are found in other taxa in the tribes Ingeae and Mimoseae (Southon, 1994).
A criticism of both these data sets is their polymorphic nature; several character states have arisen more than once (Chappill & Maslin, 1995). For example, the occurrence of albizzine in the seeds; albizzine is absent from subgenus Acacia, but is present in some, but not all taxa from subgenera Aculeijimm and Phyllodineae and tribe Ingeae. The spectrum of unrelated taxa that have albizzine present in their seeds effectively rules out a single origin of albizzine, and suggests the possibility of multiple losses of albizzine. This is true whichever phylogeny is ‘correct’. It is possible that some characters have been lost repeatedly rather than having multiple origins. Additionally, some taxa appear to be ‘united’ in their lack of certain characters, e.g. both subgenera Acule fmm and Phyllodineae lack columellae. The interpretation of both sets of data appears to depend crucially on the weight given to certain characters and how the polarity of those characters has been determined. This, when combined with an independent data set which strongly suggests a different set of relationships, would indicate that re-evaluation of these morphological and chemical characters is needed.
Recently Dayanandan, Bawa & Kesseli (1997) sequenced the rbcL gene of the cpDNA genome for several tropical woody legume taxa, including some Ingeae and one Acacia species. Unfortunately the strict consensus tree is only partially resolved. However, all the Ingeae studied (only Pithecellobium, Enterolobium, Albizia and Balizia

N 0
m
TA
EIL
E
3. D
istri
butio
n of
pol
len
char
acte
rs a
nd s
eed
free
am
ino
acid
cha
ract
ers t
hat
unite
sub
gene
ra A
cuh
im an
d I'
yWkm
. Po
llen
data
wer
e ta
ken
from
Tab
le 2
in M
aslin
(19
88),
and
the
seed
free
am
ino
acid
data
wer
e ta
ken
from
App
endi
x 1
in C
happ
ill &
Mas
lin (
1995
). Fo
r am
ino
acid
data p
olym
orph
ic in
dica
tes
that
the amino
acid
was
pre
sent
in s
ome
of t
he s
peci
es o
f the
sub
genu
s, bu
t no
t in
oth
ers,
e.g
. will
ardi
ne is
fou
nd in
se
ctio
ns M
man
thea
and
Fdi
cina
~ of s
ubge
nus A
cuh
im bu
t not
in s
ectio
n An
clcife
nrm
. The
Inge
ae d
escr
iptio
ns fo
r am
ino
acid
dis
tribu
tion
are
base
d on
ly o
n th
e ge
nera
CaW
ra, H
avan
iiu a
nd P
aras
mia
dus as C
happ
ill &
Mas
lin o
nly
incl
uded
dat
a fr
om th
ese
gene
ra i
n th
eir
infr
agen
eric
ana
lysi
s. A
dditi
onal
cha
ract
ers f
rom
Mas
lin (1
988)
Cha
ract
er
Subg
enus
Su
bgen
us
Subg
enus
Fa
idlu
rbia
Tribe
Inge
ae
Ac&
AG
Uhlfm
un
PI$J!0
&iZ
albi
da
Polle
n -a
pert
ure type
-col
umel
lae
-exh
e or
nam
enta
tion
Free a
min
o ac
ids i
n se
eds
-s-c
arbo
xeth
yl cy
stei
ne
-car
boxis
o pr
opyl
cyst
eine
-a
lbuz
ine
-wiU
ardi
ne
-alp
habe
tad&
opro
pion
ic
acid
dj
enko
lic a
cid
-N-a
cety
l dje
nkol
ic ac
id
-pip
ecol
ic ac
id
40H
-pip
ecol
ic a
cid
-5-O
H-p
ipec
olic
aci
d -2
,4-c
ii-4,
5-tr
ansd
iOH
pip
ecol
ic a
cid
Oth
er c
hara
cter
s Sp
ines
cent
stip
ules
Pri
des
Phyl
lode
s
Col
pora
te
Pres
ent
Smoo
th
Abs
ent
Abs
ent
Abs
ent
Abs
ent
Abs
ent
Pres
ent
Pres
ent
Res
ent
Pres
ent
Abs
ent
Abs
ent
Pres
ent
Pora
te
(infr
eque
ntly
ex
trap
orat
e)
Abs
ent
Smoo
th
Poly
mor
phic
Po
lym
orph
ic
Poly
mor
phic
Poly
mor
phic
Po
lym
orph
ic
Poly
mor
phic
Abs
ent
Abs
ent
Poly
mor
phic
Abs
ent
Poly
mor
phic
Abs
ent
Ext
rapo
rate
Po
rate
(i
eque
ntly
Po
==)
Abs
ent
Abs
ent
Ret
icul
ate
(rar
ely
Fain
tly a
erol
ate
aero
hte)
Poly
mor
phic
Po
lym
orph
ic
Poly
mor
phic
Po
lym
orph
ic
Poly
mor
phic
Po
lym
orph
ic
Abs
ent
Poly
mor
phic
Pr
esen
t Po
lym
orph
ic
Poly
mor
phic
Abs
ent
Abs
ent
Pres
ent
Abs
ent
Pres
ent
Abs
ent
Pres
ent
Pres
ent
Pres
ent
Pres
ent
Abs
ent
Abs
ent
Pres
ent
Abs
ent
Pres
ent i
n so
me
Abs
ent
Abs
ent
Abs
ent
Abs
ent
Pres
ent
Abs
ent
taxa
Col
pora
te i
n C
allin
ndrn
sm
. sh
: Po
rate
in
the
4
rem
aind
er o
f th
e In
geae
exc
ept e
xtra
pora
te i
n so
me s
peci
es of
Cojo
b4 M
anna
m~l
on, Ob
oling
a and
Pres
ent
in
CaUi
andr
a s
m.
sh:
Abs
ent
in
the
rem
aind
er o
f th
e In
geae
A
reol
ate
or s
omet
imes
smoo
th
*
E i2 z U
Poly
mor
phic
Po
lym
orph
ic
Poly
mor
phic
A
bsen
t
Pres
ent
Pres
ent
Pres
ent
Poly
mor
phic
Poly
mor
phic
Po
lym
orph
ic
Abs
ent
Res
ent i
n so
me taxa
(Alb
izia,
CaUi
andr
a, H
ad
ia,
Pihc
cllo
bium
). Abs
ent i
n othen
Pres
ent i
n so
me
Albi
ria. A
bsen
t in
othe
r taxa
Abs
ent

PHYLOGENY OF AGICL4 207
Barneby & J.W.Grimes) formed a clade, to which Acacia collinsii SafK, a member of subgenus Acacia, was basal. This is entirely consistent with the cpDNA restriction site data, and would support the relationships suggested by the cpDNA restriction site data.
This is not the place for a critique of Chappill & Maslin’s (1995) analysis, but there are three important points to consider. Firstly, Chappill & M a s h (1995) conducted two analyses, a generic analysis of the Mimosoideae and an infrageneric analysis of the tribe Acacieae, the results from both analyses were partially con- tradictory, including the positioning of the subgenera of Acacia. For example, in the generic analysis subgenus Aculeiimm formed a monophyletic group basal to the Ingeae. However, in the infrageneric analysis, it formed a paraphyletic group between the Ingeae and subgenus Phyllodineae. Both these results contradict Grimes (1995). Secondly, both analyses appear to contain a large amount of missing data, which can seriously hinder phylogeny reconstruction (Swofford et al., 1996). Chappill & Maslin (1995) alluded to this in their generic analysis, and in the data matrix for the infrageneric analysis (appendix 1 in Chappill & M a s h 1995) approximately 28% of the data was coded as missing. Finally, a critical feature of cladistics is that synapomorphic characters must be identified and that they alone provide the basis for clade identification (Avise, 1994). Despite this Chappell & Maslin (1 995) include characters which we believe fall into this category.
Bukhari, Koivu & Tigerstedt (1999) using cpDNA RFLP data, assessed the relationships of the three subgenera of Acacia and Faidherbia, but did not discuss the phylogeny of Acacia s.1. in relation to any other mimosoid taxa. The characters Bukhari et al. (1 999) use appear to be fragments in the fragment occurrence analysis method of Bremer (1991). This may lead to problems with scoring homologous bands in the analysis, in contrast to the site occurrence analysis (SOA) used here. Bremer (1 99 1) showed that restriction fragment methods are less accurate than restriction site methods for phylogenetic analyses. The analysis reported here used a greater number of restriction enzymes than Bukhari et al.3 (1999) analysis, resulting in a greater number of characters in the data matrix. Both analyses found each subgenus to be monophyletic, only differing in the placement of Faidherbia. Bukhari et al.’s (1999) analysis placed Faidherbia within subgenus Aculefmm, the present investigation placed it basal to a combined Ingeae and subgenus Phyllodineue. The reason for this difference cannot be determined as Bukhari et al.’s (1999) analysis lacks any Ingeae.
Grimes (in press), in a continuation of his work on the Ingeae and their inflorescence morphology (Grimes, 1992, 1995), used exemplars from the Acacieae and showed that they were divided into two groups. A clade of Faidherbia and subgenera Acacia and Acuhamm, formed a sister clade to the Fithecellobium alliance (sensu Barneby & Grimes, 1996), whilst subgenus Phyllodineae was found to be basal to all other ingroup taxa. Direct comparison with Grimes (in press) is hindered by his concern with relationships in the Ingeae taxa and the inclusion of Acacieae exemplars, whereas the study reported here was concerned with the Acacieae and the included Ingeae exemplars. However, Grimes’ (in press) conclusions that in terms of inflorescence structure and growth, subgenera Acacia and Aculkfmm are more similar to American Ingeae than to subgenus Phyllodineue are supported by the results reported here.
There are several possible causes of discordance between gene trees and species trees in a cpDNA phylogeny (Maddison, 1997; Doyle, 1992), of which two will be

208 J. ROBINSON AND S. A. HARRIS
tribe Mimoseae subgenus PhyIlodineae subgenus Aculeiferum Enterolobium Albizia Pithecelloblum subgenus Acacia Calliandra Faidherbia
Figure 3. Simplified version of Chappill & Maslin’s (1995, fig. 1, p. 82) analysis of the Mimosoideae, based on morphological characters. Only taxa relevant to this study have been retained, other taxa have been ‘pruned’.
subgenus Phyllodlneoe subgenus Aculelferum
Albizlo plthecellobfum subgenus Acocfo Faidherbto olbfdo - Acacia- type - Ingeae- type
Figure 4. A possible lineage sorting scenario, see text for details. The two cpDNA-types discussed in the text have been overlaid on Chappill & Maslin’s cladogram for the generic analysis of the Mimoseae (Chappill & Maslim 1995, fig. 1, p. 82; also in condensed form here as Fig. 3).
considered here (i) ‘capture’ of cpDNA types; and (ii) lineage sorting or deep coalescence.
Hybridization leads to ‘chloroplast capture’ events (Arnold, 1 997). However, there are no known hybrids between subgenera Acacia and Aculeaamm, although species from each subgenus are sympatric across Central America, Africa and Asia (Ross, 1981). Hybrids are known, but these are limited to species within subgenera (New, 1984), e.g. A. xlaeta R. Br. ex Benth., a hybrid between A. mellijk-a and A. Senegal (El Amin, 1976) in subgenus Acuhzmm; and in subgenus P&llodineae, A. mangium Wdd. x A. auricul$onnis A.Cunn ex Benth. (Mash & McDonald, 1996). Thus it is unlikely that the cpDNAs of subgenera Acacia and Acul&mm are similar through hybridization.
Similar problems are identified when lineage sorting is considered. In order to reconcile the positions of subgenera Acacia and Aculea$imm in the cpDNA phylogeny and in Chappill & Maslin’s (1995) phylogeny (see Fig. 3); the ‘Acacia’ cpDNA type would have to have been kept throughout the evolution of the Ingeae, and only survived in the subgenus Acacia lineage (Fig. 4). If it is assumed that there are two types of cpDNA, an Ingeae-type and an Acaciu-type, which occurred in the common ancestor of the Acacieae and Ingeae, the Ingeae type would be found in all taxa from clade A (Fig. l), whilst the Acacia-type occurs in all taxa from clades B and C. Early on in the evolutionary history of the taxa, when subgenera Phyllodineue and

PHYLOGENY OF AC4CU 209
AcuhYemm split from the rest of the taxa, both lineages retained the two cpDNA types. However, when subgenera Phyllodineae and Acuhiftemm diverged, the cpDNA types were ‘sorted’; subgenus Phyllodineae retained the Ingeae-type and subgenus Aculeiferum retained the Acacia-type. In the genera of the Ingeae, Acacia-type cpDNA is lost, and the Ingeae-type is retained. Only in subgenus Acacia is the Acacia-type cpDNA retained. However, the time over which the two cpDNA types have been retained, and the selective sorting in which only subgenera Acacia and Acuh?emm have retained the Acacia-type cpDNA, suggests that lineage sorting is an improbable factor in the discordance between the morphological and cpDNA data.
In order to reconcile the discordances between the cpDNA and the morphological data an additional independent data set should be examined, e.g. sequence analysis of the nuclear ribosomal DNA internal transcribed spacer (ITS) region (Baldwin et al., 1992).
Relationsh@s within the Ihgeae
No conclusions can be drawn regarding the relationships of members of the Ingeae to each other as not enough taxa were studied. Within the Ingeae the relationships of the few taxa that were studied are again in contrast to the relationships of these taxa in the cladogram of Chappill & Maslin (1995; Fig. 3). Comparison with Grimes (1 995) is difficult as the taxa that are in common to both investigations form an unresolved polytomy (see fig. l a in Grimes, 1995). The only conflict at present between the data sets is the placement of the Acacieae taxa. Grimes (1995) did not look at the African taxa of Albiziz; the cpDNA phylogeny suggests they are related to the Samanea Merr. alliance within the Ingeae.
TAXONOMIC IMPLICATIONS OF THE cpDNA DATA
The cpDNA data have implications for the classification of the Tribes Acacieae and Ingeae. The tribe Acacieae as circumscribed by Vassal (1 98 1) is not monophyletic, which supports the conclusion of Chappill & Masfin (1995) that the boundaries between the Ingeae and Acacieae must be reassessed. Furthermore, the cpDNA data indicates that the genus Acacia, as currently circumscribed, is polyphyletic.
The tribe Acacieae comprises the genus Acacia and the monotypic genus Faidhbia, whilst the genus Acacia is divided into three subgenera, Acacia, Acula$mm and Phyllodineae. The cpDNA data do not support this classification; subgenera Acacia and Acuh@mm are sister taxa, whilst subgenus Phyllodineae is sister to the tribe Ingeae.
The cpDNA data indicate that: (i) subgenus Phyllodineae and the genus Faidherbia could be transferred to the Ingeae; and (ii) tribe Acacieae should be reduced to comprise only the genus Acacia with two subgenera, Acacia and Acuhiemm.
Subgenus Phyllodineae, if transferred to the Ingeae, would need a new generic name. Pedley (1986), suggested Racospaa, but Maslin (1988) questioned the validity of this name, since Racospenna may not be linked to a validly published description. Faidherbiz, has been considered to be part of the tribe Acacieae, but is more closely related to the tribe Ingeae on the basis of the cpDNA data; this realignment of the genus has been suggested by Guinet (1990) on the basis of pollen data.

210 J. ROBINSON AND S. A. HARRIS
The tribe Acacieae may contain only the genus Acacia with two subgenera, Acacia and Acuhamm. The cpDNA data support a monophyletic subgenus Acacia, but the support for a monophyletic Acuhzmm is not as strong. Thus until there is a compelling need to split Acacia into two genera, it is better to maintain two subgenera, Acacia and Acuh$imm.
Chappill & Maslin (1995) suggested that the generic name Acacia should be restricted and be applied to what is at present subgenus Phyllodimue on pragmatic grounds and that this was the preferred solution of Bentham (1840). This would mean re-typifiing the name Acacai on a taxon in subgenus Phyllodimae. The cpDNA data suggest that this would be imprudent. The present work suggests that subgenera Acacia and Acuhzmm are sister taxa, corresponding to the tribe Acacieae. Subgenus Phyllodineue, on the other hand, appears to be within the Ingeae, rather than the Acacieae and should not be considered for the name Acacia.
CONCLUSIONS
If the cpDNA phylogeny presented here is an accurate representation of the species phylogeny then major reassessments of the morphological characters within the Tribes Acacieae and Ingeae, and associated nomenclatural changes, are needed. Chappill & Maslin (1 995) and Grimes (1 995), both using morphological characters, produced very different classifications. Guinet & Vassal (1978: 509) commented that “taxonomic limits are frequently obscured by continuous variation of characters . . ., levels of specialization in different characters may be very unequal in any taxon. The relationships then overlap to a greater or lesser degree and limits become difficult to establish”, and this is perhaps the greatest difficulty in classifjling the Ingeae and Acacieae.
Should further studies suggest that the cpDNA data are an accurate representation of the species tree, this has important implications for the way morphological characters are perceived in this group. If, however, the opposite is true and morphological studies are a more accurate representation of the species tree, then this is potentially an interesting example of hybridization and lineage sorting and the effects these two phenomena have on the evolution of a widespread tropical tree genus.
ACKNOWLEDGEMENTS
J.R. thanks St. Andrews University for awarding him a studentship and is indebted to R. Ingram for her supervision during this project. We thank James Grimes and an anonymous referees for useful comments on this paper.
Many thanks also go to C. Fagg (Dept. of Plant Sciences, University of Oxford) and L. Rico-Arce (RBG, Kew) for their help in acquiring material for the study. J. Doyle (Cornell University) kindly provided a complete set of the mung bean cpDNA library.

PHYLOGENY OF AC4CL4 21 1
REFERENCES
Arnold ML. 1997. Natural hybridisation and evolution. Oxford Oxford University Press. Avise JC. 1994. Molecular markers, natural history and evolution. London: Chapman and Hall. Baldwin BG. 1992. Phylogenetic utility of the internal transcribed spacers of nuclear ribosomal DNA
in plants: an example from the Compositae. Molecular Phylngmtics and Evolution 1: 3-16. Barneby RC, GrixnesJW. 1996. Silk Tree, Guanacaste, Monkeys Earring: a generic system for the
synandrous Mimosaceae of the Americas. Part I Abarema, Albkia, and allies. Memoirs ofthe New rOrk Botunic Ganien 74: 1-292.
Bentham G. 1840. Contributions towards a flora of South America - enumeration of plants collected by Mr Schomburgk in British Guiana. Journal ofBotany 2: 127-146.
Bentham G. 1841. Observations on the distinctive characters of the Papilionaceae and Caesalpinieae, Suborders of Leguminosae. Journal ofBotuny 3: 125-133.
Bentham G. 1842a. Notes on Mimoseae, with a synopsis of species. London Journal of Botany 1:
Bentham G. 1842b. Notes on Mimoseae, with a synopsis of species. London Journal of Botuny 1:
Bentham G. 1845. Notes on Mimoseae, with a synopsis of species. LondonJouml ofBotany 4: 577422. Bentham G. 1846. Notes on Mimoseae, with a synopsis of species. LondonJouml ofBotany 5: 75-108. Bentham G. 1865. Leguminosae. In: Bentham G, HookerJD. Genera Phturum. London. Bentham G. 1875. Revision of the Suborder Mimoseae. lkzn.swtionr ofthe Linnean Sock& ofLondon 30:
Brain P. 1987. Immunology and phylogeny: a preliminary study of Acacia. South A h a n Journal of
Brain P. 1990. Immunology and phylogeny 11: further studies on Acacia. Suz&Awnse &!d$zr
Bremer B. 1991. Restriction data from chloroplast DNA is there only one accurate way of scoring?
Bremer K. 1988. The limits of amino acid sequence data in angiosperm phylogenetic reconstruction.
Bukhari YM, Koivu K, Tigerstedt PMA. 1999. Phylogenetic analysis of Acacia (Mimosaceae) as
Chappill JA, Maslin BR. 1995. A phylogenetic assessment of tribe Acacieae. In: Crisp M, Doyle
Chase MW, Hills HH. 1991. Silica gel: an ideal material for field preservation of leaf samples for
Conn EE, Seigler DS, Maslin BR, Dunn J. 1989. Cyanogenesis in Acacia subgenus Aculcifmrm.
Dayanandan S, Bawa KS, Kesseli R. 1997. Conservation of microsatellites among tropical trees (Leguminosae). Amnican Journal ofBofany 84. 1658-1663.
Dowling TE, Moritz C, PalmerJD, Rieseberg LH. 1996. Nucleic acids I n analysis of fragments and restriction sites. In: Hillii DM, Moritz C, Mable BK, eds. Molccuhr SystmatiEs, 2nd. edition. Massachusetts, Sinauer Assoc. Inc. 249-320.
Doyle JJ. 1992. Gene trees and species trees: molecular systematics as one-character taxonomy. $stam& Bofany 17: 144-163.
Doyle JJ, Doyle JL. 1987. A rapid DNA isolation procedure for small amounts of fresh leaf tissue. phytochical Bulletin 19: 11-15.
El Amin HM. 1976. Acacia lacta R.Br. ex Benth., considered as a species of hybrid origin. Sudon Silva 21: 14-23.
F d s JS. 1989. The retention index and the rescaled consistency index. CMistics 5 417-419. Farris JS, Albert VA, KBLllersj6 M, Lipscomb M, Kluge AC. 1996. Parsimony Jackknifing
Feinberg AP, Vogelstein B. 1983. A technique for radiolabelling DNA restriction fragments to high
Felsenstein J. 1985. Confidence limits on phylogenies: an approach using the bootstrap. Evolution
Givnish 'lJ, Sytsrna KA. 1997. Homoplasy in molecular vs. morphological data. The likelihood of
3 18-392.
494-528.
335-664.
StimGC 83: 422-427.
Wekmkap 8 6 195-199.
P h t $ s t m d c s and Evolution 175 39-54.
Evolution 4 2 795-803.
revealed from chloroplast RFLP data. 77teorecical and AppliUi Genetics 98: 291-298.
J, eds. Advances in Legume SystematiGs 7: Phylngmy. Kew: Royal Botanic Gardens, 77-99.
DNA studies. Tarn 40: 215-220.
phyto~hi~@ 28: 8 17-820.
outperforms neighbor-joining. CMistics 12: 99-1 24.
specific activity. Anahtical Bwchishy 132: 6-13.
3 9 783-79 1.

212 J. ROBINSON AND S. A. HARRIS
correct phylogenetic inference. In: Givnish TJ, Sytsma KA, eds. Molecular Evolution and Adaptive Radiation. Cambridge: Cambridge University Press, 55- I0 1.
Grimes J. 1992. Metmerism, heterochrony and inflorescence morphology of the Pithecellobiurn-complex (Leguminosae: Mimosoideae: Ingeae). Britbnia 44: 14&159.
Grimes J. 1995. Generic relationships of Mimosoideae tribe Ingeae, with emphasis on the New World Pithcellobiurn complex. In: Crisp M, DoyleJ, eds. Advances in Legume Systematics 7: Phylogeny. Kew: Royal Botanic Gardens, 10 1- 12 1.
Grimes J. In Press. Inflorescence morphology, heterochrony, and phylogeny in the mimosoid tribes Ingeae and Acacieae. Botanical R e k 65.
Guinet P. 1969. Les MimosacCes, etude de palynologie fundamentale, correlations, evolution. Travauw de la section scient$que et technique, Imtilutfianpis de Pondti& 9 1-293.
Guinet P. 1981. Mimosoideae: the characters of their pollen grains. In: Polhill RM, Raven PH, eds. Advances in Legume Systrmatics Part 2. Kew: Royal Botanic Gardens, 835-855.
Guinet P. 1990. The genus Acacia (Leguminosae, Mimosoideae): its affinities as borne out by its pollen characters. Plant @ s h a h s and Evolution [Supplement 51: 8 1-90.
Guinet P, Vassal J. 1978. Hypotheses on the differentiation of the major groups in the genus Acacia (Leguminosae). Km Bulletin 3 2 509-527.
Harris SAY Robinson J. 1994. Preservation of Tropical Plant Material for Molecular Analyses. In: Adam, eds. Commation ofplant GenesII. Monographs. Syst. Bot. Missouri Bot. Gard. No. 48. St Louis, Missouri Botanical Garden, 83-92.
Irwin HS, Barneby RC. 1981. Cassieae. In: Polhill RM, Raven PH, eds. Advances in Legume Sysfematics. Kew: Royal Botanic Gardens, 97-106.
Jansen RK, Michaels m, Wallace RS, Kim yI, Keeley SC, Watson LE, PalmerJD. 1992. Chloroplast DNA variation in the Asteraceae: Phylogenetic and evolutionary implications. In: Soltis PS, Soltis DE, PalmerJD, eds. Molecular &hat i c s ofPlants. London: Chapman and Hall, 252-279.
Johnson US. 1976. Problems of species and genera in Euca&ptus (Myrtaceae). Plant systematics and Evolution 125: 155-167.
Kluge AG, Farris JS. 1969. Quantitative phyletics and the evolution of anurans. &shat i c /Zbology
Maddison WP. 1997. Gene trees in species trees. sysfnalic Biology 46: 523-536. Maniatis T, Fritsch EF, Sambrook J. 1982. Molecular cloning: A laboratoy manual. Cold Spring
Harbor Publications. Maslin BR. 1987. Report of discussions at I.G.S.M. Meeting No. 6 on the proposal to split Acacia.
Bulletin ofthe International Gmupfor the Study ofMimosoideae 15: 108-1 18. Maslin BR. 1988. Should Acacia be divided? Bulletin ofthe Inhational Gmupfor the Study ofMimosoideae
16: 54-76. Maslin BR, McDonald MW. 1996. A key b usejil Australian ac&for the seasonal& d y tropics. Canberra:
CSIRO. M a s h BR, Stirton CH. 1997. Generic and infra-generic classification in Acacia (Leguminosae:
Mimosoideae): a list of critical species on which to build a comparative data set. Bulletin of the International Gmupfbr the Stu& ofMimosoideae 20: 22-44.
Mason-Gamer RJ, Kellogg EA. 1996. Chloroplast DNA analysis of the monogenomic Triticeae: phylogenetic implications and genome specific markers. In: Jauhar PP, ed. Methods ofgenome ana&sis in plants. Boca Raton: CRC Press, 301-325.
18: 1-32.
Miller P. 1754. A Ganieners Lhtionay, abrd. 4th ed. London. New TR. 1984. A biology of A c a c k . Oxford Oxford University Press. Olmstead RG, Palmer JD. 1992. A chloroplast DNA phylogeny of the Solanaceae: subfamilial
PalmerJD. 1985. Evolution of chloroplast and mitochondrial DNA in plants and algae. In: MacIntyre
Palmer JD. 1986. Isolation and structural analysis of chloroplast DNA. Methodr in Enrymologv 118:
Pedley L. 1978. A revision of Acacia Mill. in Queensland. Ausfmbaihya 1: 75-234. Pedley L. 1986. Derivation and dispersal of Acacia (Leguminosae), with particular reference to
Australia, and the recognition of Senegalia and Racospenna. Botanical Journal of the Linnean So&g 92: 2 1 9-254.
relationships and character evolution. Annals ofthe Mksouri Botanic Ganien 79: 346360.
RJ, ed. Molecular Evolutionary Genetics. New York: Plenum Press, 13 1-240.
167-186.
Pedley L. 1987. In defence of Racospenna. Bulletin ofthe International Gmupfor the Study OfMimosoideae 15: 123-1 29.

PHYLOGENY OF AOlCLA 213
Pennington RT. 1995. Cladistic analysis of choloroplast DNA restriction site characeters in Andira
Ross JH. 1981. An analysis of the African Acacia species: their distribution, possible origins and
Sanderson w, Donoghue w. 1989. Patterns of variation in levels of homoplasy. Evolution 4 3
SchaHer HE. 1983. Determination of DNA fragment size from gel electrophoresis'mobility. In: Weir BS, ed. Stattitical anabsir ofDNA sequence data. New York Dekker, 1-14.
Southern EM. 1975. Detection of specific sequences and DNA fragments separated by gel electro- phoresis. Journal ofMolecular Biology 9 8 503-5 17.
Southon W, Bisby FA, Buckingham J, Harborne JB, eds. 1994. Phytochemical Dictionary ofthe Leguminoseae. London: Chapman and Hall.
Swofford DL. 1993. PAUP Phylogenetic Anabsis using Parsimony, Version 3.1. Illinois: Illinois Natural History Survey.
Swofford DL, Olsen GJ, Waddell PJ, Hillis DM. 1996. Phylogenetic inference. In: HilIis DM, Moritz C, Mable BK, eds. Molecular Systnnatics, 2nd. edition. Sunderland, MA: Sinauer, 407-5 14.
Vassal J. 1972. Apport des recherches ontogkniques et seminologiques i I'ktude rnorphologique, taxonomique et phylogknique du genre Acacia. Bulletin de la socGti d'histoire naturelle de %ulouse 108
Wiegreffe SJ, Sytsma KJ, Guries RP. 1998. The Ulmaceae, one family or two? Evidence from
Williams CB. 1964. Pattenzr in the balance ofnature. London: Academic Press. Wilson PG, Waterhouse JT. 1982. A review of the genus Tt tania R. Br.: a heterogeneous
Swofford DL. 1998. Vassal J. 1981. Acacieae Benth. In: Polhill RM, Raven PH eds. Advances in legume systematics. Part 1.
(Leguminosae: Dalbergieae). American Journal of Botany 82: 526-534.
relationships. Bothalia 13: 389-413.
1781-1795.
105-247.
chloroplast DNA restriction site mapping. Plant systematics and Evolution 210: 249-270.
assembledge of five genera. Australian Journal of Botany 30: 4 13-446.
Kew: Royal Botanic Gardens, 169-1 7 1.

214 J. ROBINSON AND S. A. HARRIS
APPENDIX
Table of the distribution of restriction site mutations among the Acacia species, Ingeae species and pitadmiu viridjfl,a used in this study, and the probe-enzyme combination which revealed them.
Enzyme probe Restriction site change' Tax mutation is present in"
APa 1 ApoI APaI
APa 1 APa I APa 1 APa 1
APa I Apa I APa I Apa I
Apa I
APa I APa I APa 1 Bum HI Bam HI Barn HI Barn HI Barn HI
Bam HI Barn HI
Barn HI Bam HI Bam HI Bam HI Bam HI Bam HI Bam HI Bam HI
Bam HI
Bam HI
Bam HI
Bam HI
Bam HI
Bam HI
Bam HI
Barn HI
MB 3 MB 3 MB3
MB 3 MB2 MB2 MB 5 MB5
MB 8+9,1 I MB 12 MB 12 MB 12
MB 12 MB 1 1 MB 12 MB3 MB7 MB 7 MB 11 MB 3
MB 3 MB 3
MB3 MB 2 MB 2 MB 5 MB 5 MB 5 MB5 MB5
MB 1
MBI
MB 1
MB 12
MB 8+9
MB 8 + 9
MB I I
MB 11
20 o 12.0+8.0
7.7 o 6.7+1.0 7.7+0.3 o 8
7.7 o 5.7f2.0 3.3+x" o LP
3.3+0.1 o 3.4 9.8+x o Lf
4.8+4.8 o 9.6
8.5 o 8.2+0.3 Lf o 9.9+x Lf @ 11.3+x Lf o 5.3fx
5.3 + 0.4 19.8 5.3
4.2+9.2 I +0.9
9 2.3+0.2 1.8+0.9
@ 5.7 o 10.4+9.4 o 4.5+0.8 e 13.4 w 1.9 o 7.7+1.3 @ 2.5 o 2.7
2.3 o 2.1 +0.2 3.4 o 3.1 +0.3
3.4 o 3.0+0.4
5 o 3.8+1.2 4.6 o 4.3+0.3
4.6+x o 7.8 4.6 o 6
3.3+ 1.2 o 4.5
3+0.8 o 3.8
20.5 o 4.5+ 16.0
4.4+0.2 o 4.6
4.4 o 4.0+0.4
4.4+ 1.5 o 5.9
3.1 e 2.7+0.4
0.6+0.7 o 1.3
3.5+0.3 o 3.8
6 o 3.0+3.0
4.0+2.0 o 6
63 46. 50. 52. 53 54; 55: 56, 57, 58, 59, 60, 61, 62, 63, 64, 65, 66,68 67 66 54, 56, 57, 58, 59, 61, 62, 63, 64, 67 49, 50, 52, 53, 57, 60 33, 35, 36, 37, 38, 39, 40, 41, 43, 45, 46, 47, 48, 50, 60, 65 54, 57, 58, 59, 60, 62, 63, 65, 66, 67, 68 65 33, 35, 36, 38, 39, 40, 41, 43, 45, 46, 48 1, 2,4,5,6, 7,8,9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 23, 27, 29, 30, 32, 54, 55, 56, 57, 58, 59, 61, 62, 63, 64, 66, 67, 68 23, 25, 26, 27, 28, 30, 31, 32 56 55,56 25, 28, 53 50, 51, 52, 53 49 25 54, 55, 56, 57, 58, 59, 60, 61, 62, 63, 64, 65, 66, 67 38, 60, 61, 62, 63, 64, 65, 66, 67, 68 33, 34, 36, 37, 38, 39, 40, 41, 44, 46, 49, 54, 56, 59 66 54, 55, 56, 57, 58, 59, 61, 62, 63, 64, 65, 66 62 60, 61, 64 36, 40, 41, 43, 44, 46, 47, 48, 49 67 33,39 33, 34, 36, 37, 38, 39, 40, 41, 43, 44, 45, 46, 47, 48, 49, 50, 51, 52, 53, 54, 55, 56, 57, 58, 59, 60, 61, 62, 63, 64, 65, 66, 67, 68 62,65
60, 63
67
43, 45, 46, 54, 55, 56, 57, 58, 59, 60, 61, 62, 63, 65, 66, 67
50, 52, 53, 54, 55, 56, 57, 58, 59, 60, 61, 62, 63, 64,65,66, 67
38, 49, 50, 51, 52, 53
67
35, 39
continued

PHYLOGENY OF AC4CU
APPENDIX-continued
215
Enzyme Probe Restriction site change' Tax mutation is present in"
Bam HI
Bam HI
Bam HI Bam HI Bam HI Bam HI Bam HI Bam HI Bam HI Bam HI Bam HI Bam HI Bam HI Bam HI Barn HI
Barn HI Barn HI Bam HI
Bam HI &1 I &1 I &I I
& I 1 &I I Bcl I Bcl I & I 1 & I 1 & I 1
&I I &I I & I 1 &I I &I1 &I1 &I1
&I I &I I
& I 1 &I1
&1 I & I 1 & I 1
MB I 1
MB 12
MB 12 MB 12 MB 12 MB 12 MB 3 MB 2 MB 5 MB 5 MB 7 MB 7 MB 7 MB 7
MB 11
MB I 1 MB 11 MB 11
MB I MB 3 MB 3
MB 3,2
M B 2 MB 2 MB 5 MB 5 M B 5 MB 5 MB 7
MB 7 MB 7 MB 7 MB 7 MB 7 MB 7 MB 7
MB 8 + 9 MB 11
MB 11 MB 11,12
MB 11,12 MB 11,12
MB1
0.9+0.7 0 1.6
3.2+0.6 0 3.8
5.0+0.6 0 5.6 0 5.6 0
5 0 1.8 @
2 0 4.6 a 3.3 ($
8.6 0 8.6 o
4.3+2.6 o 4.5 o 12 0
5.6 2.8 + 2.8 3.6+2.0 4.9 +O.l 1.6+0.2 1.7+0.3 3.5+l.l 30 + 0.24 6.2 + 2.3 4.5+4.1 6.9 3.9 + 0.6 10.0+2.2
2.6 0 2.5+0.1 6.0+2.5 0 8.5 0.8+0.8 o 1.6
4.4+0.7 0 5.1 4.7+0.3 0 5
4.8 0 2.9+ 1.9 23 0 10.0+13.0
12.0+7.0 o 19 12.0+1.0 o 13 2.3+2.2 0 4.5 1.8+0.2 a 2
1.5+1.8 o 3.3 1.9+2.7 @ 4.5
8 0 6.5+1.5
2.7 0 2.5+0.2 2.7+2.2 w 4.9 2.2+1.8 w 4
1.8 0 1.5+0.3 1.8 0 1.7+0.1
2.8 0 2.5f0.5 1.8+3.8 @ 5.6
12.5 o 10.0+2.5 4.7 o 4.3+0.4
4.7+3.5 o 8.4 12.5+1.6 o 14.1
12.5 0 10.3+2.2 12.5 0 6.9+6.1 3.5 0 3.3f0.2
54, 55, 56, 57, 58, 59, 60, 61, 62, 63, 64, 65, 66, 67
33, 35, 36, 37, 38, 39, 40, 41, 46, 49, 61, 62, 64, 65, 67 33, 34, 35, 36, 37, 38, 39, 40, 41, 44, 46, 49 61, 62, 63, 64, 65, 66, 67 46, 48, 68 14 50, 51, 52, 53 58 57 54 54 43, 45, 48 43, 45, 48 48 53, 54, 55, 56, 57, 58, 59, 60, 61, 62, 63, 64, 65, 66, 67 54, 55, 56, 57, 58, 59 54, 55, 57 38, 46, 49, 54, 55, 56, 57, 58, 59, 60, 61, 62, 63, 64, 65, 66, 67 56 28, 50, 52, 53 38, 64, 65 46, 54, 55, 56, 57, 58, 59, 60, 61, 62, 63, 64, 65,66,67 5, 43, 45, 47 36 8, 22, 23, 24, 26, 27, 28, 29, 30, 31, 32, 60 62,65 65 65 1 , 2 , 3 , 4 , 5 , 6 , 7 , 8 , 9 , 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 50, 51, 52, 53, 68 60 40, 41 65 68 33, 34, 35, 36, 37, 39, 40, 41, 44, 57, 61 10. 11, 12, 55, 62, 63 33; 34, 35, 36, 37, 38, 39, 40, 41, 43, 44, 45, 47, 48 60 1 , 2 , 3 , 4 , 5 , 6 , 7 , 8 , 9 , 1 0 , 1 1 , 1 2 , 1 3 , 1 4 , 1 5 , 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 68 65 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32 63 50, 51, 52, 53 61, 62, 63, 64, 65, 66, 67
continued

216 J. ROBINSON AND S . A . HARRIS
APPENDIX-continued
Enzyme Probe Restriction site change' Tax mutation is present in'
Bc1 I &I I Bcl I Bcl I Bcl I Bci1 Bc1 I Bcl I Bcl I Bc11 Bc11 Bc1 I Bcl1 Bgl I1
Bgl I1 Bgl I1 Bgl I1 Bgl I1 Bgl I1 Bgl I1 Bgl 11
Bgl I1
Bgl n Bgl n Bgl I1
Bgl II
Bgl I1 Bgl I1
Bgl I1
II
Bgl I1 Bgl I1 Bgl I1 Bgl I1 Bgl I1 Bgl I1 Bgl I1 Bgl II cla I cla I
Cla I
cla I cla I cla I
cla I
cla I
MB 1 MB 1 MB 1 MB 7 MB 7 MB 7 MB 11 MB 1 MB 1 MB I MB 1 MB 3 MB 5 MB 3
MB 3 M B 2 MB 2 MB 2 MB 5 MB 1 MB I MB 1 MB I M B 1
MB 8 + 9 MB 5
MB 5 MB 5 MB 7
MB 7
MB7 MB 7 MB 7 M B 7 MB7 MB 7
MB 8 + 9 MB 7,8+9
MB 3 MB 3
MB 3
MB 3 M B 3 MB 2
M B 2
M B 5
3.5+ 1.3 Q 4.8 4.4 Q 4.0+0.4 4.0 Q 3.6f0.4 2.2 e l , l + l . l 2.6 e 1.5+1.1
4.4 Q 2.4+2.0
4.2 Q 3.9+0.3 3.5 e 2.6+0.9
2.2+1.1 e 3.3
4.0+1.0 e 5
1.0+0.9 e 1.9 4.3+5.7 e 10 5.0+0.7 e 5.7 3.7+0.6 e 4.3
3.4+5.1 ($ 9.4 5.3+x e9 Lf
Lf e 7.2+x Lf e 9 2 + x
3.5 e 1.9+1.8 5.1+1.1 e 6.2
1.9+0.6 e 2.5 5.2+ 1.5 Q 6.7
2.8+0.4 e 3.2 6.0+2.9 Q 8.9
1.5 Q 1+0.5
6.2 Q 2.6+3.6
3.6 e 3.2+0.4 2.6 Q 2.4+0.2
6.1 +2.5 e 8.6
4.4+1.7 e 6.1
8.6 e 4.9+3.7 4.9 e9 3.4+1.5 2.3 e 2.0+0.3
2.3+1.7 @ 5 1.5+0.3 e 1.8
4.4 Q 3.9+0.5 4.5 e 3.9+0.6
1.6 Q 0.9+0.7 4.2+0.5 Q 4.7
4.0+2.6 Q 6.6
4 e 3.5+0.5
4 Q 1.8+2.2 5.3 e 4.3+1.3
1.7+0.1 e 1.8
1.5 Q 0.6+0.9
8.5 Q 5.5+3
68 55, 56, 57, 58, 59, 62, 63, 64, 65, 66 67 55, 57 54 54 55 54 58 54 54, 55, 57, 58, 59 59 28 54, 55, 56, 57, 58, 59, 60, 61, 62, 63, 64, 65, 66,67 14, 15, 23, 27, 31, 32, 63, 67 66, 67, 68 67, 68 66 7, 18, 20 60, 61, 62, 63, 64, 65, 66, 67 67 7, 18, 20 63 63 n z 43, 45, 54, 55, 56, 57, 58, 59, 61, 62, 63, 65, 66,67 57 54, 55, 56, 57, 58, 59, 61, 62, 63, 65, 66, 67 43, 45, 54, 55, 56, 57, 58, 59, 60, 61, 63, 64, 65, 66, 67 43, 45, 51, 52, 54, 55, 56, 57, 58, 59, 60, 61, 62, 63, 64, 65, 66, 67 54, 55, 56, 57, 58, 59 57 15, 33, 34, 35, 36, 37, 38, 39, 40, 41, 44, 46 54,57 54,57 43, 45, 46, 49, 68 3, 43, 45 31 50, 51, 52, 53 27, 29, 32, 33, 34, 35, 36, 37, 38, 39, 40, 41, 44,45 43, 45, 46, 47, 48, 49, 51, 52, 53, 54, 55, 56, 57, 58, 59, 67 66 3, 22, 27, 29, 30, 32, 38, 46, 49, 53 33, 34, 35, 36, 37, 38, 39, 40, 41, 42, 43, 44, 45, 46, 47, 48, 49, 50, 51, 52, 53, 54, 55, 56, 57, 58, 59, 60, 61, 62, 63, 64, 65, 66, 67 33, 34, 35, 36, 37, 38, 39, 40, 41, 42, 43, 44, 45, 46, 47, 48, 49, 56 28, 33, 34, 35, 36, 37, 38, 39, 40, 41, 42, 43, 44, 45, 47, 48, 49, 50, 51, 52, 53, 54, 55, 56, 57, 58, 60, 63, 64, 65, 66, 68
continued

PHYLOGENY OF ACACIA 217
APPENDM-conhued
Enzyme Probe Restriction site change' Tax mutation is present in"
cla I cla I cla I cla I Cla I cla I
cla I Cla I
cla I cla I Cla I cla I cla I cla I cla I cla 1 cla I Cla I Cla I Cla I Cla I Cla I cla I cla I Cla I
cla I Cla I
cla I cla I cla I
cla I cla I Cla I cla I cla I cla I Cla I Cla I Cla I cla 1 Eco RI
Eco RI Eco RI Eco RI Eco RI Eco RI Eco RI
MB 5 MB 5 MB 5 MB 1 M B I MB I
MB 1 MB 1
MB I MB7 MB7 MB 7 MB7 MB 7
MB 8+9 MB 8 f 9 MB 8+9 MB 8 f 9 MB 8 f 9 MB 8+9 MB 11 MB 11 MB 11 MB I 1 MB 1 1
MB 11 MB 3
MB 3,2 MB 3 MB 7
MB 7 MB7 MB 7
MB 8+9 MB 11 MB 11 MB I MB 1 MB 1 MB 3 MB 3
MB 3 MB 5 MB 5 M B 7 MB 7 MB 7
8.5 e 3.7-1-4.8 2.9 o 1.4f1.5 2.9 2.4-1-0.5 3.3 w 2.9+0.2
2.3 e 2.0f0.3
4.8 e 4.7+0.1 7.7 e 5.5+2.2
2.3fO.1 @ 2.4
8.2 o 3.9f4.2
3.2 o 2.2+ 1.0 1.2 0.7f0.5 5.1 e 1.3f3.8 11 e 5.0f7.0
5 e 3.7f1.3
3.6 e 2.9+0.7
3.6 @ 3.1 f0.5
3.2 @ 2.9f0.3
3.2f0.9 e 4.1
11.0+5.0 e 16
5.0f2.3 @ 7.3
3.6+0.4 e 4
3.2+0.7 e 3.9
1.3+0.1 e 1.4 5.0+0.4 e 5.4 3.5+5.0 e 8.5
3.5f2.5 Q. 6 2.5+0.3 2.8
0.8+ 1.6 o 2.4 6 e 5.4f0.6
11.5 w 10.0+1.5
10 6.5f3.5 3.5+1.5 e 5 5.6+4.7 e 10.3 2.5+0.8 e 3.3 5.1 +3.8 ($ 8.9 3.5+0.4 e 3.9
2.3 o 1.9f0.4 1.7 e 1.5f0.2 5.5 m 3.8+ 1.7 3.5 e 2.3+ 1.2 3.8 e 3.2+0.6
2.2 e 1.1+1.1 Lf o 4.8+x Lf o 1.9+x 1.5 e 1.0f0.5
4.3 e 2.2f2.1 2.5+1.1 e 3.6
26, 31, 62 50, 51, 52, 53, 55 63 50, 51, 52, 53, 68 10, 1 1 , 12 26, 33, 40, 42, 43, 45, 47, 48, 51, 52, 54, 56, 57, 58, 59, 62, 63, 64, 65, 66 63 26, 31, 33, 34, 35, 36, 37, 38, 39, 40, 41, 42, 43, 44, 45, 46, 47, 48, 49, 50, 51, 52, 53, 54, 55, 56, 57, 58, 59, 60, 62, 63, 64, 65, 66, 67, 68 5 64, 65 50, 51, 52, 53, 55 68 33, 34, 35, 36, 37, 38, 39, 40, 41, 44, 50, 68 64 19, 20, 23, 27, 32 39 55, 61, 62, 64, 66, 67 50, 5 I , 52, 60, 68 6, 23 63,65 53, 56, 58, 59, 60 24, 25, 26, 31, 55, 63 60 54, 55, 56, 57, 58, 59, 60, 61, 62, 63, 65, 66, 67 33, 34, 35, 36, 38, 39, 40, 41, 42, 43, 44, 45, 46, 47, 48, 49, 54, 55, 56, 57, 58, 59, 60, 61, 62, 63, 65, 66, 67, 68 50, 51, 52, 53 43, 45, 46, 48, 49, 51, 52, 53, 54, 55, 56, 57, 58, 59 55- 42, 43, 45, 47, 51, 52, 54, 55, 56, 57, 58, 59 38, 42, 43, 45, 46, 47, 48, 49, 51, 52, 53, 54, 56, 57, 58 54, 56, 57, 58 56, 58 56 43, 45, 48, 59 42 54,57 5.5 _.
54, 55, 56, 57, 58, 59 26, 31 46 54, 55, 56, 57, 58, 59, 60, 61, 62, 63, 64, 65, 66, 67, 68 67,68 67 62 68 64 3, 5, 22, 23, 24, 25, 26, 27, 28, 30, 31, 32, 33, 34, 35, 36, 37, 38, 39, 40, 41,. 42, 43, 44, 45, 46, 47, 48, 49, 50, 51, 52, 53, 54, 55, 56, 57, 58, 59, 60, 61, 62, 63, 64, 65, 68
continud

218 J. ROBINSON AND S. A. HARRIS
APPENDIX-continued
Enzyme Probe Restriction site change' Tax mutation is present in"
E c O RI Eco RI Eco RI Eco RI Eco RI
Eco RI E c O RI E c O RI E c O RI E c O RI Eco RI E c O RI E C O RI
E c O RI
EcO RI
E C O RI Eco RI Eco RI Ed0 RI E c O RI E c O RI Eco RI E c O RI h R I &a RI E c O RI E c O RI Eco RI E C O RI E c O RI Eco RV
Eco RV Eco RV Eco RV Eco RV Eco RV Eco RV Eco RV Eco RV EEo RV Eco RV Eco RV Eco RV Eco RV
Eco RV Eco RV Eco RV Eco RV Eco RV
MB 7 MB 7 MB 7 MB 7 MB7
MB7 MB 7
MB 7,8+9 MB 8+9 MB 8 + 9 MB 8+9
MB I MBI
MBI
M B I
MB 1 ME2 MB 2 MB5 MB5 MB 5 MB 7 MB 7 MB 7 MB 7
MB 8+9 MB 8+9 MB 8 + 9
MB 1 MBI MB 3
MB3 MB 3 MB3 MB 3 MB 3 MB5 MB5 MB 5 MB7 MB 7 MB7
MB 8+9 MB 11
MB 11 MB 11 MB 11 MB 11 MB 12
2.1 o 1.9+0.2 2.1+0.6 o 2.7 1.7+0.8 o 2.5
3.6+5.7 o 9.6 1.7 o 0.9+0.8
3.6 o 1.9+1.7 5.7 o 5.0+0.7
5.4+5 o 10.4 3.8+0.3 o 4.1
1+1.9 o 2.9
3.4+0.4 o 3.8
3.8 o 2.9+0.9
3.4 o 3.2+0.2
1.5 o 1.0+0.5
3.8 o 2.5+ 1.3
3.8+1.1 o 4.9 1.9+4.0 o 5.9
4 o 1.1+2.9 4 o 3.7+0.3 4 o 2.5+1.5 4 o 2.3+1.7 3 o 2.8+0.2 3 rn 2.6+0.4 3 o 2.3+0.7
11.6 o 2.7+8.9 5.2 o 4.6+0.6 5.2 o 4.3+0.9 1.9 a 1.8+0.1 3.8 o 3.5+0.3
Lf o 1.3+Lf
Lf o 8.5+x Lf o 10.8+x Lf o 8.5+x Lf o 13+9.1 Lf cs 6.1 +I4
3.1 o 2.3+0.8
0.3+3.8 o 4.1
3.1 +0.4 o 3.5 3.5+1.1 o 4.6
Lf o 2.4+Lf 19.5 o 10.0+9.5 9.7 o 6.2+3.5
6 o 4.0+2.0
6 o 3.3+3.7 6 o 4.5+1.5
3.9 o 3.4+0.5
5.6+1.2 o 6.8
6.1+3.9 o 10
6.5+1.2 o 7.9
65, 67 60 22, 24, 30, 49, 53, 55, 61, 62, 63, 64, 65 60 26, 31, 33, 34, 35, 36, 37, 38, 39, 40, 41, 42, 43, 44, 45, 46, 47, 48, 49, 54, 55, 56, 57, 58, 59, 60, 61, 62, 63, 64, 65, 66, 68 22, 30 68 1, 3, 4 61, 63, 65, 67, 68 40, 41, 50 22, 23, 24, 25, 26, 27, 30, 31, 32, 49, 52, 53 65 23, 30, 32, 33, 34, 35, 36, 37, 39, 42, 43, 44, 45, 48, 5 1, 52, 56, 64, 68 33, 34, 35, 36, 37, 38, 39, 40, 41, 42, 43, 44, 45, 46, 48, 49, 50, 51, 52, 53, 54, 55, 56, 57, 58, 59, 60, 61, 62, 63, 64, 65, 66, 67, 68 23, 24, 25, 26, 27, 28, 29, 31, 32, 50, 51, 52, 53,66 62,67 57, 60 61, 62, 64, 65, 66 55, 58, 59 57 54 28, 46, 54, 55, 56, 57, 58, 59 23, 24, 38 50, 51, 52 55 57 58, 59 54, 55, 56, 57, 58, 59 55 56, 58 24, 25, 28, 31, 33, 34, 35, 36, 37, 38, 39, 40, 41, 44, 49 27,32 64,67 67 60 54, 55, 56, 57, 58, 59, 62, 65 54, 55, 56, 57, 58, 59, 61, 63, 65, 66, 67 54, 56, 57, 60, 62, 65, 67, 68 67 50, 53 62, 65, 66 62, 65 33, 34, 35, 36, 37, 38, 39, 40, 41, 44, 64, 68 1, 2, 4, 5, 6, 7, 8, 9, 10, I I , 12, 13, 14, 15, 16, 17, 18, 19, 20, 21 61,64 63 21, 32 68 60
continued

PHYLOGENY OF AC4CL4
APPENDIX-canEinucd
219
Enzyme probe Restriction site change’ Tax mutation is present in”
Eca RV Eca RV Eca RV Eco RV Eca RV Eca RV Eca RV Eca RV Eco RV Eca RV &a RV Eco RV Eco RV Hin DIII
Hin DIII Hin DIII
Hin DIII Hin DIII Hin DIII Hin DIII Hin DIII Hin DIII Hin DIII Hin DIII Hin DIII Hin DIII
Hin DIII Hin DIII Hin DIII Hin DIII Hin DIII Hin DIII Hin DIII Hin DIII Hin DIII Hin DIII Hin DIII
Hin DIII Hin DIII Hin DIII Hin DIII Hin DIII Hin DIII Nii I Nii I Nii I Nsi I Nii I Nii I Nii I Nii I
Nii I
MB 12 MB 12 MB 12 MB 12 MB 12 MB 1 M B l MB3 MB 3 MB 3 M B I M B 1 MB2 MB 3
MB3 MB3
MB 3 MB5 MB 5 MB 5
MB 5,7 MB 5,7 MB 7 MB 11 MB 11 MB 12
MB 12 MB 12 MB 12 MB 12 MB 12 MB 12 MB 5,7
MB 8+9 MB 1 1 MB 1 I M B I
M B l MB 7
MB 8+9 MB 8+9 MB 8+9 MB 12 MB3 MB 3 MB 3 MB 3 MB3 MB5 MB 5 MB 5
MB 5
6 0 6.5 o
10.4 o 4.5 o
10.4 o 2.4 o 2.5 @ 13 o L f o L f o
0.3+2.4 o 1.1+2.4 @
10.8 o 11.0+5.0 o
3.5 + 2.5 4.3+ 1.2 4.5+5.9 3.1 + 1.4 9+1.4 2.0 f2.4 2.3+0.3 4.0+ 7.0 13.0+x 8.1 + x 2.7 3.5 6.2+4.6 16
55, 61, 65 63 33, 34, 35, 36, 37, 38, 39, 40, 41, 44, 46, 53 39 23, 60, 61, 62, 63, 64, 65, 66, 67, 68 50, 51, 52, 53 60 55 26, 46, 50, 52, 53 23 55 58 46. 49 54; 55, 56, 57, 58, 59, 60, 61, 62, 63, 64, 65, 66, 67
15.0+11.0 o 26.0 5.0+0.5 o 5.5
5.5+0.2 o 5.7 2.0+0.4 o 2.4 2.0+ 1.2 o 3.2 1.7+1.2 o 2.9
13.0+7.5 o 20.5
8.0+3.1 o 11.1
12.6 o 9.4f3.2
7.5 o 2.9+2.6
3.1 o 1.5+1.6 8 o 5.3f2.7
8 0 8.0+2.0 o
8 o 8 o 8 -
13.0+x o 13 o
14.8 o 2.9+0.8 o
3.1 o 5.0+7.0 o
4.5 + 3.5 10 6.3+ 1.7 7.5 +0.5 7.0+ 1.0 Lf 6.0+7.0 5.8+9.0 3.7 2.5 +0.6 12
1.7+0.5 4.3 4.7 11
11.2 13 6
4.3 4.3+2.1
5.7 4.3+0.7 4.4+ 1.5 5.5 +0.8 5.2+4.4
o 2.2 o 1.0+3.3 a 2.3f2.4 o 9.0+2.0 o 5.2+6.0 o 10.0+3.0 o 3.6+2.4 o 1.3+3.4 o 6.4 o 2.4+3.3 0 5 ($ 5.9 o 6.3 o 9.6
2.1 +0.3 o 2.4
60, 67 23, 30, 33, 34, 35, 36, 37, 38, 39, 40, 41, 42, 43, 44, 45, 47, 48, 49, 68 46, 50, 51, 52, 53 62,65 56, 67 3, 7, 22, 23, 30 50, 52, 53 66 50, 52, 53 63 33, 34, 35, 36, 37, 39, 40, 41, 44, 46, 49 1, 2,4,5,6, 7,8,9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 48 39 62 19, 20, 33, 34, 35, 36, 37, 39, 40, 41, 44, 60 62, 63, 67 46, 49, 68 66 43,45 43, 45, 48, 49 42, 43, 45, 46, 49 54, 55, 57 24, 25, 26, 28, 31, 38, 46, 49, 53, 54, 55, 56, 57, 58, 59 56 38 46,49 46 43, 45, 48, 49 53 50, 51, 52 60,67 9, 10, 11, 12 1 1 33, 34, 35, 36, 37, 39, 40, 41, 44, 50, 66 27, 50, 60, 61, 62, 63, 64, 65, 66, 67, 68 33, 34, 35, 36, 37, 39, 40, 41, 44, 50, 60, 61, 62, 63, 64, 65, 66, 67, 68 62,65
continued

220 J. ROBINSON AND S. A. HARRIS
APPENDIX-contininucd
Enzyme Probe Restriction site change' Tax mutation is present in"
Nii I Nsi I
Nki I Nii I Nii I Nii I Nsi I Nsi I
Nsi I Nji I
Nsi I Pst I
Pst I
Pst I
Pst I Pst I Pst I Pst I Pst I
Pst I Pst I Pst I Pst I Psl I Pst I Anc I1
Anc I1 Anc I1 Anc I1 AncII Anc I1 Anc I1 Ancn Anc I1
Anc I1 Ancn Anc I1 Anr I1 Anr I1 Anr I1 Anr I1 Sma I sma I
sma I sma I sma I
MB 5 M B 7
MB 7 MB 7
MB 8 + 9 MB 8 + 9 MB 8 + 9
MB 8+9,1 I
MB 11,12 MB 12
MB 12 MB 3
MB 3
MB 5
MB 7 MB 8+9,11 MB 8+9,11
MB 1 1 MB 12
MB 12 MB 3 M B 5
MB 8 + 9 MB 11 MB 12 MB 5
MB 5 MB 5 MB 7 MB 7
MB 7,8+9 MB 7,8+9 MB 8+9 MB 12
MB 12 MB 12 MB 5
MB 8+9,11 MB 11,12
MB 12 MB 8 + 9
M B 3 MB 2
M B 3 M B 5 MB 5
2.1 e 1.9+0.2 5.0+0.7 e 5.7
3.2+1.0 e 4.2 3.2 e 2.5+0.7 3.6 e 1.8+1.8
5.6+0.3 e 5.9 5.6+5.9 e 11.5
11 e 4+7
4.7+ 1.8 e 6.5 4.7+5.9 e 10.6
4.7+0.4 e 5.1 Lf e 12.5+Lf
2 e 1.0+1.0
Lf o 6.7+x
Lf e 15+4.5
8.7 e 6.2+2.5 5.8 ($ 4.3t-1.5 Lf cs 6.4+x
6.1+8.7 o 15
Lf e 9.4+x 14 e 9.7+3.6
7.2 e 6.3+0.9 5.8 e 5.3+0.5
Lf o 13+Lf
6.7+1.3 e 8
Lf o 8.0+x
Lf e 10.3+Lf 10.3 e 7.7+2.6 4.8 o 4.5+0.3
9 e 7.4+1.6 6.3 e 2.7+4.5
12.9 e 8.2+4.7 Lf O lO.O+Lf
4.8+0.4 O 5.2
Lf e 7.9+Lf Lf e 7.4+Lf Lf w 5.8+x
18.4 o 14.6+3.8 Lf O 5+x
4.5 0 2.75+2.75 17.5 e 12.2+5.3
1.3 O 1.0+0.3
12+11 e 23
2.4+1.6 w 4 2.6+0.4 e 3 2.6+1.6 e 4.2
67 27, 33, 34, 35, 36, 37, 39, 40, 41, 43, 44, 45, 50, 60, 61, 62, 63, 64, 65, 66, 67, 68 10, 1 I , 12,67 60, 61 63 33, 34, 35, 36, 37, 39, 40, 41, 44, 50, 63, 67, 68 62,64 33, 34, 35, 36, 37, 39, 40, 41, 44, 50, 60, 61, 62, 63, 64, 65, 66, 67, 68 33, 34, 35, 36, 37, 39, 40, 41, 44, 50 33, 34, 35, 36, 37, 39, 40, 41, 44, 50, 60, 61, 62, 63, 64, 65, 66, 67, 68 5 54, 55, 56, 57, 58, 59, 60, 61, 62, 63, 64, 65, 66,67 33, 34, 35, 36, 37, 38, 40, 41, 42, 43, 44, 45, 46, 47, 48, 49, 50, 51, 52, 53, 54, 55, 56, 57, 58, 59, 60, 61, 62, 63, 64, 65, 66, 67 50, 52, 53, 54, 55, 56, 57, 58, 59, 60, 61, 62, 63, 64, 65, 66, 67 29 36, 41, 60, 61, 62, 64, 66, 67, 68 50, 51, 52, 53, 55 43, 45, 46, 48, 49, 68 50, 51, 52, 53, 54, 56, 58, 59, 60, 61, 62, 63, 64, 65, 66, 67 68 54, 55, 56, 57, 58, 59 55,57 54, 55, 57 54,57 55.57 46; 49, 54, 55, 56, 57, 58, 59, 60, 62, 63, 64, 65, 66, 67 50, 52, 53 50, 52, 53 68 25, 26, 31, 50, 53, 60 61, 62, 64, 66 26, 31, 50, 51, 52, 53 50, 51, 52, 53 43, 45, 47, 48, 54, 55, 56, 57, 58, 59, 60, 61, 63, 64, 65, 66, 67 50, 53 51, 52, 62 22,30 42 55 30 50, 51, 52, 53 22, 30 54, 55, 56, 57, 58, 59, 60, 61, 62, 63, 64, 65, 66, 67 16, 17, 21 56, 62, 65 67

P m O G F A Y OF ACACL4 22 I
APPENDX-continwd
Enzyme Probe Restriction site change' Tax mutation is present in"
S m I sst I sst I ssl I sst I ssl I ssl I ssl I
ssl I
ssl I ssl I ssl I
ssl I ssl I ssl I ssl I ssi I slu I
slu I slu I slu I slu I slu I slu I slu I slu I slu I slu I stu I slu I sac I Xho I
xho I xho I x h o I xho I xho I Xho I Xho I
xho I XhOI xho I xho I Xho I Xho I XhOI Xho I
MB I1 MB 5 MB 5 MB 7
MB 8+9 MB 8+9 MB 11 MB 11
MB 11
MB 11 MB 11
MB 11,12
MB 12 MB 5 MB 11 MB 11 MB 11 MB 3
MB 3 MB 3 MB2 MB7 MB 7 MB 7 MB7 MB 12 M B I MB 1 MB 1 MB 7 MB 7 MB 3
MB5 MB 7 MB 7 MB 12 MB 12 M B I MB 1
MBl MB2 MB 2 MB 5
MB 12 MB 12 MB 12 MB 1
18 o 3.0+ 15.0
12 o 3.8+8.2 16 o 7.0+9.0
4.6 o 2.8+ 1.8 Lf o 1.5+x
12+x o Lf .
20.0+2.0 o 22 5.3+ 1.3 o 6.6
6.6+5.3 o 11.9
5.3 o 3.9+ 1.7 1.3 o 0.8+0.5
9 o 7.5+1.5
Lf o 4.0+Lf 10 o 8.0+2.0
5.3 o 4.3+ 1.0 6.6 o 4.7+1.9
1.0+1.0 H 2.0
7.8+7.8 o 15.6
7.8+2.1 16+11.5 6.4 + 6.4 9.3+2.3 11.6+8
9.3 + 7.0 10 Lf
6.8 + 3.7 5.5 2.3 10
3.5 19+11.5
o 9.9 o 27.5 o 12.8 o 11.6 o 19.6 o 16.3 o 5.4+4.6 o 13.5+Lf o 10.5 o 3.2+2.3 o 2.0+0.3 o 6.5+3.5 o 3.0+0.5 o 30.5
10.7+1.5 o 12.2
3.4+0.4 o 3.8 22+18 o 32
7.4 o 1.6+5.8
18.1 H 7.9+ 10.2 8.8 o 7.6+0.8 8.8 o 6.5+2.3
8.8 o 3.3+0.7 o
13.7+3.3 o 10.3 o
0.7+0.3 o 16.5 o
16 o 15+3 o
4.4+ 4.4 4.1 17 8.5+ 1.8 1 10.1 + 6.4 2.8+ 13.7 18
46 4 62 60 63 54, 56, 57, 58, 59, 60 61, 62, 64, 65, 66, 68 33,'34, 35, 36, 37, 38, 39, 40, 41, 44, 46, 49, 55, 57, 58, 59, 60, 61, 62, 64, 65, 66, 67, 68 33, 34, 35, 36, 37, 38, 39, 40, 41, 42, 43, 44, 45, 46, 47, 48, 49 50, 53 50,53 33, 34, 35, 36, 37, 39, 40, 41, 43, 44, 45, 47, 48, 52, 54, 55, 56, 57, 58, 59, 60, 61, 62, 63, 64, 65, 66, 67, 68 60 54,57 54, 55, 56, 57, 58, 59 55, 56, 57 56 42, 54, 55, 56, 57, 58, 59, 60, 61, 62, 63, 64, 65, 66, 67 16, 17, 21 67 50, 52, 53 6 I , 62, 63, 64, 65, 66 61 26, 31, 60 64 60 50, 52, 53 54, 55, 56, 57,58, 59, 60, 61,62, 63, 64, 65, 66 63 54, 55, 56, 57, 58, 59 55 50, 51, 52, 53, 54, 55, 56, 57, 58, 59, 60, 61, 62, 63, 64, 65, 66, 67 67 2 50, 52, 53 62 60 50, 51, 52, 53, 64 1,2, 3 ,4 ,5 ,6 , 7,8,9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 68 6 1, 62, 65, 66 47, 58, 59 55 54,57 43,45 58, 59 55 55
continued
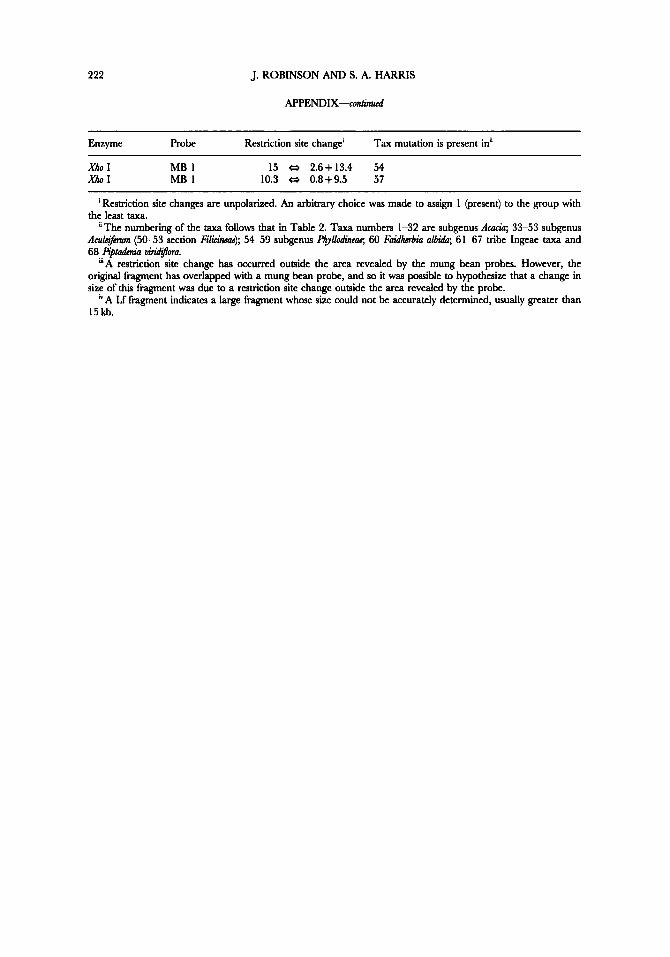
222 J. ROBINSON AND S. A. HARRIS
Enzyme Probe Restriction site change’ Tax mutation is present in”
Xho I MB 1 15 +s 2.6+13.4 54 xho I MB 1 10.3 M 0.8+9.5 57
i Restriction site changes are unpolarized. An arbitrary choice was made to assign 1 (present) to the group with the least taxa.
“The numbering of the taxa follows that in Table 2. Taxa numbers 1-32 are subgenus Acacia, 33-53 subgenus AculcJfmrm (50-53 section Filicineae); 54-59 subgenus phyllodincac; 60 Faidhcrbia albida; 6 1 4 7 tribe Ingeae taxa and 68 Rptadenia viridflora.
UiA restriction site change has occurred outside the area revealed by the mung bean probes. However, the original fragment has overlapped with a m u g bean probe, and so it was possible to hypothesize that a change in size of this fragment was due to a restriction site change outside the area revealed by the probe.
‘A Lf fragment indicates a large fragment whose size could not be accurately determined, usually greater than 15 kb.


















