
A NEW SPECIES OF THE SCINAX CATHARINAE SPECIES GROUP ...
12
A NEW SPECIES OF THE SCINAX CATHARINAE SPECIES GROUP (ANURA: HYLIDAE) FROM MINAS GERAIS, SOUTHEASTERN BRAZIL ANA CAROLINA CALIJORNE LOURENC ¸O 1,4 ,LUCIANA BARRETO NASCIMENTO 2 , AND MARIA RITA SILVE ´ RIO PIRES 3 1 Universidade Federal do Rio de Janeiro, Departamento de Vertebrados, Museu Nacional, Quinta da Boa Vista, Sa ˜o Cristo ´va ˜o, 20940-040, Rio de Janeiro, RJ, Brazil 2 Museu de Cie ˆncias Naturais, Departamento de Cie ˆncias Biolo ´gicas, and Programa de Po ´s-graduac ¸a ˜o em Zoologia de Vertebrados, PUC Minas, Av. Dom Jose ´ Gaspar 290, CEP 30535-610, Belo Horizonte, Minas Gerais, Brazil 3 Laborato ´rio de Zoologia dos Vertebrados, Instituto de Cie ˆncias Exatas e Biolo ´gicas, Universidade Federal de Ouro Preto, campus Morro do Cruzeiro, 35400-000, Ouro Preto, Minas Gerais, Brazil ABSTRACT: A new species of tree frog from the Scinax catharinae species group is described, discovered in the Municipality of Ouro Preto, southeastern Brazil. The new species is related to Scinax flavoguttatus, but differs from this species mainly by the light green color on its inguinal regions and hidden portions of flanks and thighs, by its subovoid snout, by its developed nuptial pad, and by its smaller males and females. Additionally, we describe the tadpole which is characterized by: dorsal position of nostril; dorsolateral eyes; gold spots between nostrils and eyes, short spiracle; spiracle opening on the midline of dorsal and ventral surface of body; absence of tail flagellum; oral disc with a short dorsal gap, surrounded by a uniseriate marginal papillae; M- shaped upper jaw sheath; tooth row formula 2(2)/3. Key words: Hylinae; New species; Scinax; Scinax catharinae species group; Tadpoles SCINAX is the largest genus within Hylinae, with 97 recognized species distributed from Mexico to Argentina and Uruguay (Frost et al., 2009; Moravec et al., 2009). The species of this genus occur in almost all major tropical and subtropical areas and are separated into two clades (Faivovich et al., 2005). The Scinax catharinae clade includes two species groups: Scinax catharinae and Scinax perpusillus (Faivovich et al., 2005). Presumed synapo- morphies of S. perpusillus species group are the oviposition in bromeliads and the extreme reduction of webbing between toes II and III (Peixoto, 1987; Faivovich, 2002). Currently, species from the Scinax catharinae group share the following morphological character- istics: absence of anterior process of the suprascapula, internal vocal sacs, distal divi- sion of middle branch of the muscle extensor digitorum comunis longus, and insertion of the medial side of this branch on the tendon of the muscle extensor brevis medium digiti IV. To date, 27 species are assigned to the S. catharinae group (Table 1). Despite the importance of larval characters for phyloge- netic and taxonomic studies, tadpoles of only 16 species are formally described for the S. catharinae group (see Conte et al., 2007). Herein, we describe a new species of the S. catharinae species group from southeastern Brazil, including a description of the tadpole. MATERIALS AND METHODS The specimens of the series type were collected at Estac ¸a ˜o Ecolo ´ gica do Tripuı´, Municipality of Ouro Preto, State of Minas Gerais, southeastern Brazil. These materials are housed at the Museu Nacional, Rio de Janeiro, Brazil (MNRJ) and the Museu de Cie ˆncias Naturais da Pontifı´cia Universidade Cato ´ lica de Minas Gerais, Belo Horizonte, Brazil (MCNAM). Specimens for comparisons are deposited in the Colec ¸a ˜o Adolfo Lutz (AL-MN) and herpe- tological collection of the Museu Nacional (MNRJ), Rio de Janeiro, Brazil and the Museu de Zoologia (MZUSP), Universidade de Sa ˜o Paulo, Sa ˜o Paulo, Brazil (Appendix 1). For comparisons of tadpoles, data were derived from published descriptions of 16 species of the Scinax catharinae species group (Andrade and Cardoso, 1987, 1991; Bokermann, 1964, 1967; Bokermann and Sazima, 1973; Carvalho- e-Silva, 1986; Carvalho-e-Silva and Carvalho-e- Silva, 1994, 1998; Carvalho-e-Silva et al., 1995; Conte et al., 2007; de Sa ´ et al., 1997; Haddad and Pombal, 1987; Pombal and Gordo, 1991). 4 CORRESPONDENCE: e-mail, [email protected] Herpetologica, 65(4), 2009, 468–479 E 2009 by The Herpetologists’ League, Inc. 468
Transcript of A NEW SPECIES OF THE SCINAX CATHARINAE SPECIES GROUP ...

A NEW SPECIES OF THE SCINAX CATHARINAE SPECIES GROUP(ANURA: HYLIDAE) FROM MINAS GERAIS, SOUTHEASTERN BRAZIL
ANA CAROLINA CALIJORNE LOURENCO1,4, LUCIANA BARRETO NASCIMENTO
2, AND
MARIA RITA SILVERIO PIRES3
1Universidade Federal do Rio de Janeiro, Departamento de Vertebrados, Museu Nacional, Quinta da Boa Vista,Sao Cristovao, 20940-040, Rio de Janeiro, RJ, Brazil
2Museu de Ciencias Naturais, Departamento de Ciencias Biologicas, and Programa de Pos-graduacao em Zoologia deVertebrados, PUC Minas, Av. Dom Jose Gaspar 290, CEP 30535-610, Belo Horizonte, Minas Gerais, Brazil
3Laboratorio de Zoologia dos Vertebrados, Instituto de Ciencias Exatas e Biologicas, Universidade Federal de OuroPreto, campus Morro do Cruzeiro, 35400-000, Ouro Preto, Minas Gerais, Brazil
ABSTRACT: A new species of tree frog from the Scinax catharinae species group is described, discovered inthe Municipality of Ouro Preto, southeastern Brazil. The new species is related to Scinax flavoguttatus, butdiffers from this species mainly by the light green color on its inguinal regions and hidden portions of flanksand thighs, by its subovoid snout, by its developed nuptial pad, and by its smaller males and females.Additionally, we describe the tadpole which is characterized by: dorsal position of nostril; dorsolateral eyes;gold spots between nostrils and eyes, short spiracle; spiracle opening on the midline of dorsal and ventralsurface of body; absence of tail flagellum; oral disc with a short dorsal gap, surrounded by a uniseriatemarginal papillae; M- shaped upper jaw sheath; tooth row formula 2(2)/3.
Key words: Hylinae; New species; Scinax; Scinax catharinae species group; Tadpoles
SCINAX is the largest genus within Hylinae,with 97 recognized species distributed fromMexico to Argentina and Uruguay (Frost etal., 2009; Moravec et al., 2009). The species ofthis genus occur in almost all major tropicaland subtropical areas and are separated intotwo clades (Faivovich et al., 2005). The Scinaxcatharinae clade includes two species groups:Scinax catharinae and Scinax perpusillus(Faivovich et al., 2005). Presumed synapo-morphies of S. perpusillus species group arethe oviposition in bromeliads and the extremereduction of webbing between toes II and III(Peixoto, 1987; Faivovich, 2002). Currently,species from the Scinax catharinae groupshare the following morphological character-istics: absence of anterior process of thesuprascapula, internal vocal sacs, distal divi-sion of middle branch of the muscle extensordigitorum comunis longus, and insertion ofthe medial side of this branch on the tendonof the muscle extensor brevis medium digitiIV. To date, 27 species are assigned to the S.catharinae group (Table 1). Despite theimportance of larval characters for phyloge-netic and taxonomic studies, tadpoles of only16 species are formally described for the S.catharinae group (see Conte et al., 2007).
Herein, we describe a new species of the S.catharinae species group from southeasternBrazil, including a description of the tadpole.
MATERIALS AND METHODS
The specimens of the series type werecollected at Estacao Ecologica do Tripuı,Municipality of Ouro Preto, State of MinasGerais, southeastern Brazil. These materialsare housed at the Museu Nacional, Rio deJaneiro, Brazil (MNRJ) and the Museu deCiencias Naturais da Pontifıcia UniversidadeCatolica de Minas Gerais, Belo Horizonte,Brazil (MCNAM).
Specimens for comparisons are deposited inthe Colecao Adolfo Lutz (AL-MN) and herpe-tological collection of the Museu Nacional(MNRJ), Rio de Janeiro, Brazil and the Museude Zoologia (MZUSP), Universidade de SaoPaulo, Sao Paulo, Brazil (Appendix 1). Forcomparisons of tadpoles, data were derivedfrom published descriptions of 16 species ofthe Scinax catharinae species group (Andradeand Cardoso, 1987, 1991; Bokermann, 1964,1967; Bokermann and Sazima, 1973; Carvalho-e-Silva, 1986; Carvalho-e-Silva and Carvalho-e-Silva, 1994, 1998; Carvalho-e-Silva et al., 1995;Conte et al., 2007; de Sa et al., 1997; Haddadand Pombal, 1987; Pombal and Gordo, 1991).4 CORRESPONDENCE: e-mail, [email protected]
Herpetologica, 65(4), 2009, 468–479
E 2009 by The Herpetologists’ League, Inc.
468

Sex was determined by dissection or by thereproductive condition, such as extendedvocal sacs in males and presence of oocytesin females. We follow Fabrezi and Alberch(1996), considering that the first digit is lost inanurans, thus digits are numbered II–V in thepresent study. The measurements of adultsfollow Duellman (1970) and the abbreviationsare SVL (snout vent length), HW (headwidth), HL (head length), END (eye nostrildistance), IND (internostril distance), ED(eye diameter), ESD (eye snout distance),IOD (interorbital distance), TD (tympanumdiameter), AL (arm length), TL (thigh length),SL (shank length), FL (foot length), FW (discwidth of finger III), TW (disc width of toe IV).We performed an ANOVA to verify significantvariations between the snout vent length(SVL) of the new species and each other ofthe Scinax catharinae group for which we hada sufficient sample size. In this test, twogroups of variables were formed: SVL andspecies. The first was considered dependentand the second was considered indepentent.The difference between these measurements
was considered significant when P values were,0.05. Seventeen tadpoles were collected atthe same location as the adults. Nine tadpoleswere fixed and preserved in 5% formalin.Additional tadpoles were reared to froglets inorder to allow specific identification. Eighttadpoles and eight juveniles were deposited inthe tadpole collection of the Laboratorio deZoologia dos Vertebrados, housed at theUniversidade Federal de Ouro Preto, Muni-cipality of Ouro Preto, Brazil (LZVG 062–063), and one tadpole was deposited in theMuseu Nacional (MNRJ 42889). In ourfieldwork, which took place from May, 2004to August, 2006, we observed two couples thatspawned and we followed the development ofthe tadpoles. In describing the tadpoles, weused the developmental stages of Gosner(1960), the tooth row formula of Altig(1970), and other characters measured usedby Altig and McDiarmid (1999). We alsodescribe the tadpole coloration in life and inpreservative. Abbreviations for tadpole mea-surements are TL (total length), BL (bodylength), MBH (maximum body height), MBW(maximum body width), TAL (tail length),MTH (maximum tail height), ED (eye diam-eter), ND (nostril diameter), IOD (interorbit-al distance), IND (internarial distance), NSD(nostril snout distance), END (eye nostrildistance), ODW (oral disc width). We used acaliper and ocular grid (mm) in a stereomi-croscope for adult and tadpole measurements.All measurements are in millimeters.
SPECIES ACCOUNT
Scinax tripui sp. nov.
Holotype.—MNRJ 42890, an adult male(23.0 mm SVL), collected at Estacao Ecolo-gica do Tripuı (20u 239 220 S 43u 329 320 W,elevation 1070m), Municipality of Ouro Preto,State of Minas Gerais, Brazil, collected on 19August 2005 by A.C.C. Lourenco and D.Baeta (Figs. 1–3).
Paratypes.—MCNAM 7741 adult malecollected on 23 April 2004; MCNAM 7742–7744 adult males collected on 8 August 2004;MCNAM 7745–7746 adult males collected on15 April 2005; MNRJ 48763, MNRJ 48765–48767, MCNAM 7747–7749 adult malescollected in May 2005; MCNAM 7751–7754
TABLE 1.—Species assigned to the Scinax catharinaegroup.
Species
S. agilis (Cruz and Peixoto, 1983)S. albicans (Bokermann, 1967)S. angrensis (Lutz, 1973)S. argyreornatus (Miranda-Ribeiro, 1926)S. ariadne (Bokermann, 1967)S. aromothyella (Faivovich, 2005)S. berthae (Barrio, 1962)S. brieni (De Witte, 1930)S. canastrensis (Cardoso and Haddad, 1982)S. carnevallii (Caramaschi and Kisteumacher, 1989)S. catharinae (Boulenger, 1888)S. centralis (Pombal and Bastos, 1996)S. flavoguttatus (Lutz and Lutz, 1939)S. heyeri (Peixoto and Weygoldt, 1986)S. hiemalis (Haddad and Pombal, 1987)S. humilis (Lutz, 1954)S. jureia (Pombal and Gordo, 1991)S. kautskyi (Carvalho-e-Silva and Peixoto, 1991)S. littoralis (Pombal and Gordo, 1991)S. longilineus (Lutz, 1968)S. luizotavioi (Caramaschi and Kisteumacher, 1989)S. machadoi (Bokermann and Sazima, 1973)S. obtriangulatus (Lutz, 1973)S. ranki (Andrade and Cardoso, 1987)S. rizibilis (Bokermann, 1964)S. strigilatus (Spix, 1824)S. trapicheiroi (Lutz, 1954)
December 2009] HERPETOLOGICA 469

adult males collected on 18 May 2005;MCNAM 7750 collected on 17 June 2005;MNRJ 48762 adult male collected on 3 July2005; MCNAM 7756, adult female collectedon 24 July 2005; MNRJ 48764 adult malecollected on 25 July 2005; MCNAM 7755adult female collected on 19 August 2005;MCNAM 7758 adult female collected on 3September 2005; MCNAM 7759–7761 adultmales collected on 14 October 2006; MNRJ48,744–48,745 adult males collected on 2 July2007; MNRJ 48743 adult female collected on2 July 2007, from the same locality as theholotype. All of these specimens were collect-ed by A.C.C. Lourenco, D. Baeta, L. O.Drummond, M. D. Cid Brum, V. L. Guedes,and V. S. Monteiro.
Diagnosis.—A medium-sized species ofScinax belonging to the Scinax catharinaespecies group (Faivovich et al., 2005) charac-terized by: (1) snout subovoid in dorsal view;(2) canthus rostralis slightly concave and wellmarked; (3) dorsum surface brown withscattered dark brown and green blotches; (4)nuptial pads developed, but nonhypertro-phied; (5) interorbital region with a W-shapedmark posteriorly; (6) light green coloration onhidden regions of flanks and thighs of adults;(7) belly and throat cream with great densityof dark brown spots.
Comparison with other species.—The newspecies differs from S. agilis, S. argyreornatus,
S. berthae, S. hiemalis, S. luizotavioi, S.machadoi, and S. rizibilis by its larger femalesize, and from S. argyreornatus, S. berthae,and S. machadoi by its larger male size(Table 2). Scinax tripui differs from S. albi-cans, S. catharinae, and S. flavoguttatus by itssmaller female size, and from S. albicans, S.ariadne, S. catharinae, and S. flavoguttatus byits smaller male size (Table 2). The subovoidsnout in dorsal view distinguishes S. tripuifrom S. agilis, S. albicans, S. angrensis, S.argyreornatus, S. ariadne, S. aromothyella, S.berthae, S. brieni, S. canastrensis, S. carne-vallii, S. catharinae, S. centralis, S. flavogut-tatus, S. hiemalis, S. humilis, S. jureia, S.kautskyi, S. littoralis, S. longilineus, S. luizo-tavioi, S. machadoi, S. ranki, S. rizibilis, S.strigilatus, and S. trapicheiroi (mucronate inS. kautskyi and S. littoralis; rounded in S.albicans, S. ariadne, S. berthae, S. brieni, S.carnevallii, S. catharinae, S. flavoguttatus, S.jureia, S. ranki, S. strigilatus, and S. trapi-cheiroi; sub-elliptical in S. agilis, S. angrensis,S. argyreornatus, S. aromothyella, S. canas-trensis, S. centralis, S. hiemalis, S. humilis, S.longilineus, S. luizotavioi, and S. rizibilis;truncate in S. machadoi). The well-definedcanthus rostralis differentiates S. tripui fromS. brieni, S. catharinae, S. heyeri (Peixoto andWeygoldt, 1986), S. jureia, S. machadoi, S.ranki (canthus rostralis poorly defined inthese species). Scinax tripui is distinguished
FIG. 1.—Dorsal and ventral views of Scinax tripui sp. nov., MNRJ 42890 (holotype, 23.0 mm SVL).
470 HERPETOLOGICA [Vol. 65, No. 4

from S. centralis and S. hiemalis by its discreteinguinal gland (prominent and enlarged inthese species). The presence of a nuptial pad,developed and nonhypertrophied, sets S.tripui apart from almost all species of the S.catharinae group, except S. albicans, S. agilis,S. ariadne, S. aromothyela, S. catharinae, S.kautsky, and S. trapicheiroi (hypertrophied inS. rizibilis; not developed in all others species)(Fig. 3). Vomerine teeth in two straight seriesdistinguish the new species from S. agilis, S.angrensis, S. argyreornatus, S. ariadne, S.canastrensis, S. kautskyi, S. littoralis, and S.
longilineus (the vomerine teeth are oblique inS. agilis, S. angrensis, S. argyreornatus, S.kautskyi, S. littoralis, and S. longilineus, andare convex in S. ariadne and S. canastrensis).In life, a light green inguinal region andhidden portions of flanks and thighs differen-tiates S. tripui from S. argyreornatus, S.aromothyella (Faivovich et al., 2005), S.berthae (Barrio, 1962), S. canastrensis (Car-doso and Haddad, 1982), S. centralis (Pombaland Bastos, 1996), S. flavoguttatus, S. heyeri(Peixoto and Weygoldt, 1986), S. longilineus,S. machadoi (Bokermann and Sazima, 1973),
FIG. 2.—Scinax tripui sp. nov., holotype (MNRJ 42890). (A) dorsal and (B) lateral views of the head (scale 5 5 mm),and ventral views of (C) hand, and (D) foot (scale 5 1 mm).
December 2009] HERPETOLOGICA 471

which show orange or yellow color in thesebody parts; S. brieni (Lutz, 1973), S. humilis(Lutz, 1954; Lutz, 1973), S. obtriangulatus(Lutz, 1973), and S. trapicheiroi, which showblue color in theses body parts; S. ariadne andS. catharinae, which show grayish violet colorin these body parts.
Description of holotype.—Body slender andmedium size. Head longer than wide (36% ofSVL). Snout subovoid in dorsal view andprotruding in profile. Nostrils located lateral-
ly, immediately before the tip of snout,opening directed dorsolaterally, elliptical,and protruding. Canthus rostralis distinct,well marked. Loreal region concave. Eyelarge, protruding laterally, diameter about36% of head’s width. Interorbital and inter-nostril distance about 38.7% and 23.9% ofhead’s width respectively. Tympanum large,ovoid, and well defined, except at the upperedge, about 34% of eye’s diameter. Supra-tympanic fold well defined and granulated.
FIG. 3.—Degree of development of the nuptial pad: (A) Scinax tripui, holotype (MNRJ 42890); (B) S. flavoguttatus(MNRJ 51483); and (C) S. rizibilis (MNRJ 17654). Arrows point to nuptial pads.
TABLE 2.—Range of measurements of snout vent length of the species of the Scinax catharinae group which werecompared with those of S. tripui (differences were considered significant when P was ,0.05).
Species Sex Range N P
Scinax ariadne Male 35.0–38.1 8 ,0.001Scinax argyreornatus Male 14.1–17.3 33 ,0.001Scinax albicans Male 26.6–32.0 22 ,0.001Scinax berthae Male 18.0–21.9 9 ,0.001Scinax catharinae Male 30.0–37.6 9 ,0.005Scinax flavoguttatus Male 25.0–30.0 22 ,0.001Scinax machadoi Male 15.0–16.8 6 ,0.001Scinax agilis Female 14.0–18.6 8 ,0.001Scinax argyreornatus Female 17.1–25.8 12 ,0.001Scinax albicans Female 39.0–43.8 8 ,0.005Scinax berthae Female 21.6–24.6 5 ,0.001Scinax catharinae Female 40.1–45.5 5 ,0.001Scinax flavoguttatus Female 40.0–45.4 10 ,0.001Scinax hiemalis Female 29.6–34.4 9 ,0.005Scinax humilis Female 34.0–40.9 9 ,0.5Scinax luizotavioi Female 26.3–30.0 7 ,0.001Scinax rizibilis Female 27.0–32.4 5 ,0.001
472 HERPETOLOGICA [Vol. 65, No. 4

Tongue large, cordiform, and unattached atthe posterior and lateral borders. Vomerineteeth disposed in two straight, transverseseries, each sets with five teeth. Arms slender,with longer forearms than upper arms. Handsabout 33.2% of snout vent length. Innermetacarpal tubercle elliptical and outer meta-carpal heart-shaped. Single and conical sub-articular tubercles on fingers II and III,rounded on IV and V. Supernumerary tuber-cles dispersed and small. Nuptial pads devel-oped. Webbing absent between fingers IV andV, extremely reduced between fingers II andIII, and III and IV. Elliptical finger discswider than long. Length of fingers II , III ,V , IV. Foot about 43.7% of snout vent length.Outer and inner metatarsal tubercles single andelliptical, inner larger than outer. Single, conicalsubarticular tubercles on toes. Supernumerarytubercles dispersed and small. Length of toes I, II , II , V , IV. Toe webbing formulaI-II11/2-21/2III11/2-21/2IV21/2-1+V. Toe discs el-liptical, wider than long.
Measurements of holotype.—SVL 23.0, HW7.9, HL 8.5, END 2.9, IND 2.0, ED 2.8, ESD5.0, IOD 2.8, TD 0.7, AL 5.2, TL 11.9, SL12.8, FL 10.4, FW 1.3, and TW 1.2.
Color of holotype in life.—Dorsal surfacelight brown with dark brown stripes. Snoutlight brown with irregular dark brown blotch-
es. Interocular surface with a dark brownblotch, bordered by an anterior lighter brownstripe and with a W-shaped mark at theposterior edge. Granules on dorsal surfacedark brown and green, giving a mossy aspect.Dorsal surface with a pair of dark brownstripes and two blotches of inverse-parenthe-ses shape, bordered by a light brown line.Canthus rostralis with a longitudinal darkbrown stripe. In lateral view, alternating darkbrown and light brown vertical bars extendingfrom the inferior margin of the eye to theborder of the upper lips. Dorsal surface ofarms, hands, thighs, shanks, and feet withtransverse dark brown bars on a light brownbackground. Hidden portions of flanks andthighs light green. Ventral surfaces scatteredwith brown dots on a cream background.Abdominal surface more pigmented thangular. Iris brown with dense gold dots.
Color of holotype in preservative.—Patternof stripes, blotches and dots on dorsal, lateraland ventral surfaces remain in preservative,except for the green blotches which fade and forthe interocular blotch that becomes less evi-dent. Iris grayish, gold dots lost in preservative.
Variation.—Measurements of 32 adultmales and five adult females are presentedin Table 3. Females are much larger thanmales; male size represents approximately
TABLE 3.—Mean, standard deviation (SD), and range of measurements of 32 males and 5 females of Scinax tripui(abbreviations defined in text).
Male Female
Mean SD Range Mean SD Range
SVL 23.6 1.27 21.31–26.18 38.7 0.8 37.0–39.2HW 8.0 0.6 7.1–9.4 13.3 0.1 13.2–13.4HL 8.4 0.6 7.3–9.8 13.3 0.0 13.3–13.4END 2.6 0.2 1.9–3.0 4.0 0.3 3.5–4.3NW 0.4 0.1 0.3–0.5 0.5 0.1 0.4–0.8IND 2.2 0.1 1.9–2.5 3.2 0.3 2.8–3.6ED 2.9 0.2 2.5–3.3 4.6 0.5 3.7–5.0ESD 4.1 0.4 3.1–5.0 5.8 0.4 5.1–6.4IOD 3.1 0.2 2.9–3.5 4.6 0.7 3.9–6.0TD 1.0 0.1 0.7–1.2 1.5 0.1 1.4–1.6AL 7.8 0.5 6.9–9.0 12.9 0.1 12.2–13.5UL 4.8 0.4 4.0–5.7 8.2 0.4 7.9–8.6HAL 7.8 0.5 6.8–8.8 12.2 0.3 11.8–12.4TL 11.6 1.0 10.2–16.0 18.7 0.6 18.2–19.3SL 12.5 0.7 11.3–14.1 20.2 0.5 19.8–20.7TAL 7.1 1.0 6.5–12.2 10.5 0.5 10.4–11.4FL 10.3 0.6 9.3–11.4 16.8 0.4 16.6–17.2FW 1.4 0.1 1.08–1.65 2.27 0.0 2.2–2.3TW 1.3 0.1 1.08–1.54 1.97 0.1 1.9–2.0
December 2009] HERPETOLOGICA 473
61% female size. Males are slenderer thanfemales. Mossy aspect of dorsal surface ismore or less evident among the specimens.Greenish coloration in hidden regions offlanks and thighs can be a little lighter insome individuals, and is commonly moreevident in females. Each set of vomerineteeth may have four or five teeth. Males andfemales present variable webbing formulae:I-II11/2-21/2III11/2-21/2IV21/2-1+V, I-II11/2-21/2
III11/2-32IV21/2-1+V, I-II2232III11/2-32IV3211/2V,I-II1+-21/2III1+-21/2IV21/2-1+V or I-II11/2-21/2III1+-21/2IV21/2-1+V.
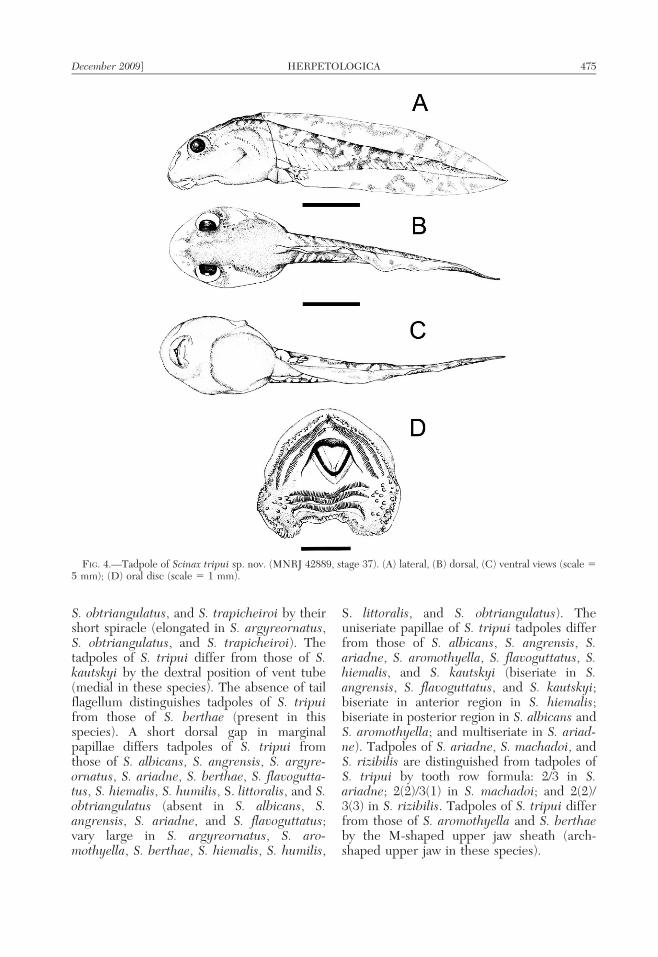
Description of tadpole.—Measurementsfrom nine tadpoles are presented in Table 4.Body oval in dorsal view and globular inlateral view (Fig. 4). Snout rounded in dorsaland lateral views. Eyes dorsolateral, eyediameter 33.3% of interorbital distance. Nos-trils rounded in dorsal view, with relativelysmall openings, almost as close to the eye as tothe snout. Spiracle lateral, sinistral, short,directed upwards and backwards, with a largeopening at the end of the middle third of thebody, on the lateral midline, and inner wallpresenting a slight ridge. Vent tube dextral,fused to the ventral fin, with dextral opening.Tail length 66.6% of the total length, higherthan the body. Dorsal fin beginning onposterior third of body. Dorsal fin convexand ventral fin with a rectilinear contour,dorsal fin somewhat higher than ventral. Oraldisc ventral (Fig. 4), not emarginated, with anarrow dorsal gap, surrounded by a uniseriaterow of papillae, in alternate disposition.
Individual submarginal papillae dispersedlaterally. Tooth row formula 2(2)/3, first andsecond anterior tooth row with same width,longer than posterior teeth rows, posteriorrows of same width. Upper and lower jawsheaths finely serrated, upper M-shaped andlower V-shaped.
Coloration of tadpole in life.—Dorsal sur-face of body brown with many fine gold dotsdispersed homogeneously, especially betweennostrils and eyes. Snout translucent andlighter than body. Eyes golden with darkbrown dots. Laterally, body the same patternas dorsum, except for a reddish coloration inthe gill region. Ventrally, abdominal surfacetranslucent with reddish brown dots. Venttube transparent with brown dots. Tail opaquewith brown dots and spots, more abundant onmusculature. At stage 37, the tadpoles withbrown transverse stripes on legs, same as onadult. Upper and lower jaw sheaths black.
Coloration in preservative.—Dorsal andlateral surfaces gray, without transparency,glossy aspect, or reddish and golden colora-tion. Body in ventral view opaque. Spots anddots of the tail the same color as in life, butless distinct. Eyes gray.
Comparison with tadpoles of the S.catharinae group.—Tadpoles of Scinax tripuiare distinguished from those of S. angrensis,S. argyreornatus, S. aromothyella, S. berthae,S. flavoguttatus, S. kautskyi, S. longilineus, S.machadoi, S. ranki, and S. rizibilis by dorsalposition of the nostrils (anterolateral in S.angrensis and S. kautskyi; dorsolateral in theothers species). Gold spots between nostrilsand eyes differentiate Scinax tripui tadpolesfrom those of S. argyreornatus, S. ariadne, S.berthae, S. catharinae, S. hiemalis, S. humilis,S. littoralis, S. obtriangulatus, S. ranki, and S.rizibilis (absent in these species). Scinax tripuitadpoles are distinguished from those of S.aromothyella, S. berthae, S. kautskyi, S.machadoi, and S. ranki by the presence ofdorsolateral eyes (lateral in these species).Except for S. catharinae, tadpoles of S. tripuiare distinguished from the other describedtadpoles of the group by their spiracle openingon the midline of dorsal and ventral surface ofthe body (spiracle opening below the bodymidline in these species). Scinax tripuitadpoles differ from those of S. argyreornatus,
TABLE 4.—Range, mean, and standard deviation (SD) ofmeasurements (mm) of Scinax tripui tadpoles (n 5 9,
stages 36–37) (abbreviation defined in text).
Mean SD Range
TL 37.5 2.8 33.7–41.0BL 11.7 0.9 10.3–12.9MBH 7.0 0.8 5.7–8.0MBW 7.4 0.8 6.4–8.6TAL 25.0 2.6 21.4–29.1MTH 7.8 1.1 6.4–9.3ED 1.9 0.1 1.7–2.1ND 0.4 0.0 0.3–0.4IOD 6.2 0.8 5.1–7.9IND 3.1 0.2 2.7–3.4NSD 2.3 0.3 1.9–2.7END 2.2 0.2 1.8–2.5ODW 3.3 0.7 2.0–4.9
474 HERPETOLOGICA [Vol. 65, No. 4
S. obtriangulatus, and S. trapicheiroi by theirshort spiracle (elongated in S. argyreornatus,S. obtriangulatus, and S. trapicheiroi). Thetadpoles of S. tripui differ from those of S.kautskyi by the dextral position of vent tube(medial in these species). The absence of tailflagellum distinguishes tadpoles of S. tripuifrom those of S. berthae (present in thisspecies). A short dorsal gap in marginalpapillae differs tadpoles of S. tripui fromthose of S. albicans, S. angrensis, S. argyre-ornatus, S. ariadne, S. berthae, S. flavogutta-tus, S. hiemalis, S. humilis, S. littoralis, and S.obtriangulatus (absent in S. albicans, S.angrensis, S. ariadne, and S. flavoguttatus;vary large in S. argyreornatus, S. aro-mothyella, S. berthae, S. hiemalis, S. humilis,
S. littoralis, and S. obtriangulatus). Theuniseriate papillae of S. tripui tadpoles differfrom those of S. albicans, S. angrensis, S.ariadne, S. aromothyella, S. flavoguttatus, S.hiemalis, and S. kautskyi (biseriate in S.angrensis, S. flavoguttatus, and S. kautskyi;biseriate in anterior region in S. hiemalis;biseriate in posterior region in S. albicans andS. aromothyella; and multiseriate in S. ariad-ne). Tadpoles of S. ariadne, S. machadoi, andS. rizibilis are distinguished from tadpoles ofS. tripui by tooth row formula: 2/3 in S.ariadne; 2(2)/3(1) in S. machadoi; and 2(2)/3(3) in S. rizibilis. Tadpoles of S. tripui differfrom those of S. aromothyella and S. berthaeby the M-shaped upper jaw sheath (arch-shaped upper jaw in these species).
FIG. 4.—Tadpole of Scinax tripui sp. nov. (MNRJ 42889, stage 37). (A) lateral, (B) dorsal, (C) ventral views (scale 55 mm); (D) oral disc (scale 5 1 mm).
December 2009] HERPETOLOGICA 475

Distribution.—The new species is currentlyknown only from the streams of the EstacaoEcologica do Tripuı, Municipality of OuroPreto, State of Minas Gerais, Brazil, located inthe Espinhaco mountain range (Fig. 5).
Natural History.—Specimens were ob-served mainly in the dry and cold season,from April to September, a time when malescan be observed calling. They were generallyassociated with roots or leaves on the edge ofstreams. Males sang very close to theirneighbors, about one meter distance betweenthem. Eggs of this species were found in calmplaces of the stream, and were deposited onthe substrate bottom. Tadpoles were inactiveduring the entire night, hiding under leavesand rocks; they emerged only during the dayand were found close to the surface. Tadpolestook approximately one month to reach thejuvenile stage, and most specimens wereobserved in the juvenile stage at the end ofAugust.
Etymology.—The specific epithet tripui isderived from the Tupi–Guarani term Ityra–poi. The name is intended to mean ‘‘slenderhill’’ (Ityra 5 hill, poi 5 slender) and is thename of the main stream of the EstacaoEcologica do Tripuı, the type locality of thedescribed species.
DISCUSSION
The light green coloration on the hiddenareas of flanks and thighs is a diagnosticcharacteristic for Scinax tripui. The species of
the S. catharinae clade have been historicallydescribed and distinguished from each otherby the vivid colors on the concealed bodyparts (Carvalho-e-Silva and Peixoto, 1991;Cochran, 1955; da Silva and Alves-Silva,2008; Haddad and Pombal, 1987; Lutz,1973; Pimenta et al., 2007). Bertha Lutz(1973) defined two groups of species basedon this color pattern: (1) species with yellow-orange flash colors, and (2) species with blueto violet flash color. The color on concealedparts of flanks and thighs is an importantdiagnostic character for this clade and doesnot vary in the same species (A.C.C.L.personal observation).
Currently, Scinax flavoguttatus is knownfrom the coastal range in the states of Rio deJaneiro and Sao Paulo, and in a few localitiesin the mountain ranges in the state of MinasGerais (Cochran, 1955; Cruz et al., 2009;Haddad et al., 2008; Lutz, 1973; Pedralli et al.,2001). According to Pedralli et al. (2001),three species of the Scinax catharinae groupoccur in the region of Ouro Preto: S.flavoguttatus, S. longilineus, and S. luizotavioi.In our fieldwork at the same locality betweenMay 2004 and August 2006, we did not findany individuals of S. flavoguttatus and alwaysfound specimens of S. tripui. Afterwards, thespecimens previously identified as S. flavo-guttatus by Pedralli et al. (2001) wererecognized as the new species described here(A. S. Guimaraes-Neto personal communica-tion). It was impossible to perform comparisonsbetween S. tripui and the specimens collectedby Pedralli et al. (2001), because no individualsexamined by these authors were housed in anymuseum collection. Although the new specieswas previously identified as S. flavoguttatus,noticeable differences can be found betweentheses species, such as the distinct color on thehidden areas of flanks and thighs, the develop-ment of the nuptial pad, and the largedifference in adult body size.
Recent work has enlarged the geographicaldistribution of a great number of endemicspecies from the type locality at Minas GeraisState (e.g., Baeta et al., 2005; Santos et al.,2009). Although data on the anuran fauna ofsoutheastern Brazil has rapidly improved inthe last years, several areas have not been wellsampled for the anuran fauna. Therefore,
FIG. 5.—Geographical distribution of Scinax tripui sp.nov. in southeastern Brazil (asterisk represents EstacaoEcologica do Tripuı, Municipality of Ouro Preto, MinasGerais, the type locality of this species).
476 HERPETOLOGICA [Vol. 65, No. 4

future work in the Espinhaco mountain rangecould enlarge the distribution of Scinax tripuiand S. flavoguttatus.
RESUMO
Neste artigo, uma nova especie do grupo deScinax catharinae e descrita do municıpio deOuro Preto, sudeste do Brasil. A nova especiee relacionada a Scinax flavoguttatus, masdifere-se desta, especialmente, pela coloracaoverde clara na regiao inguinal e nas areasocultas dos flancos e coxas, pela almofadanupcial desenvolvida e por apresentar machose femeas menores. Adicionalmente, apresen-tamos a descricao do girino.
Acknowledgments.—We would like to thank R. W.Heyer and C. A. G. Cruz for critically reading themanuscript; P. R. Nascimento for the line drawings; D.Baeta, L. O. Drummond, M. D. Cid Brum, V. L. Guedes,and V. S. Monteiro for help in fieldwork; J. A. RodriguesLoes, from the Instituto Estadual de Florestas (IEF–MG)for logistical support; Instituto Brasileiro do MeioAmbiente e dos Recursos Naturais Renovaveis (IBAMA;License of IBAMA 153/05–NUFAS/MG; process number02015.005026/05–23) and Instituto Estadual de Florestasfor collection permission. For financial support theauthors are grateful to FAPERJ; A.C.C. Lourenco toPrograma de Iniciacao Cientıfica/UFOP (PIP/UFOP) andConselho Nacional de Pesquisa (CNPq); and L.B.Nascimento to Fundo de Incentivo a Pesquisa (FIP/PUCMInas) and to FAPEMIG.
LITERATURE CITED
ALTIG, R. 1970. A key to the tadpoles of the continentalUnited States and Canada. Herpetologica 26:180–207.
ALTIG, R., AND R. W. MCDIARMID. 1999. Body plan:development and morphology. Pp. 24–51. In R. W.McDiarmid and R. Altig (Eds.), Tadpoles: The Biologyof Anuran Larvae. The University of Chicago Press,Chicago, Illinois, USA.
ANDRADE, G. V., AND A. J. CARDOSO. 1987. Reconheci-mento do grupo rizibilis e descricao de uma novaespecie de Hyla (Amphibia, Anura). Revista Brasileirade Zoologia 3:433–440.
ANDRADE, G. V., AND A. J. CARDOSO. 1991. Descricao delarvas e biologia de quatro especies de Hyla (Amphibia,Anura). Revista Brasileira de Biologia 51:391–402.
BARRIO, A. 1962. Los Hylidae de Punta lara, Provıncia deBuenos Aires. Physis 23:129–142.
BAETA, D., P. H. BERNARDO, B. ASSIS, V. A. SAO PEDRO,L. O. DRUMMOND, AND M. R. S. PIRES. 2005.Geographic Distribuition: Physalaemus maximus. Her-petological Review 36:200–200.
BOKERMANN, W. C. A. 1964. Uma nova especie de ‘‘Hyla’’da serra do mar em Sao Paulo. Revista Brasileira deBiologia 24:429–434.
BOKERMANN, W. C. A. 1967. Dos nuevas riadne de Hyla rigrupo catharinae (Amphibia, Hylidae). Neotropica13:62–66.
BOKERMANN, W. C., AND I. SAZIMA. 1973. Anfıbios da Serrado Cipo, Minas Gerais, Brasil. 1: duas novas especies deHyla (Anura, Hylidae). Revista Brasileira de Biologia33:457–472.
BOULENGER, G. A. 1888. A list of batrachians from theprovince of Santa Catharina, Brazil. Annals andMagazine of Natural History 6:415–417.
CARAMASCHI, U., AND G. KISTEUMACHER. 1989. Duas novasespecies de Ololygon Fitzinger, 1843, do Sudeste doBrasil (Amphibia, Anura, Hylidae). Boletim do MuseuNacional, Nova Serie, Zoologia 327:1–15.
CARDOSO, A. J., AND C. F. B. HADDAD. 1982. Nova especiede Hyla da Serra da Canastra (Amphibia, Anura,Hylidae). Revista Brasileira de Biologia 42:499–503.
CARVALHO-E-SILVA, S. P. 1986. Girinos de especies doGenero Ololygon pertencentes ao grupo ‘‘catharinae’’,no Estado do Rio de Janeiro (Amphibia, Anura,Hylidade). Dissertacao de mestrado, Museu Nacional,Universidade Federal do Rio de Janeiro, Brazil.
CARVALHO-E-SILVA, S. P., AND O. L. PEIXOTO. 1991. Duasnovas especies de Ololygon para os Estados do Rio deJaneiro e Espırito Santo (Amphibia, Anura, Hylidae).Revista Brasileira de Biologia 51:263–270.
CARVALHO-E-SILVA, S. P., AND A. M. P. T. CARVALHO-E-
SILVA. 1994. Descricao das larvas de Ololygon albicans ede Ololygon trapicheiroi com consideracoes sobre suabiologia (Amphibia, Anura, Hylidae). Revista Brasileirade Biologia 54:55–62.
CARVALHO-E-SILVA, S. P., AND A. M. P. T. CARVALHO-E-
SILVA. 1998. Aspects of the biology and description ofthe larvae of Scinax argyreornatus and Scinax humilis(Amphibia: Anura: Hylidae). Reveue Francesa Aqua-riologia 25:47–52.
CARVALHO-E-SILVA, S. P., M. M. DOS REIS GOMES, AND O. L.PEIXOTO. 1995. Descricao dos girinos de Scinaxangrensis (B. Lutz, 1973) e de Scinax kautskyi(Carvalho-e-Silva e Peixoto, 1991) (Amphibia, Anura,Hylidae). Revista Brasileira de Biologia 55:61–65.
COCHRAN, D. M. 1955. Frogs of Southeastern Brazil.United States National Museum Bulletin 206:1–423.
CONTE, C. E., F. NOMURA, D. C. ROSSA-FERES, A.D’HEURSEL, AND C. F. B. HADDAD. 2007. The tadpoleof Scinax catharinae (Anura: Hylidae) with descriptionof the internal oral morphology, and a review of thetadpoles from the Scinax catharinae group. Amphibia–Reptilia 28:177–192.
CRUZ, C. A. G., AND O. L. PEIXOTO. 1983. Uma novaespecie de Hyla do Estado do Espırito Santo, Brasil(Amphibia, Anura, Hylidae). Revista Brasileira deBiologia 42:721–724.
CRUZ, C. A. G., R. N. FEIO, AND U. CARAMASCHI. 2009.Anfıbios do Ibitipoca, Bicho do Mato Ed., BeloHorizonte, Minas Gerais, Brazil.
DE WITTE, G. F. 1930. Liste des reptiles et batraciensrecoltes au Bresil par la Mission Massart (1922–23) etdescription de sept nouvelles especes. Pp. 213–230. InJ. Massart (Ed.), Une mission biologique belge auBresil 2.
DA SILVA, H. R., AND R. ALVES-SILVA. 2008. New coastaland insular species of the bromeligenous Scinaxperpusillus group, from the State of Rio de Janeiro,Brazil (Anura, Hylidae). Zootaxa 1914:34–44.
December 2009] HERPETOLOGICA 477

DE SA, R. O., R. WASSERSUG, AND A. I. KEHR. 1997.Description of tadpoles of three species of Scinax(Anura: Hylidae). Herpetologica 7:13–17.
DUELLMAN, W. E. 1970. The hylid frogs of MiddleAmerica. Monograph of the Museum of NaturalHistory, University of Kansas 21:1–372.
FABREZI, M., AND P. ALBERCH. 1996. The carpal elementsof anurans. Herpetologica 52:188–204.
FAIVOVICH, J. 2002. A cladistic analysis of Scinax (Anura,Hylidae). Cladistics 18:367–393.
FAIVOVICH, J. 2005. A new species of Scinax (Anura:Hylidae) from Misiones, Argentina. Herpetologica61:69–77.
FAIVOVICH, J., C. F. B. HADDAD, P. C. A. GARCIA, D. R.FROST, J. A. CAMPBELL, AND W. C. WHEELER. 2005.Sistematic review of the frog family Hylidae, withspecial reference to Hylinae: Phylogenetic analysis andtaxonomic revision. Bulletin of the American Museumof Natural History 294:1–240.
FROST, D. R. 2009. Amphibian species of the world: Anonline reference. Version 5.3 (12 February, 2009).Electronic database accessible at http://research.amnh.org/herpetology/amphibia/index.php. American Muse-um of Natural History, New York, New York, USA.
GOSNER, K. L. 1960. A simplified table for staging anuranembryo and larvae with notes on identification.Herpetologica 16:183–190.
HADDAD, C. F. B., AND J. P. POMBAL, JR. 1987. Hylahiemalis, nova especie do grupo rizibilis do Estado deSao Paulo (Amphibia, Anura, Hylidae). Revista Brasi-leira de Biologia 47:127–132.
HADDAD, C. F. B., L. F. TOLEDO, AND C. P. A. PRADO.2008. Anfıbios da Mata Atlantica, Neotropica Ed., SaoPaulo, Sao Paulo, Brazil.
HEYER, W. R., A. S. RAND, C. A. G. CRUZ, O. L. PEIXOTO,AND C. E. NELSON. 1990. Frogs of Boraceia. Arquivos deZoologia 31:237–410.
LUTZ, B. 1954. Anfıbios anuros do Distrito Federal.Memorias do Instituto Oswaldo Cruz 52:155–238.
LUTZ, B. 1973. Brazilian species of Hyla. University ofTexas Press, Austin and London.
LUTZ, A., AND B. LUTZ. 1939. New Hylidae from Brazil.Hilideos novos do Brasil. Anais da Academia Brasileirade Ciencias XI:67–89.
MIRANDA-RIBEIRO, A. 1926. Notas para servirem ao estudodos gymnobatrachios (Anura) brasileiros. Arquivos doMuseu Nacional 27:1–227.
MORAVEC, J., I. A. TUANAMA, P. E. PEREZ, AND E. LEHR.2009. A new species of Scinax (Anura: Hylidae) fromthe area of Iquitos, Amazonian Peru. South AmericanJournal of Herpetology 4:9–16.
PEDRALLI, G., A. S. GUIMARAES NETO, AND M. C. B.TEIXEIRA. 2001. Diversidade de anfıbios na regiao deOuro Preto. Ciencia Hoje 30:70–73.
PEIXOTO, O. L. 1987. Caracterizacao do grupo ‘‘perpusilla’’e revalidacao da posicao taxonomica de Ololygonperpusilla perpusilla e Ololygon perpusilla v-signata(Amphibia, Anura, Hylidae). Arquivos da UniversidadeFederal Rural do Rio de Janeiro 10:37–49.
PEIXOTO, O. L., AND P. WEYGOLDT. 1987. Notes onOlolygon heyeri Weygoldt 1986 from Espırito Santo,Brazil (Amphibia: Salientia: Hylidae). SenckenbergianaBiologica 68:1–9.
PIMENTA, B. V. S., J. FAIVOVICH, AND J. P. POMBAL, JR. 2007.On the identity of Hyla strigilata Spix, 1824 (Anura;Hylidae): redescription and neotype designation for a‘‘ghost’’ taxon. Zootaxa 1441:35–49.
POMBAL, J. P., JR., AND M. GORDO. 1991. Duas novasespecies de Hyla da Floresta Atlantica no Estado de SaoPaulo (Amphibia, Anura). Memorias do InstitutoButantan 53:135–144.
POMBAL, J. P., JR., AND R. P. BASTOS. 1996. Nova especie deScinax Wagler, 1830 do Brasil Central (Amphibia,Anura, Hylidae). Boletim do Museu Nacional, NovaSerie, Zoologia 371:1–11.
SANTOS, J. T., E. F. OLIVEIRA, V. A. SAO PEDRO, A. C.MONTEIRO-LEONEL, AND R. N. FEIO. 2009. Amphibia,Anura, Hypsiboas stenocephalus: Distribution extensionand geographic distribution map. Check List 5:27–31.
SPIX, J. B. 1824. Animalia nova sive species novaeTestudinum et Ranarum, quas in itinere per Brasiliamannis MDC–CCXVII–MDCCCXX jussu et auspiciisMaximiliani Josephi I. Bavariae Regis. Typis FrancisSeraphici Hubschmanni, Monachii.
.Accepted: 16 November 2009
.Associate Editor: Brad Moon
APPENDIX I
Additional Specimens Examined
Adults—Scinax agilis: BRAZIL, State of Bahia: PortoSeguro (MNRJ 29803), Belmonte (MNRJ 46853–54);State of Espırito Santo: Linhares (MNRJ 4146 and MNRJ14210–13, paratypes); Guarapari (MNRJ 43094–95).Scinax albicans: BRAZIL, State of Rio de Janeiro:Parque Nacional da Serra dos Orgaos, Teresopolis (MNRJ4053, paratype, MNRJ 39930–35); Nova Friburgo (MNRJ23393–96); Cachoeira de Macacu (MNRJ 40080–82).Scinax angrensis: BRAZIL, State of Rio de Janeiro:Angra dos Reis (MNRJ 2018–2512); Manguaratiba(MNRJ 43504–08); Parati (MNRJ 44115–17). Scinaxargyreornatus: BRAZIL, State of Bahia: Porto Seguro(MNRJ 37929–30); Canavieiras (MNRJ 40303–08); Stateof Espırito Santo: Linhares (MNRJ 22967–23038); SantaTeresa (MNRJ 26100–31, MNRJ 38390–36); State of Riode Janeiro: Itaguaı (MNRJ 32036–42); Mage (MNRJ54987–95). Scinax riadne: BRAZIL, State of Sao Paulo:Sao Jose do Barreiro (MNRJ 4051, paratype, MNRJ43611); State of Rio de Janeiro: Teresopolis (MNRJ55654). Scinax aromothyella: ARGENTINA, ProvınciaMisiones: San Vicente (MNRJ 56445, paratype). Scinaxberthae: ARGENTINA, Province of Buenos Aires: PuntaLara (MNRJ 3590, paratype), San Isidro (MNRJ 59527–28); BRAZIL, State of Sao Paulo: Botucatu (MNRJ34761–65); State of Rio Grande do Sul: Santa Maria(MNRJ 39897). Scinax brieni: BRAZIL, State of SaoPaulo: Paranapiacaba (AL-MN 2592–94 topotypes). Sci-nax canastrensis: BRAZIL, State of Minas Gerais: SaoRoque de Minas (MNRJ 4147, holotype, MNRJ 4148,paratype). Scinax catharinae: BRAZIL, State of SantaCatarina: Guaratuba (MNRJ 1789), Corupa (MNRJ 168),Sao Bento do Sul (Al-001; MNRJ 44411–414, MNRJ
478 HERPETOLOGICA [Vol. 65, No. 4

1738, MNRJ 1801–1803, MNRJ 9475), Florianopolis(MNRJ 55639); State of Sao Paulo, Sao Jose do Barreiro(MNRJ 2084–87), Guaratuba (MNRJ 35106–107). Scinaxcarnevallii: BRAZIL, State of Minas Gerais: Marlieria(MNRJ 4182, holotype, MNRJ 4183–200, paratypes);Caratinga (MNRJ 4201–09). Scinax centralis: BRAZIL,State of Goias: Silvania (MNRJ 17465, holotype, MNRJ17466–17475, paratypes, MNRJ 32239–44, topotypes).Scinax flavoguttatus: BRAZIL, State of Sao Paulo:Serra da Bocaina, Sao Jose do Barreiro (AL-MN 2090,holotype, AL-MN 2091, paratype, MNRJ 23404–07,topotypes); Cambuci (MNRJ 51483–84); Duque de Caxias(MNRJ 53688–03); Parque Nacional do Itatiaia, Rio deJaneiro (MNRJ 59457); Petropolis (MNRJ 57575–76);Vale da Revolta, Teresopolis (MNRJ 53950–58); ReservaEcologica de Guapiacu, Cachoeira de Macacu (MNRJ46536, MNRJ 53311). Scinax hiemalis: BRAZIL, Stateof Sao Paulo: Campinas (MNRJ 5973–74, topotypes),Botucatu (MNRJ 30645–75; MNRJ 40008–09), SaoSebastiao (MNRJ 32530–32). Scinax humilis: BRAZIL,State of Rio de Janeiro: Duque de Caxias (MNRJ 1478,paralectotype); Nova Iguacu (MNRJ 2248, lectotype),Mangaratiba (MNRJ 39886–91), Guapimirim (MNRJ23411–12, MNRJ 40095–103, MNRJ 47987, MNRJ59027–29). Scinax jureia: BRAZIL, State of Sao Paulo:Estacao Ecologica Jureia-Itatins, Iguape (MNRJ 14202–03, paratypes). Scinax kautskyi: BRAZIL, State ofEspırito Santo: Reserva Biologica de Duas Bocas,
Cariacica (MNRJ 27889–30, MNRJ 27956). Scinaxlittoralis: BRAZIL, State of Sao Paulo: Estacao Ecolo-gica Jureia–Itatins, Iguape (MZUSP 73736–38, para-types). Scinax longilineus: BRAZIL, State of MinasGerais: Pocos de Caldas (MNRJ 4060, holotype, MNRJ40618, topotype); Belo Horizonte (MNRJ 16003–07,MNRJ 30966–69). Scinax luizotavioi: BRAZIL, Stateof Minas Gerais: Serra do Caraca, Catas Altas (MNRJ4210, holotype, MNRJ 4211–16, paratypes); Peti, SaoGoncalo do Rio Abaixo (MNRJ 4473–508 and MNRJ4509–16, paratypes, MNRJ 32462–74, MNRJ 36781–804;MNRJ 50619–21); MNRJ 52361–63, MNRJ 56475–78).Scinax machadoi: BRAZIL, State of Minas Gerais: Serrado Cipo, Jaboticatubas (MNRJ 17476–77, paratypes,MNRJ 39696, topotype). Scinax ranki: BRAZIL, Satateof Minas Gerais: Pocos de Caldas (MNRJ 49657topotype). Scinax rizibilis: BRAZIL, State of Sao Paulo:Ribeirao Branco (MNRJ 18224–25, MNRJ 17654);Parque Estadual Intervales, Ribeirao Grande (MNRJ28131–52); State of Santa Catarina: Serra Alta, RioVermelho (MNRJ 50150–97). Scinax strigilatus: BRA-ZIL, State of Bahia: Ibirapitanga, Fazenda Pedra Formosa(MNRJ 38098). Scinax obtriangulatus: BRAZIL, Stateof Rio de Janeiro: Alto Itatiaia, Brejo da Lapa (MNRJ4035, holotype of Hyla catharinae simplex). Scinaxtrapicheiroi: BRAZIL, State of Rio de Janeiro: Rio deJaneiro (MNRJ 3615–17, cotypes, MNRJ 3618–25,paratypes); Mage (MNRJ 56154–55).
DATE OF PUBLICATION
Herpetologica, Vol. 65, No. 4, was mailed 19 February 2010.
December 2009] HERPETOLOGICA 479


















