
A Gene Delivery Method Mediated by Three Arginine-rich ... · system became a novel tool for gene...
18
Advanced Studies in Biology, Vol. 5, 2013, no. 2, 71 - 88 HIKARI Ltd, www.m-hikari.com A Gene Delivery Method Mediated by Three Arginine-rich Cell-penetrating Peptides in Plant Cells Min-Jie Liu, Jyh-Ching Chou and Han-Jung Lee* Department of Natural Resources and Environmental Studies, National Dong Hwa University, Hualien 97401, Taiwan * Corresponding Author. [email protected] (H.-J.L.) Abstract The transient gene delivery systems are generally mediated by viral infection, particle bombardment, electroporation, and microinjection for transferring exogenous DNAs into host cells. Recently, the peptide-mediated DNA delivery system became a novel tool for gene transfer, and these peptides, such as cell-penetrating peptides (CPPs), contained the ability of permeating plasma membranes and carrying cargoes to enter cells. In this study, we use three different arginine-rich CPPs, synthetic nona-arginine (SR9), histidine-rich nona-arginine (HR9), and Pas nona-arginine (PR9), for gene delivery into root-tip cells of mung bean and rice. We demonstrate that these arginine-rich CPPs have the ability to transfer Cyanine 3 (Cy3)-labeled DNAs into plant cells. Moreover, the CPP-mediated DNA transfer in cells was confirmed by polymerase chain reaction (PCR) analysis. Finally, our results of the functional gene assay indicated that DNA exerts its biological activity after intracellular delivery via CPPs. This CPP-mediated gene delivery method is characterized by the simplicity, noninvasion, and efficiency. Three arginine-rich CPPs may be ideal tools for the research of plant gene transformation. Keywords: Cell-penetrating peptide (CPP), Green fluorescent protein (GFP), Protein transduction domain (PTD), Transgenic plants Abbreviations: CPP, cell-penetrating peptide; Cy3: Cyanine 3; GFP, green fluorescent protein; HR9: histidine - rich nona - arginine; N/P: nitrogen (NH 3 + )/phosphate (PO 4 – ); PCR: polymerase chain reaction; PR9: Pas nona-arginine; RFP: red fluorescent protein; SR9: synthetic nona-arginine; Tat: transactivator of transcription
Transcript of A Gene Delivery Method Mediated by Three Arginine-rich ... · system became a novel tool for gene...

Advanced Studies in Biology, Vol. 5, 2013, no. 2, 71 - 88
HIKARI Ltd, www.m-hikari.com
A Gene Delivery Method Mediated by
Three Arginine-rich Cell-penetrating Peptides
in Plant Cells
Min-Jie Liu, Jyh-Ching Chou and Han-Jung Lee*
Department of Natural Resources and Environmental Studies, National Dong Hwa University, Hualien 97401, Taiwan
* Corresponding Author. [email protected] (H.-J.L.)
Abstract
The transient gene delivery systems are generally mediated by viral infection, particle bombardment, electroporation, and microinjection for transferring exogenous DNAs into host cells. Recently, the peptide-mediated DNA delivery system became a novel tool for gene transfer, and these peptides, such as cell-penetrating peptides (CPPs), contained the ability of permeating plasma membranes and carrying cargoes to enter cells. In this study, we use three different arginine-rich CPPs, synthetic nona-arginine (SR9), histidine-rich nona-arginine (HR9), and Pas nona-arginine (PR9), for gene delivery into root-tip cells of mung bean and rice. We demonstrate that these arginine-rich CPPs have the ability to transfer Cyanine 3 (Cy3)-labeled DNAs into plant cells. Moreover, the CPP-mediated DNA transfer in cells was confirmed by polymerase chain reaction (PCR) analysis. Finally, our results of the functional gene assay indicated that DNA exerts its biological activity after intracellular delivery via CPPs. This CPP-mediated gene delivery method is characterized by the simplicity, noninvasion, and efficiency. Three arginine-rich CPPs may be ideal tools for the research of plant gene transformation. Keywords: Cell-penetrating peptide (CPP), Green fluorescent protein (GFP), Protein transduction domain (PTD), Transgenic plants Abbreviations: CPP, cell-penetrating peptide; Cy3: Cyanine 3; GFP, green fluorescent protein; HR9: histidine - rich nona - arginine; N/P: nitrogen (NH3
+)/phosphate (PO4–); PCR: polymerase chain reaction; PR9: Pas
nona-arginine; RFP: red fluorescent protein; SR9: synthetic nona-arginine; Tat: transactivator of transcription

72 Min-Jie Liu, Jyh-Ching Chou and Han-Jung Lee 1. Introduction
Transgenesis is one of the key technologies for agriculture, such as crop improvement, and for pharmaceutical industry, such as plant molecular farming. Plant molecular farming is a new branch of plant biotechnologies that plants are genetically engineered to produce recombinant proteins in great demand and large quantities [1, 2]. The first higher plant was successfully transformed by chimeric bacterial genes in 1983 [3]. And the first recombinant plant-derived pharmaceutical protein, the human growth hormone, was produced in transgenic plants in 1986 [4]. After more than two decades, many exploits clearly demonstrated that plants could be turned into biofactories for large scale production of recombinant proteins, such as human serum proteins, antibodies, vaccines, hormones, and enzymes [1]. This has been possible due to the ability to perform post-translational modifications in plants that make the recombinant proteins fold properly and maintain their structural and functional integrity.
As the most common method in plant gene transformation, nuclear transformation has produced most of the recombinant proteins to date [1,2]. Stable nuclear transformation involves the incorporation of an exogenous gene into the nuclear genome of the plant. Recently, stable plastid transformation offered a valuable alternative to nuclear transformation [2, 5]. As plastids are inherited through maternal tissues in most plant species, transgenic plants with homoplastomic chloroplast transformation provided a natural containment of transgene flow by out-crossing [5]. On the other hand, the transient production platform is perhaps the fastest and the most convenient system for plant molecular farming [1, 2, 6]. These transient systems mainly used for quick validation of expression constructs are now being applied routinely for the production of recombinant proteins in great amounts within a few weeks [7]. These systems include Agrobacterium-mediated and viral vector-based methods. However, these methods have some limitations including low transgenic efficiency and cytotoxicity.
The cell membrane is a structural barrier to prevent living cells from the surrounding environment. In general, nonpolar compounds with relatively small molecular size may passively transport across the cell membrane into the cell without utilization of protein transporters. In 1988, two groups simultaneously found that the transactivator of transcription (Tat) of the human immunodeficiency virus type 1 (HIV-1) has the capability to penetrate cell membranes and enter into live cells [8, 9]. A region containing eleven basic amino acids (YGRKKRRQRRR) of the Tat protein was proven to be the functional key domain that contributes the ability to cross cell membranes [10]. Cell-penetrating peptides (CPPs), also called protein transduction domains (PTD), were then defined as short (at most 30–35 amino acid residues), water-soluble, and polybasic peptides that are able to translocate over membranes and gain access to the cell interior either alone or associated with cargoes [11]. Further, CPPs represent a major breakthrough for the transport of large-cargo molecules, such as proteins, DNAs, RNAs, peptide nucleic acids (PNAs), liposomes, and nanomaterials, into cells [11, 12]. Although CPPs have been widely applied over

Gene delivery method 73 the past two decades, the actual mechanism by which CPPs mediate the process of cellular uptake and the subsequent intracellular trafficking is still largely unresolved. Despite some common properties of CPPs, such as their cationic nature, it is generally believed that the intracellular delivery mechanism is not the same for different CPPs [11]. Currently, there are at least two major cellular uptake mechanisms, endocytosis (an energy-dependent route) and the direct membrane translocation (an energy-independent pathway), have been proposed [11–14].
Most studies of CPPs have largely focused on the applications in mammalian cells. However, only limited studies have recently explored the possible feature and applications of CPPs in plants [15]. The gradual appearance of reports of cellular internalization of diverse CPPs and their cargoes demonstrated some sort of universal characteristics of the CPPs-mediated cellular uptake in various plant cells and species. Interestingly, several exploits have reported that CPPs can be used for gene delivery in plant cells [15–19].
We previously demonstrated that a CPP, synthetic nona-arginine (SR9), is able to transport fluorescent protein or enzyme cargos into living plant cells in a covalent [20, 21] or noncovalent manner [22–24], and into other organisms including cyanobacteria, eubacteria, archaea, and yeasts noncovalently [25]. Further, SR9 could noncovalently deliver RNAs [26] or nanoparticles [27–29] into human cells. Remarkably, we also found that SR9 can deliver DNAs into plant [17] and human [30] cells. Recently, we comparatively investigated two improved arginine-rich CPPs, histidine-rich nona-arginine (HR9) and Pas nona-arginine (PR9), in the delivery of DNAs into paramecia [31] or insect cells [32], or the delivery of nanoparticles [33, 34] and proteins [35] into human cells. Encouragingly, CPPs alone or with their cargoes have no cytotoxicity at their working concentrations [23, 29, 31–33].
In this report, our major aims were to: (1) investigate the ability of three arginine-rich CPPs to deliver plasmid DNAs into plant cells at the DNA level, and (2) confirm the CPP-mediated gene delivery at the protein level. To achieve these goals, plasmid DNA was in vitro labeled and transported by CPPs into cells of mung bean (a dicot plant) and rice (a monocot plant) monitored by a fluorescent microscope and assayed by polymerase chain reaction (PCR). Finally, the functional gene assay of the CPP-mediated gene delivery method was examined using live cell imaging.
2. Materials and Methods 2.1. Plant materials, plasmid sources, and peptide preparation
Root tips of two angiosperms, mung bean (Vigna radiata) of dicotyledons and rice (Oryza sativat) of monocotyledons, were prepared as previously described [23]. The pHBTsGFP(S65T)-NOS plasmid with a GenBank accession number EF090408 contains a re-engineered GFP coding sequence under the control of the 35S cauliflower mosaic virus (CaMV) enhancer fused to the basal

74 Min-Jie Liu, Jyh-Ching Chou and Han-Jung Lee promoter of the maize C4PPDK gene [36]. Three arginine-rich CPPs, SR9 (R9 with 1423.7 Da), HR9 (C-5H-R9-5H-C with 3001.7 Da), and PR9 (FFLIPKG-R9 with 2328.9 Da) were synthesized as previously described [31–34]. 2.2. Cyanine 3-labeling of plasmid DNA
The native pBlueScript-SK+ plasmid DNA (Agilent Technologies, Santa Clara, CA, USA) was in vitro labeled with the LabelIT Cy3 nucleic acid labeling kit (Mirus Bio, Madison, WI, USA) according to the manufacturer's instructions. 2.3. The CPP-mediated DNA delivery into plant cells
The CPP-mediated DNA delivery was conducted as previously described with relevant modifications [17]. Intact root tips of mung bean and rice were washed with double distilled water twice before treatment. One µg of the CPPs was estimated to contain about 6.3 nanomolar nitrogens (NH3
+), and 1 µg of DNA to contain 3 nanomolar phosphates (PO4
–). 7.1 µg of CPPs (SR9, HR9, or PR9) was pre-mixed with 5 µg of Cy3-labeled plasmid DNA based on a molar nitrogen/phosphate (N/P) ratio of 3 to a final volume of 50 µL in a 2 ml eppendorf tube and incubated at 37°C for 1 h. Four hundred fifty µL of double distilled water was added in this tube. Root tips were then treated with CPP/DNA mixtures at 37°C for 1 h. After the incubation, roots were washed with water in order to remove free CPP/DNA and remained in planta. To reduce red autofluorescence of plants, roots were soaked in double distilled water for 30 min. Roots were then squashed onto slides and monitored without fixation using a fluorescent microscope.
For the functional gene assay, 14.2 µg of CPPs (SR9, HR9, or PR9) was pre-mixed with 10 µg of the pHBTsGFP(S65T)-NOS plasmid DNA at an N/P ratio of 3 to a final volume of 50 µL and incubated at 37°C for 1 h. Four hundred fifty µL of double distilled water was added in this eppendorf tube. Root tips were then treated with CPP/DNA mixtures at 37°C for 1 h. After the incubation, roots were washed with water. Plant roots were incubated at 37°C in dark for additional 48 h and then monitored without fixation using a fluorescent microscope. 2.4. Fluorescent microscopy
Fluorescent and bright-field images were observed by the GFP filter, red fluorescent protein (RFP) filter, and bring light using an Eclipse E600 fluorescent microscope (Nikon, Melville, NY, USA) as previously described [17]. Images were captured using a Penguin 150CL cooled CCD camera (Pixera, Los Gatos, CA, USA). 2.5. PCR analysis
The detection of PCR was performed as previously described [31]. After the treatment of root tips with the pHBTsGFP(S65T)-NOS plasmid according to the functional gene assay, root tips were treated with DNase (10 unit/µL, Promega, Madison, WI, USA) for 30 min at 37°C to remove extra-root DNAs. Lysates of the root-tip cells were PCR-amplified using the GFP5

Gene delivery method 75 (5'-AAGAATTCCATGGTGAGCAAGGGCGAGGAGCTGTT-3') and GFP6 (5'-AAGAATTCTTAAGCTTGTACAGCTCGTCCATGCCGAGAGT-3') primers. The PCR products were analyzed by electrophoresis on a 2% (wt/vol) agarose gel for 20 min, and images were captured using a Typhoon FLA 9000 biomolecular imager (GE Healthcare, Piscataway, NJ, USA). 3. Results 3.1. Delivery of the Cy3-labeled plasmid DNA by CPPs into mung bean root-tip cells
To investigate gene delivery mediated by CPPs into plant cells, the
pBlueScript-SK+ plasmid DNA lacking of any reporter gene cassette was in vitro Cyanine 3 (Cy3)-labeled. Mung bean root-tip cells were treated with the Cy3-labeled DNA alone, CPP alone, or CPP/Cy3-labeled DNA complexes (Fig. 1). We found that there was no or little signal detected in the cells treated with water (control), the Cy3-labeled DNA, or CPP using a fluorescent microscope. Little signal could be detected in plant cells due to weak red autofluorescence. However, the cells treated with SR9/Cy3-labeled DNA, HR9/Cy3-labeled DNA, or PR9/Cy3-labeled DNA complexes displayed red fluorescence using a fluorescent microscope. These results indicated that three arginine-rich CPPs (SR9, HR9, and PR9) are able to deliver the Cy3-labeled plasmid DNAs into mung bean root-tip cells. 3.2. PCR analysis of the CPP-mediated delivery of the reporter gene-containing plasmid DNA into mung bean root-tip cells
To confirm the plasmid DNA being intracellularly delivered by CPPs and expressed in plant cells, PCR analysis was performed. Mung bean root-tip cells were treated with the pHBTsGFP(S65T)-NOS plasmid DNA alone, CPPs alone, or CPP/pHBTsGFP(S65T)-NOS complexes (Fig. 2). A specific PCR product of green fluorescent protein (GFP) coding sequence with 743 bp was amplified with the GFP5 and GFP6 primers in the positive control (lane 2) and from lysates of the cells treated with SR9/DNA, HR9/DNA, or PR9/DNA complexes (lanes 9–11). These data revealed that the CPP-mediated intracellular gene delivery can be confirmed at the DNA level.

Brig
ht-f
ield
R
FP c
hann
el
Control DNA only CPP only R
FP C
hann
el
Brig
ht-f
ield
SR9/DNA HR9/DNA PR9/DNA
76 Min-Jie Liu, Jyh-Ching Chou and Han-Jung Lee Gene delivery method 77 Fig. 1. Fluorescent microscopy of the CPP-mediated delivery of the Cy3-labeled DNA into mung bean root-tip cells. Cells were treated with water as a control, the Cy3-labeled plasmid DNA alone, a CPP (HR9) alone, SR9/Cy3-labeled DNA, HR9/Cy3-labeled DNA, or PR9/Cy3-labeled DNA complexes. Images of bright-field and the RFP channel were obtained using a fluorescent microscope at a magnification of 200×.

G
FitreTaPCtewPRprPCGbi 3.D
anplwCPdeplw
Gene delivery
ig. 2. PCR eated with aipei, TaiwCR produc
emplate, reswater (Contr
R9, SR9/DNroducts werCR product
GFP5 and Giomolecular
.3. The CPNA into m
To testnd functionlasmid DNA
were treated PP/pHBTsGetected in tlasmid DNA
with SR9/DN
ry method
analysis oCPP/DNA
wan). Lanes cts with anpectively. Lrol), the pHNA, HR9/Dre separatedt of the GF
GFP6 primer imager.
PP-mediateung bean r
t whether thnally expreA was condwith the pH
GFP(S65T)the cells trA, or CPP uNA, HR9/DN
f the GFPcomplexes2 (a positi
nd without Lanes 4–11 HBTsGFP(SDNA, and d by electroFP coding rers. Images
ed deliveryroot-tip cellhe plasmid Dssed in pladucted for tHBTsGFP(S-NOS comreated with using a fluoNA, or PR9
coding seqs. Lane 1 isive control)t the pHBT
are the PCS65T)-NOSPR9/DNA ophoresis oregion is abs were capt
y of the rls DNA can bant cells, tthe functionS65T)-NOS
mplexes (Fiwater (con
orescent mic9/DNA com
quence in ms the 100 b) and 3 (a nTsGFP(S65
CR productsS plasmid D
complexes,on a 2% agbout 743 btured using
reporter ge
be intracelluthe GFP rnal assay. M plasmid DNg. 3). No ntrol), the croscope. H
mplexes exhi
mung bean p DNA ladnegative co5T)-NOS p
of the cellDNA alone, respective
garose gel. Tp amplified
g a Typhoo
ene-contain
ularly delivereporter genMung bean NA alone, Cfluorescen
pHBTsGFPHowever, the
ibited green
7
root-tip cedder (Taqkeontrol) are tplasmid DNs treated w
e, SR9, HRely. The PCThe expectd by both ton FLA 90
ning plasm
ered by CPene-containi
root-tip ceCPP alone,
nt signal wP(S65T)-NOe cells treatn fluorescen
77
ells ey, the NA ith
R9, CR ted the 00
mid
PPs ing ells
or was OS ted nce

78 Min-Jie Liu, Jyh-Ching Chou and Han-Jung Lee using a fluorescent microscope. The results demonstrated that these arginine-rich CPPs (SR9, HR9, and PR9) can deliver the plasmid DNA into mung bean root-tip cells, and the delivered DNA can be actively expressed by cells at the protein level. 3.4. Delivery of the Cy3-labeled plasmid DNA by CPPs into rice root-tip cells
To determine the CPP-mediated gene delivery in plant cells, the
pBlueScript-SK+ plasmid DNA was Cy3-labeled. Rice root-tip cells were treated with the Cy3-labeled DNA alone, CPP alone, or CPP/Cy3-labeled DNA complexes (Fig. 4). No signal could be detected in the cells treated with water (control), the Cy3-labeled DNA, or CPP using a fluorescent microscope. Remarkably, the cells treated with SR9/Cy3-labeled DNA, HR9/Cy3-labeled DNA, or PR9/Cy3-labeled DNA complexes exhibited red fluorescence. These data suggested that SR9, HR9, and PR9 are able to deliver the Cy3-labeled plasmid DNA into rice cells. 3.5. PCR analysis of the GFP coding sequence in rice root-tip cells treated with CPP/DNA complexes
Rice root-tip cells were treated with the pHBTsGFP(S65T)-NOS plasmid
DNA alone, CPPs alone, or CPP/pHBTsGFP(S65T)-NOS complexes (Fig. 5). We found that a PCR product of GFP coding region with 743 bp can be amplified only in the positive control (lane 2) and from lysates of the cells treated with SR9/DNA, HR9/DNA, or PR9/DNA complexes (lanes 9–11). These results supported the CPP-mediated gene delivery in rice cells at the DNA level. 3.6. The CPP-mediated delivery of the reporter gene-containing plasmid DNA into rice root-tip cells
To reveal the functional assay of the CPP-mediated gene delivery, rice
root-tip cells were treated with the pHBTsGFP(S65T)-NOS plasmid DNA alone, CPP alone, or CPP/pHBTsGFP(S65T)-NOS complexes (Fig. 6). No fluorescent signal was detected in the cells treated with water (control), the pHBTsGFP(S65T)-NOS plasmid DNA, or CPP using a fluorescent microscope. However, the cells treated with SR9/DNA, HR9/DNA, or PR9/DNA complexes displayed green fluorescence. These data demonstrated that SR9, HR9, and PR9 can deliver the plasmid DNA into rice cells, and the delivered DNA can be expressed by cells.
Brig
ht-f
ield
C
hann
el
A A PR9/DNA
Brig
ht-f
ield
G
FP c
hann
el
Control DNA only CPP only
Gene delivery method 79
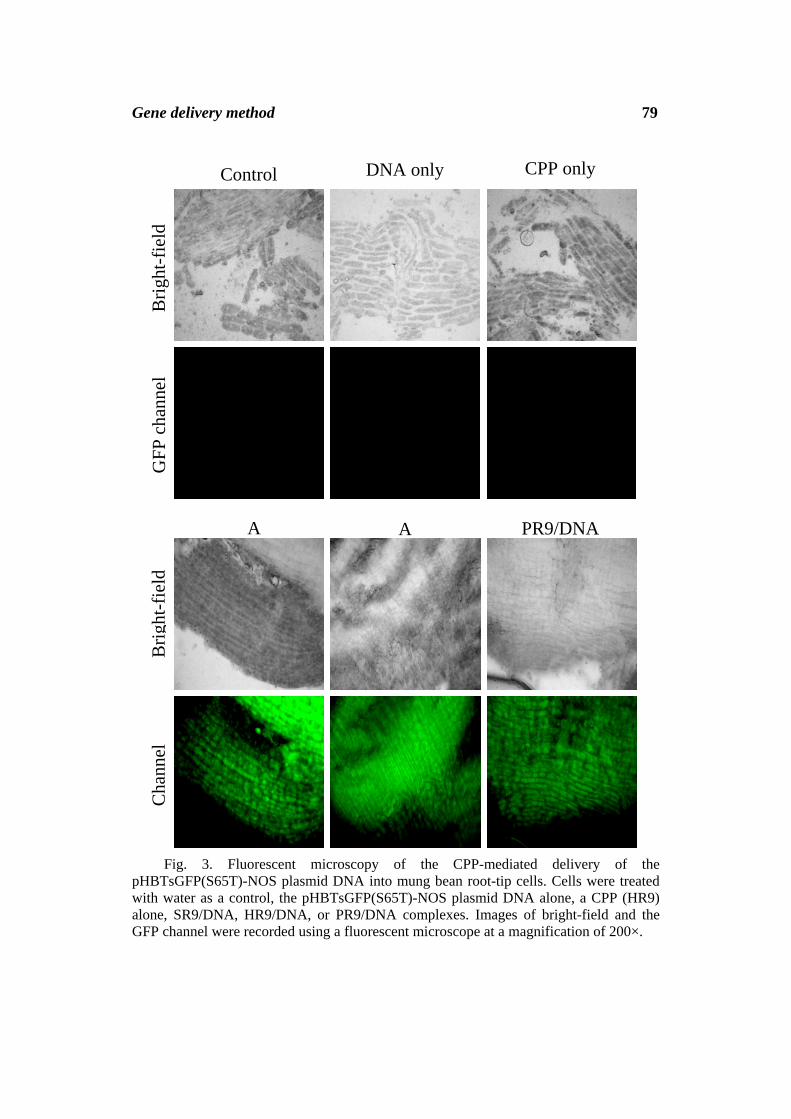
Fig. 3. Fluorescent microscopy of the CPP-mediated delivery of the pHBTsGFP(S65T)-NOS plasmid DNA into mung bean root-tip cells. Cells were treated with water as a control, the pHBTsGFP(S65T)-NOS plasmid DNA alone, a CPP (HR9) alone, SR9/DNA, HR9/DNA, or PR9/DNA complexes. Images of bright-field and the GFP channel were recorded using a fluorescent microscope at a magnification of 200×.

Brig
ht-f
ield
R
FP C
hann
el
Control DNA only CPP only
Brig
ht-f
ield
R
FP C
hann
el
SR9/DNAGFP HR9/DNASR9/DN PR9/DNAHR9/DN
80 Min-Jie Liu, Jyh-Ching Chou and Han-Jung Lee Fig. 4. Fluorescent microscopy of the CPP-mediated delivery of the Cy3-labeled DNA into rice root-tip cells. Cells were treated with water as a control, the Cy3-labeled plasmid DNA alone, a CPP (HR9) alone, SR9/Cy3-labeled DNA, HR9/Cy3-labeled DNA, or PR9/Cy3-labeled DNA complexes. Images of bright-field and the RFP channel were obtained using a fluorescent microscope at a magnification of 200×.

G
FiCPcopHPCpHHthpr
Gene delivery
ig. 5. PCR PP/DNA coontrol) and HBTsGFP(SCR produHBTsGFP(S
HR9/DNA, ahe GFP codrimers. Imag
ry method
analysis ofomplexes. 3 (a negat
S65T)-NOSucts of S65T)-NOSand PR9/DNding sequencges were ca
f GFP codiLane 1 is ttive controlS plasmid D
the cellS plasmid NA complexce is about aptured usin
ing sequencthe 100 bpl) are the P
DNA templals treated
DNA alonxes, respect743 bp amp
ng a Typhoo
ce in rice rop DNA laddPCR producate, respecti
with wne, SR9, tively. The eplified by bon FLA 900
oot-tip cellsder. Lanes cts with andively. Laneswater (CoHR9, PR9expected PCboth the GF00 biomolec
8
s treated w2 (a positi
d without ts 4–11 are tontrol), t
9, SR9/DNCR product FP5 and GFcular imager
81
ith ive the the the
NA, of P6 r.

Brig
ht-f
ield
G
FP C
hann
el
Control DNA only CPP only B
right
-fie
ld
GFP
Cha
nnel
SR9/DNA HR9/DNA PR9/DNA
82 Min-Jie Liu, Jyh-Ching Chou and Han-Jung Lee Fig. 6. Fluorescent microscopy of the CPP-mediated delivery of the pHBTsGFP(S65T)-NOS plasmid DNA into rice root-tip cells. Cells were treated with water as a control, the pHBTsGFP(S65T)-NOS plasmid DNA alone, a CPP (HR9) alone, SR9/DNA, HR9/DNA, or PR9/DNA complexes. Images of bright-field and the GFP channel were recorded using a fluorescent microscope at a magnification of 200×.

Gene delivery method 83 4. Discussion
In this report, we demonstrate that three arginine-rich CPPs (SR9, HR9, and PR9) are able to deliver plasmid DNA encoding the GFP reporter gene into mung bean and rice root-tip cells. Live cell imaging and PCR analysis revealed that the CPP-delivered DNA can be functionally expressed by plant cells at DNA and protein levels. Our results indicate that these arginine-rich CPPs are excellent nanocarriers for gene delivery in living plant cells. Studies of the CPP-mediated gene delivery may have great potential for the research of plant gene transformation and may open a new avenue for plant molecular farming in future.
Plasma membranes of plant or animal cells play important roles in the maintenance of cellular homeostasis as well as the control of molecular trafficking with the ambient environment. Cell walls which represent one of fundamental differences between plant and animal cells are composed of complicated network of various carbohydrates. In 2005, we first demonstrated that CPPs are able to deliver bioactive proteins into intact, live plant cells [17, 20]. This highlighted that cell walls are not a barrier for the characteristic internalization of CPPs into plant cells at all. Later, Mae et al. reported that CPPs can enter into plant protoplasts, which had the plant cell walls removed [37]. More detailed studies between CPPs and plasma membrane/cell wall in plants were subsequently reported [38, 39]. Collectively, our results presented here agreed well with previous data that CPPs are able to permeate through cell walls and membranes of plants. In this study, HR9 (C-5H-R9-5H-C) was designed to have a SR9 sequence flanked by polyhistidine and cysteine residues at both N- and C-terminals, and PR9 (FFLIPKG-R9) to possess a penetration accelerating sequence Pas at the N-terminal of SR9. Both HR9 and PR9 represent two improved versions of CPPs over SR9 for gene delivery in plants. Mizuno et al. found that the internalization of arginine-rich CPPs into intact tobacco cells with negative charge-rich cell walls depends on the number of arginine residues in CPPs [38]. Additionally, the position of small cargoes attached to arginine-rich CPPs markedly affected the internalization efficiency. Recently, it was proposed that the association with the cytoskeleton (actin filaments and microtubules) is essential to the cellular uptake of a CPP into living tobacco cells [39, 40].
It has been known that CPPs represent a novel tool for the transport of plasmid DNA into mammalian cells [41–45]. However, in 2005, Roberts commented for the first time that both nucleic acids and proteins can be potentially delivered into plant cells by a simple application of CPPs [16]. Until 2007, we provided the first complete evidence for the CPP-mediated plasmid DNA delivery system in plant cells [17]. SR9 was clearly demonstrated to be a nuclear routing carrier that can transfer plasmid DNA not only through cell wall/membrane but also into nuclei of plant cells for gene expression. Accordingly, our results of Figures 3 and 6 presented here are consistent with previous data obtained with SR9 and DNA in plants [17]. Interestingly, we found that the differential DNA-binding affinities among three arginine-rich CPPs are correlated well with the efficiencies of gene delivery with the order of HR9 > PR9

84 Min-Jie Liu, Jyh-Ching Chou and Han-Jung Lee > SR9 [31–33]. Nakase et al. reviewed that the actual mechanisms of CPP-mediated cellular uptake essentially influence the efficiencies and biological activities of delivered cargoes in cells [13]. Therefore, we concluded that both the differential CPP-DNA-binding affinities in vitro and the vary mechanisms of cellular uptake of CPP/DNA are two limiting factors resulting in different gene delivery efficiencies of these arginine-rich CPPs [32]. Our mechanistic studies indicated that the cellular entry for HR9/nanoparticle complexes is mediated by the direct membrane translocation [33], while the cellular uptake for SR9/protein complexes is mediated by multiple internalization pathways [46].
Most CPPs have been shown to be nontoxic at their working concentrations and do not interfere with the functionality of the delivered cargoes in plant and animal cells [15, 18, 19]. We previously demonstrated that CPPs alone or with their cargoes have no cytotoxicity in plant cells by either Trypan blue assay [17, 20, 23] or 1-(4,5-dimethylthiazol-2-yl)-3,5-diphenylformazan (MTT) assay [17]. Further, the in vitro safety of most CPPs has also been confirmed in a more detailed metabolomic analysis in Chinese hamster ovary (CHO) cells [47]. Recently, Suhorutsenko et al. evaluated the influence of CPPs and CPP/nucleic acid complexes for the undesired toxicity and immunogenicity both in vitro and in vivo [48]. They concluded that all CPPs are nontoxic and nonimmunogenic in human cell lines and in mice. Together, these findings indicated that CPPs exhibit promising potential as safe and efficient nanocarriers in the delivery of cargo molecules into live cells without risks of toxicity and inflammatory reactions. 5. Conclusion
We performed an improved method that three arginine-rich CPPs (SR9, HR9, and PR9) are capable of delivering plasmid DNA into dicot and monocot plant cells. The CPP-delivered plasmid encoding the GFP reporter gene could be successfully expressed by cells. These studies may indicate the significant utility of the improved CPP-mediated gene delivery for the research of transgenic plants in future. Acknowledgements
We thank Dr. Jen Sheen (Harvard University, MA, USA) for provision of the pHBTsGFP(S65T)-NOS plasmid. This work was supported by the Grant Number NSC 101-2320-B-259-002-MY3 from the National Science Council of Taiwan. References [1] O.O. Obembe, J.O. Popoola, S. Leelavathi and S.V. Reddy, Advances in
plant molecular farming, Biotechnology Advances, 29 (2011), 210–222.

Gene delivery method 85
[2] M.E. Horn, S.L. Woodard and J.A. Howard, Plant molecular farming:
systems and products, Plant Cell Reports, 22 (2004), 711–720. [3] R.T. Fraley, S.G. Rogers, R.B. Horsch, P.R. Sanders, J.S. Flick, S.P. Adams,
M.L. Bittner, L.A. Brand, C.L. Fink, J.S. Fry, G.R. Galluppi, S.B. Goldberg, N.L. Hoffman and S.C. Woo, Expression of bacterial genes in plant cells, Proceedings of the National Academy of Sciences of the United States of America, 80 (1983), 4803–4807.
[4] A. Barta, K. Sommengruber, D. Thompson, K. Hartmuth, M.A. Matzke and A.J.M. Matzke, The expression of a nopaline synthase – human growth hormone chimaeric gene in transformed tobacco and sunflower callus tissue, Plant Molecular Biology, 6 (1986), 347–357.
[5] B. Meyers, A. Zaltsman, B. Lacroix, S.V. Kozlovsky and A. Krichevsky, Nuclear and plastid genetic engineering of plants: comparison of opportunities and challenges, Biotechnology Advances, 28 (2010), 747–756.
[6] E.P. Rybicki, Plant-made vaccines for humans and animals, Plant Biotechnology Journal, 8 (2010), 620–637.
[7] L.P. Vezina, L. Faye, P. Lerouge, M.A. D'Aoust, E. Marquet-Blouin, C. Burel, P.O. Lavoie, M. Bardor and V. Gomord, Transient co-expression for fast and high-yield production of antibodies with human-like N-glycans in plants, Plant Biotechnology Journal, 7 (2009), 442–455.
[8] M. Green and P.M. Loewenstein, Autonomous functional domains of chemically synthesized human immunodeficiency virus Tat trans-activator protein, Cell, 55 (1988), 1179–1188.
[9] A.D. Frankel and C.O. Pabo, Cellular uptake of the Tat protein from human immunodeficiency virus, Cell, 55 (1988), 1989–1993.
[10] E. Vives, P. Brodin and B. Lebleu, A truncated HIV-1 Tat protein basic domain rapidly translocates through the plasma membrane and accumulates in the cell nucleus, Journal of Biological Chemistry, 272 (1997), 16010–16017.
[11] F. Madani, S. Lindberg, U. Langel, S. Futaki and A. Graslund, Mechanisms of cellular uptake of cell-penetrating peptides, Journal of Biophysics, 2011 (2011), 414729.
[12] J.M. Gump and S.F. Dowdy, TAT transduction: the molecular mechanism and the therapeutic prospects, Trends in Molecular. Medicine, 13 (2007), 443–448.
[13] I. Nakase, S. Kobayashi and S. Futaki, Endosome-disruptive peptides for improving cytosolic delivery of bioactive macromolecules, Biopolymers, 94 (2010), 763–770.
[14] N. Schmidt, A. Mishra, G.H. Lai and G.C. Wong, Arginine-rich cell-penetrating peptides, FEBS Letters, 584 (2010), 1806–1813.

86 Min-Jie Liu, Jyh-Ching Chou and Han-Jung Lee
[15] A. Chugh, F. Eudes and Y.S. Shim, Cell-penetrating peptides: nanocarrier for
macromolecule delivery in living cells, IUBMB Life, 62 (2010), 183–193. [16] M.R. Roberts, Fast-track applications: the potential for direct delivery of
proteins and nucleic acids to plant cells for the discovery of gene function, Plant Methods, 1 (2005), 12.
[17] C.P. Chen, J.C. Chou, B.R. Liu, M. Chang and H.J. Lee, Transfection and expression of plasmid DNA in plant cells by an arginine-rich intracellular delivery peptide without protoplast preparation, FEBS Letters, 581 (2007), 1891–1897.
[18] A. Chugh and F. Eudes, Study of uptake of cell penetrating peptides and their cargoes in permeabilized wheat immature embryos, FEBS Journal, 275 (2008), 2403–2414.
[19] A. Chugh, E. Amundsen and F. Eudes, Translocation of cell-penetrating peptides and delivery of their cargoes in triticale microspores, Plant Cell Reports, 28 (2009), 801–810.
[20] M. Chang, J.C. Chou and H.J. Lee, Cellular internalization of fluorescent proteins via arginine-rich intracellular delivery peptide in plant cells, Plant and Cell Physiology, 46 (2005), 482–488.
[21] K. Liu, H.J. Lee, S.S. Leong, C.L. Liu and J.C. Chou, A bacterial indole-3-acetyl-L-aspartic acid hydrolase inhibits mung bean (Vigna radiata L.) seed germination through arginine-rich intracellular delivery, Journal of Plant Growth and Regulation, 26 (2007), 278–284.
[22] Y.H. Wang, C.P. Chen, M.H. Chan, M. Chang, Y.W. Hou, H.H. Chen, H.R. Hsu, K. Liu and H.J. Lee, Arginine-rich intracellular delivery peptides noncovalently transport protein into living cells, Biochemical and Biophysical Research Communications, 346 (2006), 758–767.
[23] M. Chang, J.C. Chou, C.P. Chen, B.R. Liu and H.J. Lee, Noncovalent protein transduction in plant cells by macropinocytosis, New Phytologist, 174 (2007), 46–56.
[24] S.W. Lu, J.W. Hu, B.R. Liu, C.Y. Lee, J.F. Li, J.C. Chou and H.J. Lee, Arginine-rich intracellular delivery peptides synchronously deliver covalently and noncovalently linked proteins into plant cells, Journal of Agricultural and Food Chemistry, 58 (2010), 2288–2294.
[25] B.R. Liu, J.C. Chou and H.J. Lee, Cell membrane diversity in noncovalent protein transduction, Journal of Membrane Biology, 222 (2008), 1–15.
[26] Y.H. Wang, Y.W. Hou and H.J. Lee, An intracellular delivery method for siRNA by an arginine-rich peptide, Journal of Biochemical and Biophysical Methods, 70 (2007), 579–586.

Gene delivery method 87
[27] B.R. Liu, J.F. Li, S.W. Lu, H.J. Lee, Y.W. Huang, K.B. Shannon and R.S.
Aronstam, Cellular internalization of quantum dots noncovalently conjugated with arginine-rich cell-penetrating peptides, Journal of Nanoscience and Nanotechnology, 10 (2010), 6534–6543.
[28] B.R. Liu, Y.W. Huang, H.J. Chiang and H.J. Lee, Cell-penetrating peptide-functionized quantum dots for intracellular delivery, Journal of Nanoscience and Nanotechnology, 10 (2010), 7897–7905.
[29] Y. Xu, B.R. Liu, H.J. Lee, K.S. Shannon, J.G. Winiarz, T.C. Wang, H.J. Chiang and Y.W. Huang, Nona-arginine facilitates delivery of quantum dots into cells via multiple pathways, Journal of Biomedicine and Biotechnology, 2010 (2010), 948543.
[30] C.Y. Lee, J.F. Li, J.S. Liou, Y.C. Charng, Y.W. Huang and H.J. Lee, A gene delivery system for human cells mediated by both a cell-penetrating peptide and a piggyBac transposase, Biomaterials, 32 (2011), 6264–6276.
[31] Y.H. Dai, B.R. Liu, H.J. Chiang and H.J. Lee, Gene transport and expression by arginine-rich cell-penetrating peptides in Paramecium, Gene 489 (2011), 89–97.
[32] Y.J. Chen, B.R. Liu, Y.H. Dai, C.Y. Lee, M.H. Chan, H.H. Chen, H.J. Chiang and H.J. Lee, A gene delivery system for insect cells mediated by arginine-rich cell-penetrating peptides, Gene 493 (2012), 201–210.
[33] B.R. Liu, Y.W. Huang, J.G. Winiarz, H.J. Chiang and H.J. Lee, Intracellular delivery of quantum dots mediated by a histidine- and arginine-rich HR9 cell-penetrating peptide through the direct membrane translocation mechanism, Biomaterials, 32 (2011), 3520–3537.
[34] B.R. Liu, Y.W. Huang, H.J. Chiang and H.J. Lee, Primary effectors in the mechanisms of transmembrane delivery of arginine-rich cell-penetrating peptides, Advanced Studies in Biology, 5 (2013), 11–25.
[35] J.S. Liou, B.R. Liu, A.L. Martin, Y.W. Huang, H.J. Chiang and H.J. Lee, Protein transduction in human cells is enhanced by cell-penetrating peptides fused with an endosomolytic HA2 sequence, Peptides, 37 (2012), 273–284.
[36] W. Chiu, Y. Niwa, W. Zeng, T. Hirano, H. Kobayashi and J. Sheen, Engineered GFP as a vital reporter in plants, Current Biology, 6 (1996), 325–330.
[37] M. Mae, H. Myrberg, Y. Jiang, H. Paves, A. Valkna and U. Langel, Internalization of cell-penetrating peptides into tobacco protoplasts, Biochimica et Biophysica Acta, 1669 (2005), 101–107.
[38] T. Mizuno, M. Miyashita and H. Miyagawa, Cellular internalization of arginine-rich peptides into tobacco suspension cells: a structure-activity relationship study, Journal of Peptide Science, 15 (2009), 259–263.

88 Min-Jie Liu, Jyh-Ching Chou and Han-Jung Lee
[39] K. Eggenberger, C. Mink, P. Wadhwani, A. S.Ulrich and P. Nick, Using the
peptide Bp100 as a cell-penetrating tool for the chemical engineering of actin filaments within living plant cells, ChemBioChem, 12 (2011), 132–137.
[40] K. Eggenberger, E. Birtalan, T. Schroder, S. Brase and P. Nick, Passage of Trojan peptides into plant cells, ChemBioChem, 10 (2009), 2504–2512.
[41] M. Lindgren, M. Hallbrink, A. Prochiantz and U. Langel, Cell-penetrating peptides, Trends in Pharmacological Sciences, 21 (2000), 99–103.
[42] K. Rittner, A. Benavente, A. Bompard-Sorlet, F. Heitz, G. Divita, R. Brasseur and E. Jacobs, New basic membrane-destabilizing peptides for plasmid-based gene delivery in vitro and in vivo, Molecular Therapy, 5 (2002), 104–114.
[43] E.L. Synder and S.F. Dowdy, Recent advances in the use of protein transduction domains for the delivery of peptides, proteins and nucleic acids in vivo, Expert Opinion on Drug Delivery, 2 (2005), 43–51.
[44] Z. Liu, M. Li, D. Cui and J. Fei, Macro-branched cell-penetrating peptide design for gene delivery, Journal of Controlled Release, 102 (2005), 699–710.
[45] H.S. Choi, H.H. Kim, J.M. Yang and S. Shin, An insight into the gene delivery mechanism of the arginine peptide system: role of the peptide/DNA complex size, Biochimica et Biophysica Acta, 1760 (2006), 1604–1612.
[46] J.W. Hu, B.R. Liu, C.Y. Wu, S.W. Lu and H.J. Lee, Protein transport in human cells mediated by covalently and noncovalently conjugated arginine-rich intracellular delivery peptides, Peptides, 30 (2009), 1669–1678.
[47] K. Kilk, R. Mahlapuu, U. Soomets and U. Langel, Analysis of in vitro toxicity of five cell-penetrating peptides by metabolic profiling, Toxicology, 265 (2009), 87–95.
[48] J. Suhorutsenko, N. Oskolkov, P. Arukuusk, K. Kurrikoff, E. Eriste, D.M. Copolovici and U. Langel, Cell-penetrating peptides, PepFects, show no evidence of toxicity and immunogenicity in vitro and in vivo, Bioconjugate Chemistry, 22 (2011), 2255–2262.
Received: November, 2012


















