
A dissertation submitted in partial satisfaction of the...
161
Core Promoter Recognition Complex Switching in Liver Development and Regeneration by Joseph Anthony D’Alessio A.B. (Bowdoin College) 1998 A dissertation submitted in partial satisfaction of the requirements for the degree of Doctor of Philosophy in Molecular and Cell Biology in the Graduate Division of the University of California, Berkeley Committee in charge: Professor Robert Tjian, Chair Professor Michael Levine Professor Iswar Hariharan Professor David V. Schaffer Fall 2009
Transcript of A dissertation submitted in partial satisfaction of the...

Core Promoter Recognition Complex Switching in Liver Development and Regeneration
by
Joseph Anthony D’Alessio A.B. (Bowdoin College) 1998
A dissertation submitted in partial satisfaction
of the requirements for the degree of
Doctor of Philosophy
in
Molecular and Cell Biology
in the
Graduate Division
of the
University of California, Berkeley
Committee in charge:
Professor Robert Tjian, Chair Professor Michael Levine Professor Iswar Hariharan
Professor David V. Schaffer
Fall 2009


Abstract
Core Promoter Recognition Complex Switching
in Liver Development and Regeneration
by
Joseph Anthony D’Alessio
Doctor of Philosophy in Molecular and Cell Biology
University of California, Berkeley
Professor Robert Tjian, Chair
The exquisite structural and functional complexity of the mammalian body
requires the exactingly precise spatial and temporal regulation of gene expression. The
most critically regulated step of which is the transcription of DNA to RNA by the
elaborate interplay of sequence-specific activators and repressors, chromatin modifiers,
coactivators, the basal machinery and RNA polymerase II. Until recently, developmental
stage and tissue-specific gene expression patterns were believed to be regulated primarily
by the combinatorial actions of DNA binding activators and repressors, while the core
machinery was held to be invariant. New evidence suggests that significant differences
in coactivator and core promoter recognition complex composition between cell types
and developmental time points may facilitate the global changes in gene expression
programs necessary to confer functional diversity. Because of its critical role in
vertebrate development and homeostasis, the role of coactivator and core promoter
recognition complex switching in the developing liver was examined.

This work shows that mouse liver progenitors or hepatoblasts contain significant
levels of the canonical core promoter recognition complex TFIID, including the TATA
binding protein and multiple associated factors or TAFs, and the canonical coactivator
complex Mediator. These complexes and their constitutive proteins are significantly
downregulated in adult hepatocytes. Furthermore, the promoters of several TFIID
components become enriched for repressive chromatin marks in adult hepatocytes, and an
in vitro model of liver development recapitulates the downregulation of these complexes
observed in vivo. In contrast, expression of the TBP-related factor TRF3, the TAF7
paralogue TAF7l, and TAF13 is maintained or induced upon hepatic differentiation.
Additionally, these proteins associate with high molecular weight complexes in vitro
where they are enriched at hepatocyte-specific promoters, and their depletion attenuates
hepatic gene induction. Together these observations support a model wherein liver
development and the induction of hepatic gene expression requires the down regulation
of TFIID and its replacement with one or more complexes likely containing TRF3,
TAF7l and TAF13.

For my parents
Anne and Richard D’Alessio
who have supported all my adventures
and
with appreciation for Laurie McDonough
my best friend

Table of Contents
Chapter 1: Introduction
Summary 1
TFIID, TAFs, and the Basal Transcription Machinery 3
Alternative TAF Complexes, TBP-Related Factors, 7 and Tissue-Specific TAFs
TAFs in Metazoan Development and Differentiation 11
Embryonic and Postnatal Liver Development 14
Hepatic Regeneration 18
Embryonic Stem Cells and Therapeutic Approaches 22
Figures 26
References 32
Chapter 2: TBP, TRFs, and TAFs in Liver Development and Regeneration
Summary 54
Introduction 55
Results 58
Discussion 68
Materials and Methods 71
Figures 76
References 94
Chapter 3: TRF3, TAF7l, and TAF13 in Hepatic Gene Regulation
Summary 99
Introduction 100

Results 110
Discussion 119
Materials and Methods 124
Figures 129
References 145

Chapter 1: Introduction 1
Chapter 1: Introduction
Summary
The precise spatial and temporal regulation of gene expression essential to
metazoan development and cellular homeostasis requires a complex interplay between
sequence-specific activators and repressors, coactivators, chromatin modifiers, general
transcription factors (GTFs), and RNA polymerase II (PolII). The first and most
regulated step in this process is the exacting recognition of unique gene-specific DNA
regulatory sequences by diverse activator proteins. These activators then recruit the core
promoter recognition complex TFIID consisting of the TATA box binding protein (TBP)
and 12-15 associated factors (TAFs) to the site of transcription. Subsequently,
coactivators including the Mediator complex (Med), chromatin modifiers, and the
additional GTFS TFIIA-F are recruited to the core promoter to form the preinitiation
complex (PIC). This allows for the recruitment of PolII and the start of productive RNA
transcription.
Historically, developmental stage and tissue-specific patterns of gene expression
were thought to be determined primarily by DNA regulatory sequences and their
associated activators, while the general transcription machinery including TFIID was
held to be invariant (reviewed by Thomas and Chiang 2006). However, recent studies
have shown that during skeletal muscle myogenesis a novel mechanism of regulating
global patterns of gene expression by switching of core promoter recognition complexes
is employed (reviewed by Reina and Hernandez 2007). Whether this novel mechanism

Chapter 1: Introduction 2
of transcriptional regulation exists in other tissues and developmental programs or is
restricted to muscle is currently unknown.
Chapter two examines patterns of TBP and TAF expression in diverse adult
mouse tissues and finds that they are significantly down regulated in the adult liver.
Further by purifying fetal liver progenitors (hepatoblasts), partially differentiated
neonatal hepatocytes, adult hepatocytes, and cycling regenerative hepatocytes, I
demonstrate that TFIID is significantly upregulated in liver progenitors and entirely
absent from the fully differentiated cell type, while at intermediate levels in slowly
dividing or not fully differentiated populations. In striking contrast, the TBP-related
factor TRF3 and TAF13 are expressed at the same levels throughout liver development
and the unique TAF paralogoue TAF7l is highly induced upon differentiation.
Importantly, these changes occur at both the transcriptional and protein levels and can be
reconstituted in an in vitro cellular model of liver differentiation.
The third chapter of this thesis confirms the presence of TRF3, TAF7l, and
TAF13 polypeptides in differentiated liver cells and provides evidence that they are
components of one or more high molecular weight complexes in vivo. Furthermore,
RNAi mediated depletion of each of these three proteins attenuates hepatic gene
induction in an in vitro model of liver differentiation, and all three proteins show varying
degrees of enrichment at the proximal promoters of hepatocyte-specific genes. Finally,
an embryonic stem cell based hepatic differentiation protocol suggests that these changes
in core promoter recognition complex architecture may extend beyond the murine model.

Chapter 1: Introduction 3
TFIID, TAFs, and the Basal Transcription Machinery
Proteins, the basic functional units of every living cell, are produced by the
transcription of DNA to messenger RNA, the splicing and nuclear export of this mRNA,
and the translation of mRNAs to polypeptides in a cell’s cytoplasm; a process known as
the Central Dogma (Crick 1958). The most critical and regulated step in this process is
the precise transcription of protein coding regions within a cell’s DNA by RNA
polymerase II (PolII) to form mRNA. The recruitment of PolII and initiation of
transcription at specific genes requires the exquisite coordination of hundreds of
polypeptides including the core promoter recognition complex TFIID (Figure 1) (Thomas
and Chiang 2006).
TFIID was initially identified as a biochemical activity required for in vitro
transcription by PolII (Matsui et al. 1980). Upon further purification, it was identified as
a 750kD complex containing the TATA box binding protein (TBP) and 12-15 associated
factors (TAFs) that are required for activated transcription both in vitro and in vivo
(Dynlacht et al. 1991; Pugh and Tjian 1991). It is now known that both TBP and most of
the TAFs are highly conserved in yeast (S. cerevisiae), worms (C. elegans), flies (D.
melanogaster) and humans (H. sapiens) and partially conserved in the metazoan
predecessor M. brevicollis, suggesting an ancient evolutionary origin (Burley and Roeder
1996; Albright and Tjian 2000; King et al. 2008).
The process of regulated transcription takes place by the stepwise assembly of the
preinitiation complex (PIC), followed by promoter escape, productive elongation, and
termination. PIC assembly is initiated through the precise recognition and binding of

Chapter 1: Introduction 4
distinct DNA sequences or enhancers within a gene’s distal or proximal regulatory
regions by a diverse set of tissue and developmental stage-specific transcriptional
activators. These activators in turn position TFIID at the core promoter through binding
of their activation domains either directly to TBP or a subset of specific TAFs (Stringer et
al. 1990; Chen et al. 1994; Thut et al. 1995). TFIID’s affinity for the core promoters is
further stabilized by its recognition of conserved motifs within the promoter itself
through both TBP and a limited number of TAFs (Smale and Kadonaga 2003). The most
widespread and important of these motifs include the TATA box (-30), the initiator (Inr,
+1), and the downstream promoter element (DPE, +30), while unique core promoter
elements continue to be discovered and their significance analyzed (Juven-Gershon et al.
2008). Currently, it is poorly understood whether distinct core promoter recognition
complexes are selectively recruited to specific genes based on their preference for certain
core promoter motifs (Muller et al. 2007).
TFIID’s association with promoter DNA is further stabilized through the
sequential recruitment of heterodimeric TFIIA, which can bind directly to TBP, TAFs
and activators, and the single subunit TFIIB, which interacts with TFIID, PolII and
promoter DNA itself (Maldonado et al. 1990). As part of this heterotrimeric complex,
TFIIB in turn recruits TFIIF, which maintains multiple contacts with, and thus brings to
the promoter, the PolII holoenzyme (Ha et al. 1993). Subsequently, TFIIF recruits
heterodimeric TFIIE, which stabilizes PolII binding and primarily functions to recruit the
large multisubunit TFIIH, which posses multiple enzymatic activities required for the
initiation of transcription (Maxon et al. 1994). Hence, TFIIH contains a helicase activity

Chapter 1: Introduction 5
necessary for the melting of promoter DNA, an ATPase activity necessary for formation
of the first phosphodiester bond, and a kinase domain which phosphorylates the C-
terminus of PolII to signal the transition from initiation to productive elongation (Lu et al.
1992; Holstege et al. 1996). Thus, once recruited to the core promoter by sequence-
specific activators, TFIID coordinates the stepwise assembly of all general transcription
factors required for initiation of productive PolII transcription.
Extensive structural and mapping studies have begun to provide a picture of how
TBP and the TAFs are assembled and positioned within the larger TFIID holoenzyme
(Cler et al. 2009). One of the first insights into TFIID structure came from the
identification of histone fold domains within several of the TAFs, leading to speculation
that TFIID may contain nucleosome like octomers. Biochemical studies have confirmed
that TAFs with like histone fold domains do pair to form heterodimers that make up the
basic structural subunits of the complex, including the pairs TAF4-TAF12, TAF6-TAF9,
TAF8-TAF10, and TAF11-TAF13 (Selleck et al. 2001; Leurent et al. 2002). Though the
exact stoichiometry of TAFs within the complex is unclear, biochemical studies suggest
that certain TAFs, including TAF1, TAF2, and TAF7, are present as a single copy while
the majority of TAFs are present as two or more subunits per TFIID molecule (Sanders et
al. 2002). Though X-ray crystallographic structures of intact TFIID have not yet been
solved, lower resolution electron microscopy and particle reconstruction has provided
some insight to the three dimensional structure of the complex (Andel et al. 1999; Brand
et al. 1999; Sanders et al. 2002). These structures reveal a tri-lobed horseshoe like
structure in which TBP can be localized to the central cleft, suggesting that this is the

Chapter 1: Introduction 6
sight of DNA binding. Further, TAF12, TAF4, TAF6, and TAF9 appear to be present in
each lobe, while TAF5 is present in the linker region joining all three lobes consistent
with a scaffolding function (Leurent et al. 2004). RNAi and overexpression studies in
insect cells suggest that TAF4, TAF5, TAF6, TAF9, and TAF12 are essential for the
formation of a stable core complex in vivo, which is then decorated by additional TAFs to
form holo-TFIID (Leurent et al. 2002; Wright et al. 2006).
Through a combination of in vitro and in vivo experiments, the specific functions
of individual TAFs have been greatly illuminated (Albright and Tjian 2000). A subset of
TAFs has been found to bind known sequence-specific activators and thus link these
proteins’ activation domains to the rest of the basal machinery. Some of these observed
interactions include binding of p53 to TAF6 and TAF9, binding of SP1 and Pygopus to
TAF4, binding of VP16 to TAF9, and binding of multiple activators to TAF7 (Goodrich
et al. 1993; Weinzierl et al. 1993a; Chiang and Roeder 1995; Thut et al. 1995; Wright and
Tjian 2009). Yet other TAFs bind specific motifs within the core promoter to help
stabilize TFIID position and PIC assembly. Hence, TAF1 and TAF2 can bind the Inr
element while TAF6 and TAF9 contact the DPE (Verrijzer et al. 1995; Burke and
Kadonaga 1997). More recently, some TAFs have been shown to interact directly with
chromatin marks such as TAF1 with acetylated histone tails and TAF3 with trimethylated
histone H3 lysine 4 residues (Jacobson et al. 2000; Vermeulen et al. 2007). Interestingly,
TAF1 possesses kinase, ubiquitin ligase, and acetylase enzymatic activities which may
act on histones or activators to further modulate transcription (Wassarman and Sauer

Chapter 1: Introduction 7
2001). Thus, individual TAFs serve specific functions within the context of the larger
complex, many of which are yet to be fully understood.
Alternative TAF Complexes, TBP-Related Factors, and Tissue-Specific TAFs
While TFIID is thought to be ubiquitously expressed and present at most genes,
several complexes have been discovered which contain a subset of TAFs, are free of
TBP, and likely act on a more limited number of promoters (Figure 2). The human TBP-
free TAF-containing complex (TFTC) which contains many of the TAFs as well as
additional proteins not found in TFIID, is able to bind multiple activators and facilitate in
vitro transcription, though its role in vivo is not well understood (Wieczorek et al. 1998).
The yeast Spt-Ada-GCN5 acetyltransferase complex (SAGA) contains both overlapping
and unique subunits with TFTC and appears to be important in the activation and
recruitment of free TBP to a subset of mostly stress responsive genes (Grant et al. 1998;
Huisinga and Pugh 2004). Similarly, SAGA-like (SLIK) and the human Spt3-TAF9-
GCN5L acetyltransferase complex (STAGA) have very similar composition to SAGA
and likely many of the same functions but are less well studied (Martinez et al. 1998;
Pray-Grant et al. 2002). The p300/CBP-associated factor (PCAF) complex has many
similarities to yeast SAGA and is involved in both global histone modifications and
locus-specific coactivation; additionally its disruption has been implicated in oncogenic
transformation (Ogryzko et al. 1998; Nagy and Tora 2007). Hence, while the
physiological roles of these alternative TAF containing complexes are not yet fully

Chapter 1: Introduction 8
understood, it is clear that TAFs have important functions outside the context of
canonical TFIID.
To date three distinct TBP-related factors (TRFs) or TBP-like proteins (TBPL)
have been discovered, from worms to humans, and their biological significance is
beginning to be unraveled (Figure 2) (Aoyagi and Wassarman 2000; Hochheimer and
Tjian 2003). TBP-related factor 1 (TRF1) was the first of these proteins to be identified
and appears to be unique to Drosophila, where it is highly expressed in the nervous
system and testes (Crowley et al. 1993). Similarly to TBP, TRF1 is found in a large
complex, binds TFIIA, TFIIB, and TATA containing DNA, and can promote PolII
transcription in vitro (Hansen et al. 1997). However, TRF1 appears to be distinct from
TBP in that its expression is restricted to a limited number of tissues, it is present at
relatively few promoters, and it also binds to a unique TC box core promoter element
(Holmes and Tjian 2000). Initially identified based on homology, TBP-related factor 2
(TRF2) is found from worms to humans and is more closely related to TBP than TRF1
(Dantonel et al. 1999; Rabenstein et al. 1999). Though it does interact with TFIIA and
TFIIB, TRF2 fails to bind a canonical TATA box and appears to bind novel promoters
not occupied by TBP or TRF1 (Hansen et al. 1997). Hence, it has been suggested that
TRF2 may also act as a repressor by disrupting PIC formation at TATA containing
promoters (Moore et al. 1999). Further, TRF2 is part of a high molecular weight
complex which lacks TAFs but contains known chromatin remodeling factors
(Hochheimer et al. 2002). Knockdown studies in worms and frogs (X. laevis) showed
that loss of TRF2 caused early embryonic arrest through the disruption of developmental

Chapter 1: Introduction 9
gene expression (Dantonel et al. 2000; Veenstra et al. 2000); however, TRF2 knockout
mice are viable with the only apparent phenotype being disrupted spermiogenesis (Zhang
et al. 2001).
The most recent and perhaps most interesting of these proteins to be identified is
TBP-related factor 3 (TRF3/TBPL2). Initially isolated based on the near identity of its
C-terminus to that of TBP including the DNA and GTF binding domains, TRF3 is
conserved in vertebrates from fish (Fugu) to humans but is absent in lower metazoans
(Persengiev et al. 2003). Initial reports showed that it was widely but variably expressed
in adult mouse and human tissues, migrated as part of a larger 150-200 kD complex, and
that its nuclear import was regulated during the cell cycle asynchronously from that of
TBP. Subsequent studies showed TRF3 to be highly enriched in the ovary and to a lesser
extent in the testes where it had a different pattern of nuclear import than TBP. However,
these reports conflicted as to whether TRF3 is expressed in early embryogenesis
following fertilization (Xiao et al. 2006; Yang et al. 2006; Gazdag et al. 2007; Jacobi et
al. 2007). Additionally, one of these studies found that TRF3 and TBP occupied unique
sets of promoters in mouse embryonic stem cells, while two others showed that TRF3 but
not TBP was required for the transcription of a majority of embryonic genes.
Knockdown studies in frogs and zebrafish (D. rerio) determined that TRF3 is required for
embryogenesis and may be able to partially rescue loss of TBP in the early embryo
(Bartfai et al. 2004; Jallow et al. 2004). Interestingly, regulation of the mespa gene by
TRF3 is shown to be the earliest and an essential step in the commitment of mesoderm to
hematopoietic precursors, suggesting that TRF3 plays a role in cell-type-specific

Chapter 1: Introduction 10
differentiation programs (Hart et al. 2007). Perhaps the best evidence for TRF3’s role in
tissue-specific differentiation comes from studies of the transition from muscle precursors
to mature muscle fibers. In this myogenic program it was found that canonical TFIID,
including both TBP and most of the TAFs, is completely degraded, while a unique
complex of TRF3 and TAF3 is retained in the adult cell type (Deato and Tjian 2007).
Furthermore, knockdown of TRF3 blocks myogenic differentiation and TRF3 but not
TBP is enriched at the promoters of key myogenic genes. Additional in vitro dissection
of this mechanism found that the TRF3/TAF3 complex cooperates with the myogenic
activator MyoD to initiate transcription of the myogenin promoter, and that this complex
is necessary and sufficient for the process (Deato et al. 2008).
In addition to TBP-related factors, a series of novel tissue-specific TAF paralogs
have been identified which further expand the diversity of core promoter recognition
complexes (Figure 2). Apparently unique to flies are the testes-specific TAFs, no hitter
(dTAF4b), cannonball (dTAF5b), ryan express (dTAF12b), and meiosis I arrest
(dTAF6b), all of which are required for spermatogenesis and normal germ cell-specific
gene expression (Hiller et al. 2001). Originally isolated from B-cells and highly
expressed in the testes and ovary, human TAF4b is the best understood tissue-specific
TAF. It associates with a slightly altered TFIID, which may or may not contain copies of
TAF4 and is required for expression of a subset of ovary-specific genes (Freiman et al.
2001; Geles et al. 2006). Furthermore, TAF4b knockout mice display defects in
folliculogenesis and spermatogenesis but no other apparent phenotypes, consistent with a
germ cell-specific pattern of expression and function (Falender et al. 2005). Similarly,

Chapter 1: Introduction 11
abundant expression of TAF7l coincides with reduced TAF7 expression in mouse
spermatids, and TAF7l associates with TBP and TAF1 presumably to form an altered
TFIID like complex (Pointud et al. 2003). Additionally, TAF7l knockout mice produce
abnormal sperm, have reduced fertility and overall weight, and show altered patterns of
testes-specific gene expression (Cheng et al. 2007). However, to date the full range of
phenotypes of these mice have not been characterized. The less well-studied TAF5l has
been associated with an increased risk of type I diabetes, but its pattern of expression and
association with TFIID remain to be determined (Cooper et al. 2007). Similarly, initial
characterization of TAF9l shows that it is a component of both TFIID and TBP-free
complexes, and that its loss affects the expression of a unique and overlapping set of
genes from that of TAF9 (Frontini et al. 2005). Finally, cell-type-specific alternative
splicing of TAF6 and possibly other TAFs may further expand the diversity of core
promoter recognition complexes (Weinzierl et al. 1993b).
TAFs in Metazoan Development and Differentiation
The central role TAFs and canonical TFIID in organismal development is
supported by the initial discovery of several TAFs in forward genetics screens and
confirmed by the requirement for many others as shown in depletion and knockout
experiments. Despite these findings, most TAFs, with the exception of tissue-specific
paralogs, are assumed to be ubiquitously expressed and canonical TFIID present at a
majority of promoters in nearly all tissue types. Hence, the first evidence of TFIID’s in
vivo function came from mutations in TAF4 and TAF6, which were homozygous

Chapter 1: Introduction 12
embryonic lethal and prevented proper complex formation in a sensitized Drosophila
suppressor screen (Karim et al. 1996; Sauer et al. 1996). Further analysis of the
heterozygous mutants showed a dosage dependent requirement for these TAFs as co-
activators of a known transcriptional activator at two important embryonic promoters
(Zhou et al. 1998; Pham et al. 1999). An independent mutation in TAF9 was found to
have dosage dependent affects on the expression of multiple genes, and these different
affects are potentially influenced by the genes differing core promoter architecture
(Soldatov et al. 1999). Finally, loss of function mutations in TAF1 were found to affect
the development of specific Drosophila tissues including the eye, ovaries, wing and
bristle by altering cell cycle progression and tissue-specific differentiation (Wassarman et
al. 2000).
Initial knockout studies in mice showed that loss of TAF8 or TAF10 destabilized
TFIID resulting in apoptosis of the blastocyst's inner cell mass and embryonic lethality,
though trophoblast viability appeared to be unaffected (Voss et al. 2000; Mohan et al.
2003). Conditional TAF knockout mice, which avoid the embryonic lethality phenotype,
are providing greater insight to tissue-specific TFIID function. As expected, a
conditional TAF4 allele failed to produce viable offspring when deleted in ES cells, but
was successfully employed to generate proliferative embryonic fibroblasts devoid of
TAF4 expression (Mengus et al. 2005). While these cell lines divided normally at routine
serum concentrations they displayed altered morphology, accelerated serum independent
growth, and disrupted expression of genes responsible for TGF signaling. Targeted
deletion of TAF4 in basal keratinocytes demonstrated that it is absolutely required for

Chapter 1: Introduction 13
fetal epidermis differentiation but partially dispensable for adult skin maintenance
(Fadloun et al. 2007). Furthermore, TAF4 inactivation increased the incidence of
epidermal hyperplasia, and this phenotype was traced to upregulation of multiple EGF
genes and downregulation of MAPK pathway components.
A similar approach to keratinocytes-specific deletion of TAF10 demonstrated
slightly altered requirements for this subunit relative to that of TAF4 (Indra et al. 2005).
Specifically, TAF10 is required for fetal epidermal differentiation, skin barrier formation,
and normal patterns of fetal skin gene expression, but dispensable for adult keratinocyte
homeostasis. Hence, adult epidermis lacking TAF10 showed normal gene expression
patterns, responses to UV irradiation, and proper regeneration after wounding. Recently,
the requirements of TAF10 in fetal liver development and adult homeostasis were
analyzed by targeted deletion of the same allele (Tatarakis et al. 2008). These studies
showed that TAF10 is required for the expression of most fetal hepatic genes and normal
liver organogenesis, and that its deletion disrupts normal TFIID complex assembly along
with general transcription factor promoter occupancy. Conversely, loss of TAF10 in
adult hepatocytes has no detectable adverse phenotypes, causes downregulation of only a
small subset of genes, and surprisingly derepresses genes that are normally silenced
neonatally. These observations led the authors to propose a multiclass gene model in
which some promoters require TFIID for their ongoing transcription, others are actively
repressed by TFIID occupancy, and the majority employ TFIID for the first round of fetal
transcription but continue to be activated in the adult despite its absence. New
conditional knockout models and novel approaches to in vivo TAF ablation, including

Chapter 1: Introduction 14
virally mediated RNA, will continue to enhance our understanding of TFIID’s function in
mammalian development and differentiation.
Embryonic and Neonatal Liver Development
The liver is unique to vertebrates and is responsible for many of the most critical
metabolic and homeostatic functions of adult organisms in addition to being required for
hematopoiesis during fetal and neonatal development. In the adult the liver is the
primary site of carbohydrate, lipid, and protein metabolism and storage. Additionally, it
is responsible for the synthesis and secretion of most serum proteins, including many
growth and coagulation factors and several digestive enzymes. Finally, the liver is a
critical site for the removal of endogenous waste products and exogenous toxins,
including most pharmaceuticals. Thus because of its central and varied role in
organismal function, there is great interest in understanding how the vertebrate liver
develops and is maintained (Zaret 2002; Zaret and Grompe 2008).
While the molecular mechanisms of liver organogenesis are not yet fully
understood, the morphogenetic steps of liver development have been well characterized.
At mouse embryonic day 6.5-7.5 or 15-16 in humans, the primitive streak forms anterior
to posterior along the ventral midline initiating gastrulation and leading to the creation of
the three germ layers (Figure 3). Of these the definitive endoderm results from
ingression of epiblast cells through anterior portions of the streak and eventually gives
rise to the organs of the embryonic gut including the liver, pancreas, lung, and thyroid; at
the same time the extra-embryonic visceral endoderm is displaced laterally (Lewis and

Chapter 1: Introduction 15
Tam 2006). Cell lineage experiments show that the hepatic endoderm arises soon after
gastrulation from lateral portions of the ventral foregut epithelium and a smaller group of
cells along the ventral midline epithelium, which are joined upon tube closure (Lawson et
al. 1991; Tremblay and Zaret 2005). As the hepatic endodermal cells rapidly proliferate,
they migrate into the neighboring septum transversum mesenchyme to form the liver bud,
which becomes increasingly vascularized and by embryonic day 10 develops the stromal
spaces that will eventually form the sinusoids of the adult liver. By embryonic day 9
these cells already express important early hepatic markers, including albumin (ALB), -
fetoprotein (AFP), and transthyretin (TTR) and are now considered hepatoblasts, the
bipotential progenitors that will differentiate into the major cell types of the adult liver
(Nobuyoshi Shiojiri1984; Germain et al. 1988; Gualdi et al. 1996). Hepatocytes
differentiate from hepatoblasts between embryonic day 14 and birth, and the neonatal
hepatocytes continue to proliferate and induce adult hepatic gene expression between
birth and neonatal day 21.
Hepatoblasts are small stem-like cells that are defined by their expression of both
hepatocyte and cholangiocyte markers and their ability to differentiate into both cell
types. They expand as a homogenous progenitor population in the liver bud until
embryonic day 14.5, when a subpopulation reduces its expression of hepatocyte genes
and differentiates towards the cholangiocyte or bile duct epithelial cell lineage; the
remaining hepatoblasts commit to the hepatocyte lineage between embryonic day 16 and
birth. Primary hepatoblasts can be reproducibly purified from mouse embryonic day 13.5
livers by enrichment of hepatoblast-specific markers including E-cadherin (CDH1) and

Chapter 1: Introduction 16
delta-like 1 (DLK), or depletion of the hematopoietic markers TER119 and CD45 (Nitou
et al. 2002; Tanimizu et al. 2003; Nava et al. 2005). Additionally human heptaoblasts
have been successfully purified and characterized by a variety of techniques, and shown
to expand and differentiate when transplanted to immunocompromised mice (Mahieu-
Caputo et al. 2004; Wauthier et al. 2008). Several groups have reported the ex vivo
culture of primary hepatoblasts and their faithful bipotential differentiation stimulated by
extracellular factors; in particular the interleukin-6 family member oncostatin M (OSM)
appears to be absolutely required for hepatocyte-specific differentiation (Kamiya et al.
1999; Kamiya et al. 2002). Further, immortalized hepatoblast cell lines that maintain
progenitor gene expression patterns and the ability to differentiate into cholangiocytes
and hepatocytes in vitro have been developed (Rogler 1997; Strick-Marchand and Weiss
2002; Tanimizu et al. 2004). Together these in vitro techniques have helped to elucidate
the signaling and transcriptional mechanisms of in vivo liver development (Strick-
Marchand et al. 2004; Tanimizu and Miyajima 2004; Ader et al. 2006). The extracellular
signaling events that lead to hepatic endoderm specification and hepatoblast commitment
have been extensively studied by tissue explant and gene knockout experiments. Hence,
hepatic fate decisions are initiated by suppression of Wnt and fibroblast growth factor 4
(FGF4) signaling in the foregut mesoderm and suppressed by active Wnt signaling in the
posterior gut mesoderm (Wells and Melton 2000). The hepatic endoderm’s anterior-
posterior position is further refined by retinoic acid signaling from the paraxial
mesoderm, and following tube closure, solidified by suppression of sonic hedgehog
(SHH) signaling in the dorsal endoderm (Apelqvist et al. 1997; Kumar et al. 2003).

Chapter 1: Introduction 17
Foregut hepatic differentiation is further stimulated by multiple FGFs secreted by the
cardiac mesoderm and bone morphogenetic proteins (BMPs) secreted by the septum
transversum mesenchyme (Jung et al. 1999; Rossi et al. 2001). While it is known that
specific extracellular signaling events alter intracellular mitogen-activated protein kinase
activity (MAPK) and cause tractable changes in cellular transcription factor expression
levels, the mechanism by which soluble factors transduce global transcription programs
to specify hepatoblast cell fate are not well understood (Wandzioch and Zaret 2009).
The transcriptional control of hepatic differentiation is currently attributed to a
large and dynamic network of liver-enriched classical sequence-specific activators
(Cereghini 1996; Costa et al. 2003). These proteins are categorized based on the
structural similarity of their DNA binding domains and include the onecut homeodomain
family (HNF-1), leucine zipper family (C/EBP), winged helix family (HNF-3/FOXA),
and nuclear hormone receptor family (HNF4, COUP-TFII,, LRH-1, FXR, PXR). While
these activators are widely detectable throughout multiple tissues and phases of liver
development, their onset and degree of expression correlates well with their unique
requirements in sequential phases of hepatic organogenesis (Kyrmizi et al. 2006). Hence,
the initial specification of hepatoblasts within the definitive endoderm requires HNF-3
and HNF-3 , which are expressed as early as gastrulation (Duncan et al. 1998; Lee et al.
2005). HNF-4 and GATA-6 are expressed in the developing hepatic endoderm and are
required for liver bud expansion and the hepatoblast to hepatocyte transition (Duncan et
al. 2000; Li et al. 2000; Zhao et al. 2005). HNF-1b and HNF-6 are first expressed in the
developing liver bud and are required for the lineage split of hepatoblasts to hepatocytes

Chapter 1: Introduction 18
and cholangiocytes (Clotman et al. 2002; Coffinier et al. 2002). Further, C/EBP , which
is first expressed at birth, is necessary for the expression of metabolic genes which will
function in the adult liver (Wang et al. 1995). The delicate spatial and temporal
regulation of liver-specific gene expression in both the developing tissue and the adult
organ is proposed to be achieved by the combinatorial activation and repression of each
unique promoter by this diverse toolbox of proteins; regulatory diversity is further
complicated by the ability of many of these factors to heterodimerize. Additionally, the
liver-enriched transcription factors have been shown to autoregulate their own and each
other’s expression throughout development and adult homeostasis (Odom et al. 2004;
Odom et al. 2006). While the circuitry of liver-enriched sequence-specific activators has
been extensively investigated, the role of core promoter recognition complexes, co-
activators, and chromatin remodelers in liver specification is remains to be understood.
Hepatic Regeneration
The liver is distinct among metazoan organs for it intrinsic regenerative ability
following acute physical, toxic, or viral injury (Fausto and Campbell 2003; Fausto et al.
2006). While much attention has been given to the potential role of resident liver stem
cells or oval cells in maintaining the chronically injured liver, the primary source of liver
regeneration in healthy animals is thought to be the limited proliferation of adult
hepatocytes (Figure 3). This unique regenerative capacity has made the liver a target of
human transplantation therapies as well as a model system for understanding adult stem
cell differentiation and organ plasticity.

Chapter 1: Introduction 19
Within 36-42 hours of partial hepectomy (PH), the most accepted experimental
model of hepatic injury, 95% of quiescent mouse hetapocytes synchronously transition
from G0 to G1 and enter S-phase to begin DNA replication (Grisham 1962; Mitchell and
Willenbring 2008). The hepatocytes divide once or twice and the full liver mass can be
restored within one to two weeks in rodents or approximately one to two months in
humans, although in most cases the original liver architecture is altered. Mouse models
of chronic liver injury suggest that adult hepatocytes have an even greater replicative
potential, which may allow for novel therapeutic applications. Hence, when wild type
hepatocytes are transplanted into urokinase plasminogen activaor (uPA) transgenic mice
they undergo up to 15 cell divisions almost fully replacing the necrotic host liver (Rhim
et al. 1994). Similarly, serial repopulation of the chronically injured fumarylacetoacetate
hydrolase (FAH) knockout mouse liver with wild type hepatocytes suggests that at least
80 rounds of division are possible (Overturf et al. 1996; Overturf et al. 1999).
Much like developing heptaoblasts, hepatocyte regeneration depends on a
complex network of extracellular signaling molecules and cellular transcription factors
(Fausto et al. 1995). Hence, within hours of hepatic injury nonparenchymal cell derived
tumor necrosis factor (TNF) and interleukin-6 (IL-6) stimulate the nuclear factor-kappa B
(NF B), signal transducer, and activator of transcription 3 (STAT3) pathways
respectively, inducing immediate early gene expression, promoting the G0 to G1
transition, and priming the quiescent growth factor immune hepatocytes for activation by
additional extracellular signals (Akerman et al. 1992; Webber et al. 1998; Iwai et al.
2001). Next mesenchyme derived hepatocyte growth factor (HGF) and hepatocyte

Chapter 1: Introduction 20
expressed transforming growth factor alpha (TGF ) activate the mitogen-activated
protein kinase (MAPK) and phosphoinositide-3-kinase (PI3K) cascades to promote
progression past the G1-S restriction point and induce expression of additional
transcription factors (Mead and Fausto 1989; Webber et al. 1993; Borowiak et al. 2004;
Huh et al. 2004). The activation of NFkB and STAT3 is the result of post translational
modifications and occurs independent of new protein synthesis, allowing these factors to
induce immediate early gene expression within hours of hepectomy (Cressman et al.
1994; Cressman et al. 1995). The activated immediate early genes include
transmembrane receptors required for continued mitogen responsiveness, cyclins and
cyclin dependent kinases which mediate cell cycle progression, and additional
transcription factors that drive the regenerative program (Mohn et al. 1991). Among
these transcription factors are known mitogen dependent cell cycle regulators not
normally expressed in the adult liver including c-Jun, c-Fos (AP-1), and c-Myc; the
overexpression or deletion of these genes has been shown to promote or impair liver
regeneration respectively (Thompson et al. 1986; Morello et al. 1990; Behrens et al.
2002). A second class of immediate early genes includes liver-enriched transcription
factors such as C/EBP which are responsible for the reestablishment of normal
hepatocytic function and coincident with these is an increase in the expression of liver
proteins such as albumin (ALB), 1-antitrypsin (AAT) and Phosphoenolpyruvate
carboxykinase (PEPCK) (Park et al. 1990; Diehl 1998). Intriguingly, gene expression
profiles of regenerative and neonatal hepatocytes are similar suggesting, a common
transcriptional program. More recently FoxM1B was shown to be an immediate late

Chapter 1: Introduction 21
gene in regenerative hepatocytes which is responsible for expression of cyclins regulating
the G2-M transition and exit from the regenerative program; consistent with this
overexpression, of FoxM1B can speed up liver regeneration or restore it in older
hepatocytes that have lost their regenerative capacity (Ye et al. 1999; Wang et al. 2001).
However, as with the embryo, the precise influence of ligands and signaling cascades on
intercellular transcriptional events during hepatic regeneration is not well understood.
Chronic toxic injury, certain genetic abnormalities, and age related degeneration
may limit or eliminate the hepatocytes proliferative capacity; under these conditions rare
resident liver stem cells, or oval cells, can expand and differentiate, contributing to
hepatic regeneration (Figure 3) (Fausto and Campbell 2003). While there has been
considerable disagreement concerning the oval cell’s phenotype and capacity, a
consensus exists that the portal region of the hepatic lobule contains a bipotential
progenitor with large oval nuclei that expresses hepatic, biliary, and hematopoietic
markers (Farber 1956; Dorrell et al. 2008). Though several groups have reported the
purification of hepatic progenitors from healthy liver, most of the work on oval cells
employs a model of chronic liver injury involving dietary deficiency, extended feeding of
carcinogenic or hepatotoxic compounds, and partial hepectomy (Wilson and Leduc 1958;
Dabeva et al. 1998; Preisegger et al. 1999; Wang et al. 2003a). Expanded oval cell
populations from these injury models have been purified based on differential
centrifugation or cell surface marker enrichment and used for in vivo bipotential
differentiation studies as well as the derivation of immortalized cell lines (Wilson and
Leduc 1958; Hayner et al. 1984; Yaswen et al. 1984; Germain et al. 1985; Dabeva et al.

Chapter 1: Introduction 22
1998; Preisegger et al. 1999; Petersen et al. 2003; Wang et al. 2003a). Furthermore,
purified oval cells and immortalized cell lines have been successfully used to partially
repopulate chronically injured liver models, providing the greatest evidence to date for
the oval cells proliferative and regenerative capacity (Wang et al. 2003b; Song et al.
2004). While oval cell expansion is likely dependent on many of the same extracellular
signaling pathways as hepatocyte proliferation, it also appears to require additional
unique signaling events typically associated with hematopoiesis, such as the SCF/c-kit
system (Fujio et al. 1994). Intriguingly, the oval cell’s temporal pattern of liver-enriched
transcription factor expression has been shown to closely mirror that of the developing
hepatoblast, suggesting that these two populations may differentiate through shared
transcriptional mechanisms. However transcription in oval cells has not been
investigated in any greater detail and warrants further study (Nagy et al. 1994).
Importantly, oval cell populations have been identified in the chronically injured human
liver, and their bipotential and proliferative capacity are beginning to be understood
(Haruna et al. 1996; Baumann et al. 1999; Roskams et al. 2004).
Embryonic Stem Cells and Therapeutic Approaches
The liver’s central place in many aspects of organismal function, combined with a
restricted supply of human liver donors and the complications associated with whole
organ transplantation, has generated considerable interest in cell based therapeutic
approaches to liver repopulation (Grompe 2006; Nussler et al. 2006). Donor derived
mature hepatocyte transplantation has been reported for the treatment of acute liver

Chapter 1: Introduction 23
failure, genetic metabolic disorders, and end stage cirrhosis with some success (Mito et
al. 1992; Fox et al. 1998; Bilir et al. 2000). Hepatocyte transplantation, has considerable
advantages over whole liver transplantation including the ability to treat multiple patients
from a single donor, decreased immunogenicity, cryopreservation of donor cells, and
reduced morbidity associated with the procedure itself. However, it also suffers from
considerable drawbacks such as donor supply, low engraftment rates, limited
proliferation of transplanted cells, and the inability to restore normal liver architecture
(Fox and Chowdhury 2004). Thus, a principal aim of hepatology research is to create an
indefinite source of autologous regenerative hepatocytes from patients’ own stem or
progenitor cells. Understanding the transcriptional mechanisms of hepatic differentiation
and regeneration is critical to this effort.
Embryonic stem cells (ES) hold great promise as a source of transplantable
hepatocytes because of their indefinite proliferative capacity and ability to differentiate
into every major cell type of the body (Keller 2005; Dalgetty et al. 2009). Mouse ES
cells have been successfully differentiated into functional hepatocytes and used to
partially restore liver function in a murine model of hepatic failure (Soto-Gutierrez et al.
2006). More critically, multiple groups have reported the directed differentiation of
human ES cells into hepatocyte like cells which faithfully recapitulate the gene
expression profiles and metabolic functions of primary hepatocytes (Duan et al. 2007;
Hay et al. 2007; Hay et al. 2008b). In most cases these differentiation protocols are based
on our current understanding of key signaling events required for hepatic development
and employ the same timing and extracellular signals observed in vivo (Schuldiner et al.

Chapter 1: Introduction 24
2000). Furthermore, new discoveries concerning in vivo liver development have been
successfully applied to improve the efficiency and accuracy of in vitro ES cell
differentiation toward hepatocytes (Hay et al. 2008a). Recent reports demonstrate that
these human ES cell derived hepatocytes can partially restore liver function when
transplanted into an immunocompromised mouse model, further supporting their
therapeutic potential (Basma et al. 2009). Similarly, multiple groups have
transdifferntiated adult hematopoietic stem cells into functional hepatocytes by
employing our understanding of key signaling events in liver developments thereby
avoiding the ethical issues associated with ES cell based therapies (Alison et al. 2000;
Lagasse et al. 2000). Thus, our expanding knowledge of the molecular mechanisms
required for hepatic development is currently being employed to generate potentially
therapeutic hepatocytes from non-hepatic stem cells.
The recent discovery that adult somatic cells can be reprogrammed into
proliferative pluripotent stem cells termed induced pluripotent stem cells (iPS) could
contribute greatly to our understanding of hepatogenesis, and hasten the advancement of
therapeutic liver repopulation (Yamanaka 2007; Jaenisch and Young 2008). Intriguingly,
iPS cells have been successfully generated from adult hepatocytes suggesting that the
mature cell type possesses all but a few factors required for proliferation, and raising the
possibility that damaged liver cells could be dedifferentiated, modified, and transplanted
(Aoi et al. 2008). While iPS cells have not yet been differentiated into functioning
hepatocytes, they have been specifically differentiated to cardiomyocytes, adipocytes,
retinal cells, and multiple hematopoitic lineages (Schenke-Layland et al. 2008; Hirami et

Chapter 1: Introduction 25
al. 2009; Senju et al. 2009; Zhang et al. 2009). Furthermore, comparative studies suggest
that iPS cells can be differentiated by the same protocols and with similar efficiency as
ES cells (Taura et al. 2009). Importantly, directed in vivo differentiation of genetically
modified autologous iPS cells followed by transplantation has been used to correct mouse
models of human disease including sickle cell anemia, fanconi anaemia, and hearing loss
(Nishimura et al. 2009; Raya et al. 2009). Such experiments raise the possibility that a
patient’s own somatic cells could be used for therapeutic liver repopulation.
Understanding the transcriptional mechanisms of liver development, regeneration, and
dedifferentiation is essential to realizing this potential.

Chapter 1: Introduction 26
Figure 1: Model of Preinitiation Complex Assembly
Sequence-specific activators (green) bind proximal (PA) and distal (DA) enhancer
elements within a genes regulatory DNA and recruit TFIID (yellow) to the core promoter.
These activators and/or TFIID in turn recruit chromatin remodeling complexes such as
SWI/SNF (orange), coactivators including Mediator (MED, blue), and TFIIA and TFIIB
(purple). The TFIID/TFIIA/TFIIB heterotrimer sequentially recruits TFIIE, TFIIF, PolII
(red), and TFIIH (purple), allowing for promoter escape and productive transcriptional
elongation.

Chapter 1: Introduction 27
H
PETATA
TFIID
DE
MED
PolII
E
F
SWI/SNF
BA

Chapter 1: Introduction 28
Figure 2: Combinatorial Model of Core Promoter Recognition Complex Diversity
Canonical TFIID consisting of TBP (red) and 15 associated TAFs (yellow) is
believed to be present at most promoters and in most cell types. Additional regulatory
complexity comes from TBP-free TAF containing complexes such as TFTC which
contain some TAFs plus additional unique proteins (burgundy), TBP-Related Factor
complexes such as TAF3-TRF3 (green), and alternate forms of TFIID that use tissue-
specific TAFs such as TAF5b and TAF7l (blue). Novel, as yet unidentified complexes
may be present in specific tissues or developmental programs.

Chapter 1: Introduction 29
TAF3
-TR
F3M
yoge
nesi
s
TAF4
b-II
DFo
llicu
loge
nesi
s/Sp
erm
atog
enes
is
TFI
IA-T
RF2
TFT
C
TAF7
l-II
DSp
erm
ioge
nesi
s
Can
onic
al T
FIID

Chapter 1: Introduction 30
Figure 3: Hepatic Development and Regeneration
Hepatoblasts, the common bipotential liver progenitors, develop from the
definitive endoderm between embryonic day 8.5 and 13.5. Starting at embryonic day
14.5 they differentiate to cholangiocytes or bile duct epithelial cells and hepatocytes.
Hepatocytes continue to proliferate and mature from before birth until at least neonatal
day 21. Following acute liver injury, mature hepatocytes reenter the cell cycle to
regenerate liver mass and function. Rare resident liver stem cells, or oval cells, may also
expand and differentiate contributing to liver regeneration.

Chapter 1: Introduction 31
Hea
d
Para
xial
Mes
oder
m
Som
ites
Car
diac
Mes
oder
m
Hep
atic
End
oder
mD
orsa
l End
oder
m
Ven
tral
End
oder
m
Sept
um T
rans
vers
um
Hep
atob
last
sE
8.5-
E14
.5
Adu
lt C
hola
ngio
cyte
sO
val C
ells
Neo
nata
l Hep
atoc
ytes
Adu
lt H
epat
ocyt
es
Cyc
ling
Hep
atoc
ytesIn
jury
and
Reg
ener
atio
nFe
tal a
nd N
eona
tal D
evel
opm
ent

Chapter 1: Introduction 32
References
Ader, T., Norel, R., Levoci, L., and Rogler, L.E. 2006. Transcriptional profiling implicates TGFbeta/BMP and Notch signaling pathways in ductular differentiation of fetal murine hepatoblasts. Mech Dev 123(2): 177-194.
Akerman, P., Cote, P., Yang, S.Q., McClain, C., Nelson, S., Bagby, G.J., and Diehl, A.M. 1992. Antibodies to tumor necrosis factor-alpha inhibit liver regeneration after partial hepatectomy. Am J Physiol 263(4 Pt 1): G579-585.
Albright, S.R. and Tjian, R. 2000. TAFs revisited: more data reveal new twists and confirm old ideas. Gene 242(1-2): 1-13.
Alison, M.R., Poulsom, R., Jeffery, R., Dhillon, A.P., Quaglia, A., Jacob, J., Novelli, M., Prentice, G., Williamson, J., and Wright, N.A. 2000. Hepatocytes from non-hepatic adult stem cells. Nature 406(6793): 257.
Andel, F., 3rd, Ladurner, A.G., Inouye, C., Tjian, R., and Nogales, E. 1999. Three-dimensional structure of the human TFIID-IIA-IIB complex. Science 286(5447): 2153-2156.
Aoi, T., Yae, K., Nakagawa, M., Ichisaka, T., Okita, K., Takahashi, K., Chiba, T., and Yamanaka, S. 2008. Generation of pluripotent stem cells from adult mouse liver and stomach cells. Science 321(5889): 699-702.
Aoyagi, N. and Wassarman, D.A. 2000. Genes encoding Drosophila melanogaster RNA polymerase II general transcription factors: diversity in TFIIA and TFIID components contributes to gene-specific transcriptional regulation. J Cell Biol 150(2): F45-50.
Apelqvist, A., Ahlgren, U., and Edlund, H. 1997. Sonic hedgehog directs specialised mesoderm differentiation in the intestine and pancreas. Curr Biol 7(10): 801-804.
Bartfai, R., Balduf, C., Hilton, T., Rathmann, Y., Hadzhiev, Y., Tora, L., Orban, L., and Muller, F. 2004. TBP2, a vertebrate-specific member of the TBP family, is required in embryonic development of zebrafish. Curr Biol 14(7): 593-598.

Chapter 1: Introduction 33
Basma, H., Soto-Gutierrez, A., Yannam, G.R., Liu, L., Ito, R., Yamamoto, T., Ellis, E.,
Carson, S.D., Sato, S., Chen, Y. et al. 2009. Differentiation and transplantation of human embryonic stem cell-derived hepatocytes. Gastroenterology 136(3): 990-999.
Baumann, U., Crosby, H.A., Ramani, P., Kelly, D.A., and Strain, A.J. 1999. Expression of the stem cell factor receptor c-kit in normal and diseased pediatric liver: identification of a human hepatic progenitor cell? Hepatology 30(1): 112-117.
Behrens, A., Sibilia, M., David, J.P., Mohle-Steinlein, U., Tronche, F., Schutz, G., and Wagner, E.F. 2002. Impaired neonatal hepatocyte proliferation and liver regeneration in mice lacking c-jun in the liver. EMBO J 21(7): 1782-1790.
Bilir, B.M., Guinette, D., Karrer, F., Kumpe, D.A., Krysl, J., Stephens, J., McGavran, L., Ostrowska, A., and Durham, J. 2000. Hepatocyte transplantation in acute liver failure. Liver Transpl 6(1): 32-40.
Borowiak, M., Garratt, A.N., Wustefeld, T., Strehle, M., Trautwein, C., and Birchmeier, C. 2004. Met provides essential signals for liver regeneration. Proc Natl Acad Sci U S A 101(29): 10608-10613.
Brand, M., Leurent, C., Mallouh, V., Tora, L., and Schultz, P. 1999. Three-dimensional structures of the TAFII-containing complexes TFIID and TFTC. Science 286(5447): 2151-2153.
Burke, T.W. and Kadonaga, J.T. 1997. The downstream core promoter element, DPE, is conserved from Drosophila to humans and is recognized by TAFII60 of Drosophila. Genes Dev 11(22): 3020-3031.
Burley, S.K. and Roeder, R.G. 1996. Biochemistry and structural biology of transcription factor IID (TFIID). Annu Rev Biochem 65: 769-799.
Cereghini, S. 1996. Liver-enriched transcription factors and hepatocyte differentiation. FASEB J 10(2): 267-282.

Chapter 1: Introduction 34
Chen, J.L., Attardi, L.D., Verrijzer, C.P., Yokomori, K., and Tjian, R. 1994. Assembly of
recombinant TFIID reveals differential coactivator requirements for distinct transcriptional activators. Cell 79(1): 93-105.
Cheng, Y., Buffone, M.G., Kouadio, M., Goodheart, M., Page, D.C., Gerton, G.L., Davidson, I., and Wang, P.J. 2007. Abnormal sperm in mice lacking the Taf7l gene. Mol Cell Biol 27(7): 2582-2589.
Chiang, C.M. and Roeder, R.G. 1995. Cloning of an intrinsic human TFIID subunit that interacts with multiple transcriptional activators. Science 267(5197): 531-536.
Cler, E., Papai, G., Schultz, P., and Davidson, I. 2009. Recent advances in understanding the structure and function of general transcription factor TFIID. Cell Mol Life Sci 66(13): 2123-2134.
Clotman, F., Lannoy, V.J., Reber, M., Cereghini, S., Cassiman, D., Jacquemin, P., Roskams, T., Rousseau, G.G., and Lemaigre, F.P. 2002. The onecut transcription factor HNF6 is required for normal development of the biliary tract. Development 129(8): 1819-1828.
Coffinier, C., Gresh, L., Fiette, L., Tronche, F., Schutz, G., Babinet, C., Pontoglio, M., Yaniv, M., and Barra, J. 2002. Bile system morphogenesis defects and liver dysfunction upon targeted deletion of HNF1beta. Development 129(8): 1829-1838.
Cooper, J.D., Smyth, D.J., Bailey, R., Payne, F., Downes, K., Godfrey, L.M., Masters, J., Zeitels, L.R., Vella, A., Walker, N.M. et al. 2007. The candidate genes TAF5L, TCF7, PDCD1, IL6 and ICAM1 cannot be excluded from having effects in type 1 diabetes. BMC Med Genet 8: 71.
Costa, R.H., Kalinichenko, V.V., Holterman, A.X., and Wang, X. 2003. Transcription factors in liver development, differentiation, and regeneration. Hepatology 38(6): 1331-1347.
Cressman, D.E., Diamond, R.H., and Taub, R. 1995. Rapid activation of the Stat3 transcription complex in liver regeneration. Hepatology 21(5): 1443-1449.

Chapter 1: Introduction 35
Cressman, D.E., Greenbaum, L.E., Haber, B.A., and Taub, R. 1994. Rapid activation of
post-hepatectomy factor/nuclear factor kappa B in hepatocytes, a primary response in the regenerating liver. J Biol Chem 269(48): 30429-30435.
Crick, F.H. 1958. On protein synthesis. Symp Soc Exp Biol 12: 138-163.
Crowley, T.E., Hoey, T., Liu, J.K., Jan, Y.N., Jan, L.Y., and Tjian, R. 1993. A new factor related to TATA-binding protein has highly restricted expression patterns in Drosophila. Nature 361(6412): 557-561.
Dabeva, M.D., Laconi, E., Oren, R., Petkov, P.M., Hurston, E., and Shafritz, D.A. 1998. Liver regeneration and alpha-fetoprotein messenger RNA expression in the retrorsine model for hepatocyte transplantation. Cancer Res 58(24): 5825-5834.
Dalgetty, D.M., Medine, C.N., Iredale, J.P., and Hay, D.C. 2009. Progress and future challenges in stem cell-derived liver technologies. Am J Physiol Gastrointest Liver Physiol 297(2): G241-248.
Dantonel, J.C., Quintin, S., Lakatos, L., Labouesse, M., and Tora, L. 2000. TBP-like factor is required for embryonic RNA polymerase II transcription in C. elegans. Mol Cell 6(3): 715-722.
Dantonel, J.C., Wurtz, J.M., Poch, O., Moras, D., and Tora, L. 1999. The TBP-like factor: an alternative transcription factor in metazoa? Trends Biochem Sci 24(9): 335-339.
Deato, M.D., Marr, M.T., Sottero, T., Inouye, C., Hu, P., and Tjian, R. 2008. MyoD targets TAF3/TRF3 to activate myogenin transcription. Mol Cell 32(1): 96-105.
Deato, M.D. and Tjian, R. 2007. Switching of the core transcription machinery during myogenesis. Genes Dev 21(17): 2137-2149.
Diehl, A.M. 1998. Roles of CCAAT/enhancer-binding proteins in regulation of liver regenerative growth. J Biol Chem 273(47): 30843-30846.

Chapter 1: Introduction 36
Dorrell, C., Erker, L., Lanxon-Cookson, K.M., Abraham, S.L., Victoroff, T., Ro, S.,
Canaday, P.S., Streeter, P.R., and Grompe, M. 2008. Surface markers for the murine oval cell response. Hepatology 48(4): 1282-1291.
Duan, Y., Catana, A., Meng, Y., Yamamoto, N., He, S., Gupta, S., Gambhir, S.S., and Zern, M.A. 2007. Differentiation and enrichment of hepatocyte-like cells from human embryonic stem cells in vitro and in vivo. Stem Cells 25(12): 3058-3068.
Duncan, M.K., Cvekl, A., Li, X., and Piatigorsky, J. 2000. Truncated forms of Pax-6 disrupt lens morphology in transgenic mice. Invest Ophthalmol Vis Sci 41(2): 464-473.
Duncan, S.A., Navas, M.A., Dufort, D., Rossant, J., and Stoffel, M. 1998. Regulation of a transcription factor network required for differentiation and metabolism. Science 281(5377): 692-695.
Dynlacht, B.D., Hoey, T., and Tjian, R. 1991. Isolation of coactivators associated with the TATA-binding protein that mediate transcriptional activation. Cell 66(3): 563-576.
Fadloun, A., Kobi, D., Pointud, J.C., Indra, A.K., Teletin, M., Bole-Feysot, C., Testoni, B., Mantovani, R., Metzger, D., Mengus, G. et al. 2007. The TFIID subunit TAF4 regulates keratinocyte proliferation and has cell-autonomous and non-cell-autonomous tumour suppressor activity in mouse epidermis. Development 134(16): 2947-2958.
Falender, A.E., Freiman, R.N., Geles, K.G., Lo, K.C., Hwang, K., Lamb, D.J., Morris, P.L., Tjian, R., and Richards, J.S. 2005. Maintenance of spermatogenesis requires TAF4b, a gonad-specific subunit of TFIID. Genes Dev 19(7): 794-803.
Farber, E. 1956. Similarities in the sequence of early histological changes induced in the liver of the rat by ethionine, 2-acetylamino-fluorene, and 3'-methyl-4-dimethylaminoazobenzene. Cancer Res 16(2): 142-148.
Fausto, N. and Campbell, J.S. 2003. The role of hepatocytes and oval cells in liver regeneration and repopulation. Mech Dev 120(1): 117-130.

Chapter 1: Introduction 37
Fausto, N., Campbell, J.S., and Riehle, K.J. 2006. Liver regeneration. Hepatology 43(2
Suppl 1): S45-53.
Fausto, N., Laird, A.D., and Webber, E.M. 1995. Liver regeneration. 2. Role of growth factors and cytokines in hepatic regeneration. FASEB J 9(15): 1527-1536.
Fox, I.J. and Chowdhury, J.R. 2004. Hepatocyte transplantation. Am J Transplant 4 Suppl 6: 7-13.
Fox, I.J., Chowdhury, J.R., Kaufman, S.S., Goertzen, T.C., Chowdhury, N.R., Warkentin, P.I., Dorko, K., Sauter, B.V., and Strom, S.C. 1998. Treatment of the Crigler-Najjar syndrome type I with hepatocyte transplantation. N Engl J Med 338(20): 1422-1426.
Freiman, R.N., Albright, S.R., Zheng, S., Sha, W.C., Hammer, R.E., and Tjian, R. 2001. Requirement of tissue-selective TBP-associated factor TAFII105 in ovarian development. Science 293(5537): 2084-2087.
Frontini, M., Soutoglou, E., Argentini, M., Bole-Feysot, C., Jost, B., Scheer, E., and Tora, L. 2005. TAF9b (formerly TAF9L) is a bona fide TAF that has unique and overlapping roles with TAF9. Mol Cell Biol 25(11): 4638-4649.
Fujio, K., Evarts, R.P., Hu, Z., Marsden, E.R., and Thorgeirsson, S.S. 1994. Expression of stem cell factor and its receptor, c-kit, during liver regeneration from putative stem cells in adult rat. Lab Invest 70(4): 511-516.
Gazdag, E., Rajkovic, A., Torres-Padilla, M.E., and Tora, L. 2007. Analysis of TATA-binding protein 2 (TBP2) and TBP expression suggests different roles for the two proteins in regulation of gene expression during oogenesis and early mouse development. Reproduction 134(1): 51-62.
Geles, K.G., Freiman, R.N., Liu, W.L., Zheng, S., Voronina, E., and Tjian, R. 2006. Cell-type-selective induction of c-jun by TAF4b directs ovarian-specific transcription networks. Proc Natl Acad Sci U S A 103(8): 2594-2599.

Chapter 1: Introduction 38
Germain, L., Blouin, M.J., and Marceau, N. 1988. Biliary epithelial and hepatocytic cell
lineage relationships in embryonic rat liver as determined by the differential expression of cytokeratins, alpha-fetoprotein, albumin, and cell surface-exposed components. Cancer Res 48(17): 4909-4918.
Germain, L., Goyette, R., and Marceau, N. 1985. Differential cytokeratin and alpha-fetoprotein expression in morphologically distinct epithelial cells emerging at the early stage of rat hepatocarcinogenesis. Cancer Res 45(2): 673-681.
Goodrich, J.A., Hoey, T., Thut, C.J., Admon, A., and Tjian, R. 1993. Drosophila TAFII40 interacts with both a VP16 activation domain and the basal transcription factor TFIIB. Cell 75(3): 519-530.
Grant, P.A., Schieltz, D., Pray-Grant, M.G., Steger, D.J., Reese, J.C., Yates, J.R., 3rd, and Workman, J.L. 1998. A subset of TAF(II)s are integral components of the SAGA complex required for nucleosome acetylation and transcriptional stimulation. Cell 94(1): 45-53.
Grisham, J.W. 1962. A morphologic study of deoxyribonucleic acid synthesis and cell proliferation in regenerating rat liver; autoradiography with thymidine-H3. Cancer Res 22: 842-849.
Grompe, M. 2006. Principles of therapeutic liver repopulation. J Inherit Metab Dis 29(2-3): 421-425.
Gualdi, R., Bossard, P., Zheng, M., Hamada, Y., Coleman, J.R., and Zaret, K.S. 1996. Hepatic specification of the gut endoderm in vitro: cell signaling and transcriptional control. Genes Dev 10(13): 1670-1682.
Ha, I., Roberts, S., Maldonado, E., Sun, X., Kim, L.U., Green, M., and Reinberg, D. 1993. Multiple functional domains of human transcription factor IIB: distinct interactions with two general transcription factors and RNA polymerase II. Genes Dev 7(6): 1021-1032.
Hansen, S.K., Takada, S., Jacobson, R.H., Lis, J.T., and Tjian, R. 1997. Transcription properties of a cell type-specific TATA-binding protein, TRF. Cell 91(1): 71-83.

Chapter 1: Introduction 39
Hart, D.O., Raha, T., Lawson, N.D., and Green, M.R. 2007. Initiation of zebrafish
haematopoiesis by the TATA-box-binding protein-related factor Trf3. Nature 450(7172): 1082-1085.
Haruna, Y., Saito, K., Spaulding, S., Nalesnik, M.A., and Gerber, M.A. 1996. Identification of bipotential progenitor cells in human liver development. Hepatology 23(3): 476-481.
Hay, D.C., Fletcher, J., Payne, C., Terrace, J.D., Gallagher, R.C., Snoeys, J., Black, J.R., Wojtacha, D., Samuel, K., Hannoun, Z. et al. 2008a. Highly efficient differentiation of hESCs to functional hepatic endoderm requires ActivinA and Wnt3a signaling. Proc Natl Acad Sci U S A 105(34): 12301-12306.
Hay, D.C., Zhao, D., Fletcher, J., Hewitt, Z.A., McLean, D., Urruticoechea-Uriguen, A., Black, J.R., Elcombe, C., Ross, J.A., Wolf, R. et al. 2008b. Efficient differentiation of hepatocytes from human embryonic stem cells exhibiting markers recapitulating liver development in vivo. Stem Cells 26(4): 894-902.
Hay, D.C., Zhao, D., Ross, A., Mandalam, R., Lebkowski, J., and Cui, W. 2007. Direct differentiation of human embryonic stem cells to hepatocyte-like cells exhibiting functional activities. Cloning Stem Cells 9(1): 51-62.
Hayner, N.T., Braun, L., Yaswen, P., Brooks, M., and Fausto, N. 1984. Isozyme profiles of oval cells, parenchymal cells, and biliary cells isolated by centrifugal elutriation from normal and preneoplastic livers. Cancer Res 44(1): 332-338.
Hiller, M.A., Lin, T.Y., Wood, C., and Fuller, M.T. 2001. Developmental regulation of transcription by a tissue-specific TAF homolog. Genes Dev 15(8): 1021-1030.
Hirami, Y., Osakada, F., Takahashi, K., Okita, K., Yamanaka, S., Ikeda, H., Yoshimura, N., and Takahashi, M. 2009. Generation of retinal cells from mouse and human induced pluripotent stem cells. Neurosci Lett 458(3): 126-131.
Hochheimer, A. and Tjian, R. 2003. Diversified transcription initiation complexes expand promoter selectivity and tissue-specific gene expression. Genes Dev 17(11): 1309-1320.

Chapter 1: Introduction 40
Hochheimer, A., Zhou, S., Zheng, S., Holmes, M.C., and Tjian, R. 2002. TRF2 associates
with DREF and directs promoter-selective gene expression in Drosophila. Nature 420(6914): 439-445.
Holmes, M.C. and Tjian, R. 2000. Promoter-selective properties of the TBP-related factor TRF1. Science 288(5467): 867-870.
Holstege, F.C., van der Vliet, P.C., and Timmers, H.T. 1996. Opening of an RNA polymerase II promoter occurs in two distinct steps and requires the basal transcription factors IIE and IIH. EMBO J 15(7): 1666-1677.
Huh, C.G., Factor, V.M., Sanchez, A., Uchida, K., Conner, E.A., and Thorgeirsson, S.S. 2004. Hepatocyte growth factor/c-met signaling pathway is required for efficient liver regeneration and repair. Proc Natl Acad Sci U S A 101(13): 4477-4482.
Huisinga, K.L. and Pugh, B.F. 2004. A genome-wide housekeeping role for TFIID and a highly regulated stress-related role for SAGA in Saccharomyces cerevisiae. Mol Cell 13(4): 573-585.
Indra, A.K., Mohan, W.S., 2nd, Frontini, M., Scheer, E., Messaddeq, N., Metzger, D., and Tora, L. 2005. TAF10 is required for the establishment of skin barrier function in foetal, but not in adult mouse epidermis. Dev Biol 285(1): 28-37.
Iwai, M., Cui, T.X., Kitamura, H., Saito, M., and Shimazu, T. 2001. Increased secretion of tumour necrosis factor and interleukin 6 from isolated, perfused liver of rats after partial hepatectomy. Cytokine 13(1): 60-64.
Jacobi, U.G., Akkers, R.C., Pierson, E.S., Weeks, D.L., Dagle, J.M., and Veenstra, G.J. 2007. TBP paralogs accommodate metazoan- and vertebrate-specific developmental gene regulation. EMBO J 26(17): 3900-3909.
Jacobson, R.H., Ladurner, A.G., King, D.S., and Tjian, R. 2000. Structure and function of a human TAFII250 double bromodomain module. Science 288(5470): 1422-1425.
Jaenisch, R. and Young, R. 2008. Stem cells, the molecular circuitry of pluripotency and nuclear reprogramming. Cell 132(4): 567-582.

Chapter 1: Introduction 41
Jallow, Z., Jacobi, U.G., Weeks, D.L., Dawid, I.B., and Veenstra, G.J. 2004. Specialized
and redundant roles of TBP and a vertebrate-specific TBP paralog in embryonic gene regulation in Xenopus. Proc Natl Acad Sci U S A 101(37): 13525-13530.
Jung, J., Zheng, M., Goldfarb, M., and Zaret, K.S. 1999. Initiation of mammalian liver development from endoderm by fibroblast growth factors. Science 284(5422): 1998-2003.
Juven-Gershon, T., Hsu, J.Y., Theisen, J.W., and Kadonaga, J.T. 2008. The RNA polymerase II core promoter - the gateway to transcription. Curr Opin Cell Biol 20(3): 253-259.
Kamiya, A., Kinoshita, T., Ito, Y., Matsui, T., Morikawa, Y., Senba, E., Nakashima, K., Taga, T., Yoshida, K., Kishimoto, T. et al. 1999. Fetal liver development requires a paracrine action of oncostatin M through the gp130 signal transducer. EMBO J 18(8): 2127-2136.
Kamiya, A., Kojima, N., Kinoshita, T., Sakai, Y., and Miyaijma, A. 2002. Maturation of fetal hepatocytes in vitro by extracellular matrices and oncostatin M: induction of tryptophan oxygenase. Hepatology 35(6): 1351-1359.
Karim, F.D., Chang, H.C., Therrien, M., Wassarman, D.A., Laverty, T., and Rubin, G.M. 1996. A screen for genes that function downstream of Ras1 during Drosophila eye development. Genetics 143(1): 315-329.
Keller, G. 2005. Embryonic stem cell differentiation: emergence of a new era in biology and medicine. Genes Dev 19(10): 1129-1155.
King, N., Westbrook, M.J., Young, S.L., Kuo, A., Abedin, M., Chapman, J., Fairclough, S., Hellsten, U., Isogai, Y., Letunic, I. et al. 2008. The genome of the choanoflagellate Monosiga brevicollis and the origin of metazoans. Nature 451(7180): 783-788.
Kumar, M., Jordan, N., Melton, D., and Grapin-Botton, A. 2003. Signals from lateral plate mesoderm instruct endoderm toward a pancreatic fate. Dev Biol 259(1): 109-122.

Chapter 1: Introduction 42
Kyrmizi, I., Hatzis, P., Katrakili, N., Tronche, F., Gonzalez, F.J., and Talianidis, I. 2006.
Plasticity and expanding complexity of the hepatic transcription factor network during liver development. Genes Dev 20(16): 2293-2305.
Lagasse, E., Connors, H., Al-Dhalimy, M., Reitsma, M., Dohse, M., Osborne, L., Wang, X., Finegold, M., Weissman, I.L., and Grompe, M. 2000. Purified hematopoietic stem cells can differentiate into hepatocytes in vivo. Nat Med 6(11): 1229-1234.
Lawson, K.A., Meneses, J.J., and Pedersen, R.A. 1991. Clonal analysis of epiblast fate during germ layer formation in the mouse embryo. Development 113(3): 891-911.
Lee, C.S., Friedman, J.R., Fulmer, J.T., and Kaestner, K.H. 2005. The initiation of liver development is dependent on Foxa transcription factors. Nature 435(7044): 944-947.
Leurent, C., Sanders, S., Ruhlmann, C., Mallouh, V., Weil, P.A., Kirschner, D.B., Tora, L., and Schultz, P. 2002. Mapping histone fold TAFs within yeast TFIID. EMBO J 21(13): 3424-3433.
Leurent, C., Sanders, S.L., Demeny, M.A., Garbett, K.A., Ruhlmann, C., Weil, P.A., Tora, L., and Schultz, P. 2004. Mapping key functional sites within yeast TFIID. EMBO J 23(4): 719-727.
Lewis, S.L. and Tam, P.P. 2006. Definitive endoderm of the mouse embryo: formation, cell fates, and morphogenetic function. Dev Dyn 235(9): 2315-2329.
Li, J., Ning, G., and Duncan, S.A. 2000. Mammalian hepatocyte differentiation requires the transcription factor HNF-4alpha. Genes Dev 14(4): 464-474.
Lu, H., Zawel, L., Fisher, L., Egly, J.M., and Reinberg, D. 1992. Human general transcription factor IIH phosphorylates the C-terminal domain of RNA polymerase II. Nature 358(6388): 641-645.
Mahieu-Caputo, D., Allain, J.E., Branger, J., Coulomb, A., Delgado, J.P., Andreoletti, M., Mainot, S., Frydman, R., Leboulch, P., Di Santo, J.P. et al. 2004. Repopulation of athymic mouse liver by cryopreserved early human fetal hepatoblasts. Hum Gene Ther 15(12): 1219-1228.

Chapter 1: Introduction 43
Maldonado, E., Ha, I., Cortes, P., Weis, L., and Reinberg, D. 1990. Factors involved in
specific transcription by mammalian RNA polymerase II: role of transcription factors IIA, IID, and IIB during formation of a transcription-competent complex. Mol Cell Biol 10(12): 6335-6347.
Martinez, E., Kundu, T.K., Fu, J., and Roeder, R.G. 1998. A human SPT3-TAFII31-GCN5-L acetylase complex distinct from transcription factor IID. J Biol Chem 273(37): 23781-23785.
Matsui, T., Segall, J., Weil, P.A., and Roeder, R.G. 1980. Multiple factors required for accurate initiation of transcription by purified RNA polymerase II. J Biol Chem 255(24): 11992-11996.
Maxon, M.E., Goodrich, J.A., and Tjian, R. 1994. Transcription factor IIE binds preferentially to RNA polymerase IIa and recruits TFIIH: a model for promoter clearance. Genes Dev 8(5): 515-524.
Mead, J.E. and Fausto, N. 1989. Transforming growth factor alpha may be a physiological regulator of liver regeneration by means of an autocrine mechanism. Proc Natl Acad Sci U S A 86(5): 1558-1562.
Mengus, G., Fadloun, A., Kobi, D., Thibault, C., Perletti, L., Michel, I., and Davidson, I. 2005. TAF4 inactivation in embryonic fibroblasts activates TGF beta signalling and autocrine growth. EMBO J 24(15): 2753-2767.
Mitchell, C. and Willenbring, H. 2008. A reproducible and well-tolerated method for 2/3 partial hepatectomy in mice. Nat Protoc 3(7): 1167-1170.
Mito, M., Kusano, M., and Kawaura, Y. 1992. Hepatocyte transplantation in man. Transplant Proc 24(6): 3052-3053.
Mohan, W.S., Jr., Scheer, E., Wendling, O., Metzger, D., and Tora, L. 2003. TAF10 (TAF(II)30) is necessary for TFIID stability and early embryogenesis in mice. Mol Cell Biol 23(12): 4307-4318.

Chapter 1: Introduction 44
Mohn, K.L., Laz, T.M., Hsu, J.C., Melby, A.E., Bravo, R., and Taub, R. 1991. The
immediate-early growth response in regenerating liver and insulin-stimulated H-35 cells: comparison with serum-stimulated 3T3 cells and identification of 41 novel immediate-early genes. Mol Cell Biol 11(1): 381-390.
Moore, P.A., Ozer, J., Salunek, M., Jan, G., Zerby, D., Campbell, S., and Lieberman, P.M. 1999. A human TATA binding protein-related protein with altered DNA binding specificity inhibits transcription from multiple promoters and activators. Mol Cell Biol 19(11): 7610-7620.
Morello, D., Fitzgerald, M.J., Babinet, C., and Fausto, N. 1990. c-myc, c-fos, and c-jun regulation in the regenerating livers of normal and H-2K/c-myc transgenic mice. Mol Cell Biol 10(6): 3185-3193.
Muller, F., Demeny, M.A., and Tora, L. 2007. New problems in RNA polymerase II transcription initiation: matching the diversity of core promoters with a variety of promoter recognition factors. J Biol Chem 282(20): 14685-14689.
Nagy, P., Bisgaard, H.C., and Thorgeirsson, S.S. 1994. Expression of hepatic transcription factors during liver development and oval cell differentiation. J Cell Biol 126(1): 223-233.
Nagy, Z. and Tora, L. 2007. Distinct GCN5/PCAF-containing complexes function as co-activators and are involved in transcription factor and global histone acetylation. Oncogene 26(37): 5341-5357.
Nava, S., Westgren, M., Jaksch, M., Tibell, A., Broome, U., Ericzon, B.G., and Sumitran-Holgersson, S. 2005. Characterization of cells in the developing human liver. Differentiation 73(5): 249-260.
Nishimura, K., Nakagawa, T., Ono, K., Ogita, H., Sakamoto, T., Yamamoto, N., Okita, K., Yamanaka, S., and Ito, J. 2009. Transplantation of mouse induced pluripotent stem cells into the cochlea. Neuroreport.
Nitou, M., Sugiyama, Y., Ishikawa, K., and Shiojiri, N. 2002. Purification of fetal mouse hepatoblasts by magnetic beads coated with monoclonal anti-e-cadherin antibodies and their in vitro culture. Exp Cell Res 279(2): 330-343.

Chapter 1: Introduction 45
NOBUYOSHI SHIOJIRI. 1984. Analysis of Differentiation of Hepatocytes and Bile
Duct Cells in Developing Mouse Liver by Albumin Immunofluorescence. Development Growth & Differentiation 26(6): 555-561.
Nussler, A., Konig, S., Ott, M., Sokal, E., Christ, B., Thasler, W., Brulport, M., Gabelein, G., Schormann, W., Schulze, M. et al. 2006. Present status and perspectives of cell-based therapies for liver diseases. J Hepatol 45(1): 144-159.
Odom, D.T., Dowell, R.D., Jacobsen, E.S., Nekludova, L., Rolfe, P.A., Danford, T.W., Gifford, D.K., Fraenkel, E., Bell, G.I., and Young, R.A. 2006. Core transcriptional regulatory circuitry in human hepatocytes. Mol Syst Biol 2: 2006 0017.
Odom, D.T., Zizlsperger, N., Gordon, D.B., Bell, G.W., Rinaldi, N.J., Murray, H.L., Volkert, T.L., Schreiber, J., Rolfe, P.A., Gifford, D.K. et al. 2004. Control of pancreas and liver gene expression by HNF transcription factors. Science 303(5662): 1378-1381.
Ogryzko, V.V., Kotani, T., Zhang, X., Schiltz, R.L., Howard, T., Yang, X.J., Howard, B.H., Qin, J., and Nakatani, Y. 1998. Histone-like TAFs within the PCAF histone acetylase complex. Cell 94(1): 35-44.
Overturf, K., Al-Dhalimy, M., Finegold, M., and Grompe, M. 1999. The repopulation potential of hepatocyte populations differing in size and prior mitotic expansion. Am J Pathol 155(6): 2135-2143.
Overturf, K., Al-Dhalimy, M., Tanguay, R., Brantly, M., Ou, C.N., Finegold, M., and Grompe, M. 1996. Hepatocytes corrected by gene therapy are selected in vivo in a murine model of hereditary tyrosinaemia type I. Nat Genet 12(3): 266-273.
Park, E.A., Roesler, W.J., Liu, J., Klemm, D.J., Gurney, A.L., Thatcher, J.D., Shuman, J., Friedman, A., and Hanson, R.W. 1990. The role of the CCAAT/enhancer-binding protein in the transcriptional regulation of the gene for phosphoenolpyruvate carboxykinase (GTP). Mol Cell Biol 10(12): 6264-6272.

Chapter 1: Introduction 46
Persengiev, S.P., Zhu, X., Dixit, B.L., Maston, G.A., Kittler, E.L., and Green, M.R. 2003.
TRF3, a TATA-box-binding protein-related factor, is vertebrate-specific and widely expressed. Proc Natl Acad Sci U S A 100(25): 14887-14891.
Petersen, B.E., Grossbard, B., Hatch, H., Pi, L., Deng, J., and Scott, E.W. 2003. Mouse A6-positive hepatic oval cells also express several hematopoietic stem cell markers. Hepatology 37(3): 632-640.
Pham, A.D., Muller, S., and Sauer, F. 1999. Mesoderm-determining transcription in Drosophila is alleviated by mutations in TAF(II)60 and TAF(II)110. Mech Dev 84(1-2): 3-16.
Pointud, J.C., Mengus, G., Brancorsini, S., Monaco, L., Parvinen, M., Sassone-Corsi, P., and Davidson, I. 2003. The intracellular localisation of TAF7L, a paralogue of transcription factor TFIID subunit TAF7, is developmentally regulated during male germ-cell differentiation. J Cell Sci 116(Pt 9): 1847-1858.
Pray-Grant, M.G., Schieltz, D., McMahon, S.J., Wood, J.M., Kennedy, E.L., Cook, R.G., Workman, J.L., Yates, J.R., 3rd, and Grant, P.A. 2002. The novel SLIK histone acetyltransferase complex functions in the yeast retrograde response pathway. Mol Cell Biol 22(24): 8774-8786.
Preisegger, K.H., Factor, V.M., Fuchsbichler, A., Stumptner, C., Denk, H., and Thorgeirsson, S.S. 1999. Atypical ductular proliferation and its inhibition by transforming growth factor beta1 in the 3,5-diethoxycarbonyl-1,4-dihydrocollidine mouse model for chronic alcoholic liver disease. Lab Invest 79(2): 103-109.
Pugh, B.F. and Tjian, R. 1991. Transcription from a TATA-less promoter requires a multisubunit TFIID complex. Genes Dev 5(11): 1935-1945.
Rabenstein, M.D., Zhou, S., Lis, J.T., and Tjian, R. 1999. TATA box-binding protein (TBP)-related factor 2 (TRF2), a third member of the TBP family. Proc Natl Acad Sci U S A 96(9): 4791-4796.

Chapter 1: Introduction 47
Raya, A., Rodriguez-Piza, I., Guenechea, G., Vassena, R., Navarro, S., Barrero, M.J.,
Consiglio, A., Castella, M., Rio, P., Sleep, E. et al. 2009. Disease-corrected haematopoietic progenitors from Fanconi anaemia induced pluripotent stem cells. Nature 460(7251): 53-59.
Reina, J.H. and Hernandez, N. 2007. On a roll for new TRF targets. Genes Dev 21(22): 2855-2860.
Rhim, J.A., Sandgren, E.P., Degen, J.L., Palmiter, R.D., and Brinster, R.L. 1994. Replacement of diseased mouse liver by hepatic cell transplantation. Science 263(5150): 1149-1152.
Rogler, L.E. 1997. Selective bipotential differentiation of mouse embryonic hepatoblasts in vitro. Am J Pathol 150(2): 591-602.
Roskams, T.A., Theise, N.D., Balabaud, C., Bhagat, G., Bhathal, P.S., Bioulac-Sage, P., Brunt, E.M., Crawford, J.M., Crosby, H.A., Desmet, V. et al. 2004. Nomenclature of the finer branches of the biliary tree: canals, ductules, and ductular reactions in human livers. Hepatology 39(6): 1739-1745.
Rossi, J.M., Dunn, N.R., Hogan, B.L., and Zaret, K.S. 2001. Distinct mesodermal signals, including BMPs from the septum transversum mesenchyme, are required in combination for hepatogenesis from the endoderm. Genes Dev 15(15): 1998-2009.
Sanders, S.L., Garbett, K.A., and Weil, P.A. 2002. Molecular characterization of Saccharomyces cerevisiae TFIID. Mol Cell Biol 22(16): 6000-6013.
Sauer, F., Wassarman, D.A., Rubin, G.M., and Tjian, R. 1996. TAF(II)s mediate activation of transcription in the Drosophila embryo. Cell 87(7): 1271-1284.
Schenke-Layland, K., Rhodes, K.E., Angelis, E., Butylkova, Y., Heydarkhan-Hagvall, S., Gekas, C., Zhang, R., Goldhaber, J.I., Mikkola, H.K., Plath, K. et al. 2008. Reprogrammed mouse fibroblasts differentiate into cells of the cardiovascular and hematopoietic lineages. Stem Cells 26(6): 1537-1546.

Chapter 1: Introduction 48
Schuldiner, M., Yanuka, O., Itskovitz-Eldor, J., Melton, D.A., and Benvenisty, N. 2000.
Effects of eight growth factors on the differentiation of cells derived from human embryonic stem cells. Proc Natl Acad Sci U S A 97(21): 11307-11312.
Selleck, W., Howley, R., Fang, Q., Podolny, V., Fried, M.G., Buratowski, S., and Tan, S. 2001. A histone fold TAF octamer within the yeast TFIID transcriptional coactivator. Nat Struct Biol 8(8): 695-700.
Senju, S., Haruta, M., Matsunaga, Y., Fukushima, S., Ikeda, T., Takahashi, K., Okita, K., Yamanaka, S., and Nishimura, Y. 2009. Characterization of dendritic cells and macrophages generated by directed differentiation from mouse induced pluripotent stem cells. Stem Cells 27(5): 1021-1031.
Smale, S.T. and Kadonaga, J.T. 2003. The RNA polymerase II core promoter. Annu Rev Biochem 72: 449-479.
Soldatov, A., Nabirochkina, E., Georgieva, S., Belenkaja, T., and Georgiev, P. 1999. TAFII40 protein is encoded by the e(y)1 gene: biological consequences of mutations. Mol Cell Biol 19(5): 3769-3778.
Song, S., Witek, R.P., Lu, Y., Choi, Y.K., Zheng, D., Jorgensen, M., Li, C., Flotte, T.R., and Petersen, B.E. 2004. Ex vivo transduced liver progenitor cells as a platform for gene therapy in mice. Hepatology 40(4): 918-924.
Soto-Gutierrez, A., Kobayashi, N., Rivas-Carrillo, J.D., Navarro-Alvarez, N., Zhao, D., Okitsu, T., Noguchi, H., Basma, H., Tabata, Y., Chen, Y. et al. 2006. Reversal of mouse hepatic failure using an implanted liver-assist device containing ES cell-derived hepatocytes. Nat Biotechnol 24(11): 1412-1419.
Strick-Marchand, H., Morosan, S., Charneau, P., Kremsdorf, D., and Weiss, M.C. 2004. Bipotential mouse embryonic liver stem cell lines contribute to liver regeneration and differentiate as bile ducts and hepatocytes. Proc Natl Acad Sci U S A 101(22): 8360-8365.
Strick-Marchand, H. and Weiss, M.C. 2002. Inducible differentiation and morphogenesis of bipotential liver cell lines from wild-type mouse embryos. Hepatology 36(4 Pt 1): 794-804.

Chapter 1: Introduction 49
Stringer, K.F., Ingles, C.J., and Greenblatt, J. 1990. Direct and selective binding of an
acidic transcriptional activation domain to the TATA-box factor TFIID. Nature 345(6278): 783-786.
Tanimizu, N. and Miyajima, A. 2004. Notch signaling controls hepatoblast differentiation by altering the expression of liver-enriched transcription factors. J Cell Sci 117(Pt 15): 3165-3174.
Tanimizu, N., Nishikawa, M., Saito, H., Tsujimura, T., and Miyajima, A. 2003. Isolation of hepatoblasts based on the expression of Dlk/Pref-1. J Cell Sci 116(Pt 9): 1775-1786.
Tanimizu, N., Saito, H., Mostov, K., and Miyajima, A. 2004. Long-term culture of hepatic progenitors derived from mouse Dlk+ hepatoblasts. J Cell Sci 117(Pt 26): 6425-6434.
Tatarakis, A., Margaritis, T., Martinez-Jimenez, C.P., Kouskouti, A., Mohan, W.S., 2nd, Haroniti, A., Kafetzopoulos, D., Tora, L., and Talianidis, I. 2008. Dominant and redundant functions of TFIID involved in the regulation of hepatic genes. Mol Cell 31(4): 531-543.
Taura, D., Noguchi, M., Sone, M., Hosoda, K., Mori, E., Okada, Y., Takahashi, K., Homma, K., Oyamada, N., Inuzuka, M. et al. 2009. Adipogenic differentiation of human induced pluripotent stem cells: comparison with that of human embryonic stem cells. FEBS Lett 583(6): 1029-1033.
Thomas, M.C. and Chiang, C.M. 2006. The general transcription machinery and general cofactors. Crit Rev Biochem Mol Biol 41(3): 105-178.
Thompson, N.L., Mead, J.E., Braun, L., Goyette, M., Shank, P.R., and Fausto, N. 1986. Sequential protooncogene expression during rat liver regeneration. Cancer Res 46(6): 3111-3117.
Thut, C.J., Chen, J.L., Klemm, R., and Tjian, R. 1995. p53 transcriptional activation mediated by coactivators TAFII40 and TAFII60. Science 267(5194): 100-104.

Chapter 1: Introduction 50
Tremblay, K.D. and Zaret, K.S. 2005. Distinct populations of endoderm cells converge to
generate the embryonic liver bud and ventral foregut tissues. Dev Biol 280(1): 87-99.
Veenstra, G.J., Weeks, D.L., and Wolffe, A.P. 2000. Distinct roles for TBP and TBP-like factor in early embryonic gene transcription in Xenopus. Science 290(5500): 2312-2315.
Vermeulen, M., Mulder, K.W., Denissov, S., Pijnappel, W.W., van Schaik, F.M., Varier, R.A., Baltissen, M.P., Stunnenberg, H.G., Mann, M., and Timmers, H.T. 2007. Selective anchoring of TFIID to nucleosomes by trimethylation of histone H3 lysine 4. Cell 131(1): 58-69.
Verrijzer, C.P., Chen, J.L., Yokomori, K., and Tjian, R. 1995. Binding of TAFs to core elements directs promoter selectivity by RNA polymerase II. Cell 81(7): 1115-1125.
Voss, A.K., Thomas, T., Petrou, P., Anastassiadis, K., Scholer, H., and Gruss, P. 2000. Taube nuss is a novel gene essential for the survival of pluripotent cells of early mouse embryos. Development 127(24): 5449-5461.
Wandzioch, E. and Zaret, K.S. 2009. Dynamic signaling network for the specification of embryonic pancreas and liver progenitors. Science 324(5935): 1707-1710.
Wang, J., Clark, J.B., Rhee, G.S., Fair, J.H., Reid, L.M., and Gerber, D.A. 2003a. Proliferation and hepatic differentiation of adult-derived progenitor cells. Cells Tissues Organs 173(4): 193-203.
Wang, N.D., Finegold, M.J., Bradley, A., Ou, C.N., Abdelsayed, S.V., Wilde, M.D., Taylor, L.R., Wilson, D.R., and Darlington, G.J. 1995. Impaired energy homeostasis in C/EBP alpha knockout mice. Science 269(5227): 1108-1112.
Wang, X., Foster, M., Al-Dhalimy, M., Lagasse, E., Finegold, M., and Grompe, M. 2003b. The origin and liver repopulating capacity of murine oval cells. Proc Natl Acad Sci U S A 100 Suppl 1: 11881-11888.

Chapter 1: Introduction 51
Wang, X., Quail, E., Hung, N.J., Tan, Y., Ye, H., and Costa, R.H. 2001. Increased levels
of forkhead box M1B transcription factor in transgenic mouse hepatocytes prevent age-related proliferation defects in regenerating liver. Proc Natl Acad Sci U S A 98(20): 11468-11473.
Wassarman, D.A., Aoyagi, N., Pile, L.A., and Schlag, E.M. 2000. TAF250 is required for multiple developmental events in Drosophila. Proc Natl Acad Sci U S A 97(3): 1154-1159.
Wassarman, D.A. and Sauer, F. 2001. TAF(II)250: a transcription toolbox. J Cell Sci 114(Pt 16): 2895-2902.
Wauthier, E., Schmelzer, E., Turner, W., Zhang, L., LeCluyse, E., Ruiz, J., Turner, R., Furth, M.E., Kubota, H., Lozoya, O. et al. 2008. Hepatic stem cells and hepatoblasts: identification, isolation, and ex vivo maintenance. Methods Cell Biol 86: 137-225.
Webber, E.M., Bruix, J., Pierce, R.H., and Fausto, N. 1998. Tumor necrosis factor primes hepatocytes for DNA replication in the rat. Hepatology 28(5): 1226-1234.
Webber, E.M., FitzGerald, M.J., Brown, P.I., Bartlett, M.H., and Fausto, N. 1993. Transforming growth factor-alpha expression during liver regeneration after partial hepatectomy and toxic injury, and potential interactions between transforming growth factor-alpha and hepatocyte growth factor. Hepatology 18(6): 1422-1431.
Weinzierl, R.O., Dynlacht, B.D., and Tjian, R. 1993a. Largest subunit of Drosophila transcription factor IID directs assembly of a complex containing TBP and a coactivator. Nature 362(6420): 511-517.
Weinzierl, R.O., Ruppert, S., Dynlacht, B.D., Tanese, N., and Tjian, R. 1993b. Cloning and expression of Drosophila TAFII60 and human TAFII70 reveal conserved interactions with other subunits of TFIID. EMBO J 12(13): 5303-5309.
Wells, J.M. and Melton, D.A. 2000. Early mouse endoderm is patterned by soluble factors from adjacent germ layers. Development 127(8): 1563-1572.

Chapter 1: Introduction 52
Wieczorek, E., Brand, M., Jacq, X., and Tora, L. 1998. Function of TAF(II)-containing
complex without TBP in transcription by RNA polymerase II. Nature 393(6681): 187-191.
Wilson, J.W. and Leduc, E.H. 1958. Role of cholangioles in restoration of the liver of the mouse after dietary injury. J Pathol Bacteriol 76(2): 441-449.
Wright, K.J., Marr, M.T., 2nd, and Tjian, R. 2006. TAF4 nucleates a core subcomplex of TFIID and mediates activated transcription from a TATA-less promoter. Proc Natl Acad Sci U S A 103(33): 12347-12352.
Wright, K.J. and Tjian, R. 2009. Wnt signaling targets ETO coactivation domain of TAF4/TFIID in vivo. Proc Natl Acad Sci U S A 106(1): 55-60.
Xiao, L., Kim, M., and DeJong, J. 2006. Developmental and cell type-specific regulation of core promoter transcription factors in germ cells of frogs and mice. Gene Expr Patterns 6(4): 409-419.
Yamanaka, S. 2007. Strategies and new developments in the generation of patient-specific pluripotent stem cells. Cell Stem Cell 1(1): 39-49.
Yang, Y., Cao, J., Huang, L., Fang, H.Y., and Sheng, H.Z. 2006. Regulated expression of TATA-binding protein-related factor 3 (TRF3) during early embryogenesis. Cell Res 16(7): 610-621.
Yaswen, P., Hayner, N.T., and Fausto, N. 1984. Isolation of oval cells by centrifugal elutriation and comparison with other cell types purified from normal and preneoplastic livers. Cancer Res 44(1): 324-331.
Ye, H., Holterman, A.X., Yoo, K.W., Franks, R.R., and Costa, R.H. 1999. Premature expression of the winged helix transcription factor HFH-11B in regenerating mouse liver accelerates hepatocyte entry into S phase. Mol Cell Biol 19(12): 8570-8580.
Zaret, K.S. 2002. Regulatory phases of early liver development: paradigms of organogenesis. Nat Rev Genet 3(7): 499-512.

Chapter 1: Introduction 53
Zaret, K.S. and Grompe, M. 2008. Generation and regeneration of cells of the liver and
pancreas. Science 322(5907): 1490-1494.
Zhang, D., Penttila, T.L., Morris, P.L., Teichmann, M., and Roeder, R.G. 2001. Spermiogenesis deficiency in mice lacking the Trf2 gene. Science 292(5519): 1153-1155.
Zhang, J., Wilson, G.F., Soerens, A.G., Koonce, C.H., Yu, J., Palecek, S.P., Thomson, J.A., and Kamp, T.J. 2009. Functional cardiomyocytes derived from human induced pluripotent stem cells. Circ Res 104(4): e30-41.
Zhao, R., Watt, A.J., Li, J., Luebke-Wheeler, J., Morrisey, E.E., and Duncan, S.A. 2005. GATA6 is essential for embryonic development of the liver but dispensable for early heart formation. Mol Cell Biol 25(7): 2622-2631.
Zhou, J., Zwicker, J., Szymanski, P., Levine, M., and Tjian, R. 1998. TAFII mutations disrupt Dorsal activation in the Drosophila embryo. Proc Natl Acad Sci U S A 95(23): 13483-13488.

Chapter 2: TBP, TRFs, and TAFs in Liver Development and Regeneration
Chapter 2: TBP, TRFs, and TAFs in Liver Development and Regeneration Summary
The precise spatial and temporal control of gene expression required for
organismal development and cellular differentiation is primarily determined at the level
of transcriptional regulation. Historically, diverse cellular programs of proliferation,
terminal differentiation, and specialized function have been attributed to a repertoire of
sequence-specific activators and repressors. General transcription factors including
coactivators such as Mediator and core promoter recognition factors such as TFIID were
assumed to be ubiquitous and invariant between tissues and developmental time points.
Recent work from a variety of model organisms and mammalian differentiation programs
supports an expanding role for TAF paralogs, TBP-related factors, and alternative core
promoter recognition complexes in determining tissue and developmental stage-specific
transcriptional programs. The vertebrate liver serves varied and critical functions in
metabolism, growth control, and detoxification; however, the precise transcriptional
mechanisms of hepatic development, differentiation, and regeneration remain elusive.
Herein, I examine the expression of TBP, TRFs, TAFs and other coactivators in adult
mouse tissues and purified liver cells. Furthermore, I demonstrate that during liver
development and hepatocyte commitment canonical TFIID is in toto eliminated and its
gene locus transcriptionally silenced coincident with retention of TRF3 expression.
Importantly, an in vitro model of hepatic development recapitulates these observations.

Chapter 2: TBP, TRFs, and TAFs in Liver Development and Regeneration
Introduction
The highly specific and exquisitely complex patterns of gene expression required
for organismal development and cellular differentiation have traditionally been attributed
to sequence-specific DNA binding activators, while the general transcription factors
including the core promoter recognition complex TFIID and the coactivator Mediator
were assumed to be invariant (Thomas and Chiang 2006). In the mammalian liver
developmental commitment and functional specialization are primarily attributed to a
large group of heterogeneous nuclear factors (HNFs) and other liver-enriched activators,
while changes in the basal machinery have remained largely uninvestigated (Schrem et
al. 2002). Recent advances in genetic, biochemical, and cell culture techniques have
allowed for a more thorough examination of core promoter recognition and coactivator
proteins in diverse cell types and differentiation programs.
The first indication that the canonical core promoter recognition complex TFIID
may not be essential at all developmental stages or for the transcription of all genes came
from multiple gene disruption studies. Hence, while depletion of TBP in the mouse or
zebrafish leads to eventual embryonic lethality it does not prevent the onset of zygotic
transcription or reduce RNA PolII activity at the earliest embryonic stages, suggesting
that an alternative mechanism of core promoter induction must take its place in this
particular setting (Muller et al. 2001; Martianov et al. 2002). Similarly, disruption of the
TBP associated factors TAF10 and TAF8 also led to eventual lethality but only after
expansion of the inner cell mass, and staining experiments suggest differing requirements
for these factors in the developing blastocyst (Voss et al. 2000; Mohan et al. 2003).

Chapter 2: TBP, TRFs, and TAFs in Liver Development and Regeneration
Conditional inactivation of the key TFIID subunit, TAF4 in embryonic fibroblasts
showed it to be nonessential for cellular growth and differentiation and required for the
transcription of many but not all fibroblastic genes, while depletion in developing
keratinocytes suggested that it is essential to epidermal differentiation and maintenance,
consistent with differential tissue-specific functions (Mengus et al. 2005). Intriguingly,
TAF10 is also required for embryonic keratinocyte development but unlike TAF4 is
dispensable for maintenance of the adult epidermis (Indra et al. 2005). Most recently,
TAF10 was shown to be required for embryonic liver development but dispensable for
the maintenance of adult hepatocytes and the ongoing expression of most liver-specific
genes, suggesting that an alternative mechanism of core promoter activation may replace
TFIID in the hepatogenic program (Tatarakis et al. 2008).
Additional evidence for tissue-specific differences in TFIID activity comes from
several limited observations of differences in TBP and TAF expression throughout adult
tissues. Hence, in an early study TBP and TAF12 expression were shown to vary widely
between adult mouse tissue, being highly enriched in the testes and nearly undetectable in
the heart and nervous system (Perletti et al. 1999). Furthermore, differences in TBP
abundance were inversely correlated with TRF levels suggesting that TRFs may partially
replace TBP in certain tissues. In another report, TAF4 was shown to be highly
upregulated during neuronal differentiation and required for the induction of adult
neuron-specific genes (Perletti et al. 1999). Furthermore, increased TAF8 (TBN)
expression was shown to occur during adipogenesis, and functional TAF8 shown to be
required for induction of the adipogenic gene expression program (Perletti et al. 1999).

Chapter 2: TBP, TRFs, and TAFs in Liver Development and Regeneration
Additionally, an investigation of the mouse TAF4 paralogue, TAF4b, demonstrated that
TAF4 is significantly donwregulated in the adult liver at least at the mRNA level
(Freiman et al. 2001). While it is important to note that many of these studies are
observational and may depend on the accuracy of specific reagents, collectively they
provide a precedent for the idea that significant differences do exist in the expression of
TFIID components between tissues and cell types.
The most recent indication of developmental stage-specific changes in the core
promoter recognition machinery occurs in the myogenic program. Hence, in both
primary cells and an in vitro model of myogenesis TBP and many TAFs are in toto
degraded and their mRNA levels significantly decreased during the transition from
myoblasts to myotubes (Deato and Tjian 2007). Importantly, levels of other general
transcription factors including TFIIA-H and RNA PolII are unchanged as are levels of
TAF3 and the TBP-related factor TRF3. Furthermore, TFIID was shown in vitro to be
dispensable for the activated transcription of the myoube-specific myogenin promoter,
suggesting that TBP and TAFs are at least in part functionally nonessential for the onset
and/or maintenance of transcription in the adult cell type (Deato et al. 2008).
Additionally, multiple components of the canonical coactivator complex mediator are
also significantly donwregulated during the myogenic transition, suggesting that like
TFIID, Med activity may vary significantly between developmental stages and tissues.
Importantly, downregulation of TFIID does not appear to be a hallmark of all
developmental transitions, as TBP and TAF levels are shown to be unaltered at both the

Chapter 2: TBP, TRFs, and TAFs in Liver Development and Regeneration
protein and mRNA level during the differentiation of mouse ES cells to motor neurons
(Francisco Herrera, personal communication).
Results
Varied Expression of TFIID Subunits in Diverse Mouse Tissues
The recent seminal finding that TFIID including TBP and many TAF subunits is
wholly degraded and replaced by a novel core promoter recognition complex consisting
of TRF3 and TAF3 during terminal differentiation has thus far been restricted to murine
muscle development (Deato and Tjian 2007). To better understand if this intriguing
transcriptional mechanism is unique to myogenesis or is a common feature of other
distinct differentiation programs we investigated the disposition of TBP and multiple
TAFs in diverse adult mouse tissues at both the mRNA and protein level.
To determine the relative levels of TBP and TAF protein in differentiated mouse
tissues we subjected whole tissue lystates to Western blot analysis with specific
antibodies against TBP, TRF3, TAF1, and TAF4 (Figure 1). Equal amounts of total
protein were analyzed and GAPDH was used as a loading control. Importantly, TBP and
multiple TAFs are enriched in proliferative C2C12 myoblasts and whole E13.5 embryos
which contain multiple progenitor cell types including myoblasts and hepatoblasts, but at
significantly reduced levels in differentiated C2C12 myotubes as previously reported.
Additionally, TFIID components are highly expressed in the testes and spleen which
poses significant stem and progenitor cell populations and were formerly shown to
contain high levels of TBP (Persengiev et al. 1996; Perletti et al. 1999). Intriguingly,

Chapter 2: TBP, TRFs, and TAFs in Liver Development and Regeneration
TBP, TAF1, and TAF4 protein appear to be all but absent from many adult tissues and at
exceedingly low levels in the whole liver; possibly at lower levels than in differentiated
myotubes. Conversely, TRF3 protein is as abundant in many adult tissues as it is in
C2C12 myoblasts or the whole embryo, and appears to be particularly abundant in the
liver sample. Hence, core promoter recognition complex protein levels vary widely
between tissues and TFIID protein levels appear to be particularly low in the
differentiated mouse liver.
To further investigate the tissue-specific expression of TFIID we determined the
relative levels of key subunit transcripts in the same tissues by reverse transcription and
quantitative real-time PCR (qPCR) (Figure 2). Because no single mRNA is expressed at
the same level in all tissues of the mammalian body, and the relative proportion of
mRNA to total RNA varies considerably between tissues, accurate normalization of cross
tissue qPCR is inherently problematic. To partially abrogate this problem, we used the
same quantity of total RNA in reverse transcription reactions and normalized against both
the 18s RNA (Figure 1A) and GAPDH transcript (Figure 2B), two of the most widely
accepted total RNA and mRNA standards respectively. As with protein levels,
significant differences are observed in TFIID mRNA levels between tissues, with adult
liver showing some of the lowest levels of both TBP and multiple TAFs. Importantly,
the whole embryo, testes and ovary, which contain more progenitors and proliferative
cells than most adult tissues have the highest transcript levels of key TFIID components.
While significant differences exist for TBP and TAF levels between the GAPDH and 18s
normalizations, the general trend of reduced TFIID expression in the liver is the same.

Chapter 2: TBP, TRFs, and TAFs in Liver Development and Regeneration
As with the protein analysis, levels of TRF3 transcript do not correlate with TBP or TAFs
and are at least as high in the liver and other adult tissues as the embryo or testes.
Differential Expression of TBP, TRFs and TAFs in Highly Purified Liver Cells
The embryonic and adult livers both contain multiple highly specialized cell types
with unique functions and transcriptional programs (Zaret 2002; Zaret and Grompe
2008). While hepatocytes comprise sixty to eighty percent of the adult liver mass
significant populations of cholangiocytes and Kupffer cells, and smaller populations of
several other specialized cell types are present and serve critical physiological functions.
Similarly, the bipotential hepatoblasts that eventually give rise to hepatocytes and
cholangiocytes are a minority population in the developing liver bud, where erythrocytes,
leukocytes and other hematopoietic cells are highly abundant. Consistent with their
specialized functions and gene expression patterns each of these cell types maintains a
unique transcriptional program (Cereghini 1996; Costa et al. 2003). Hence, analysis of
core promoter recognition complex subunits in whole liver tissue is likely complicated by
differences in expression between the unique cell types that comprise the tissue mass, and
may represent an average of different expression levels between the various cell types.
More generally, most differentiated tissues are composed of multiple highly specialized
cell types in addition to progenitors, such that any whole tissue expression analysis likely
misrepresents the abundance of a given gene product in a specific functional cell.
To overcome this complication we chose to focus our analysis on highly purified
embryonic and adult liver cells. Multiple protocols have been established for the

Chapter 2: TBP, TRFs, and TAFs in Liver Development and Regeneration
purification of hepatoblasts from the embryonic liver both by enrichment of hepatoblast-
specific markers and depletion of hematopoietic cells (Weiss and Strick-Marchand 2003;
Wauthier et al. 2008). We found that collagenase based dissociation of individually
dissected E13.5 livers followed by magnetic bead based depletion of erythrocytes
(TER119) and hematopoietic cells (CD45) consistently produced hepatoblast populations
which were ninety five percent pure based on both negative and positive FACS analysis
(Suzuki et al. 2000). Protocols for the purification of neonatal, partial hepatectomy, and
adult hepatocytes by two step in situ perfusion with collagenase and differential
centrifugation have been employed for more than 50 years (Wang et al. 2003a). The
most excepted protocol for purification of regenerative cycling hepatocyte is 2/3 partial
hepatectomy followed by in situ perfusion (Mitchell and Willenbring 2008).
To determine the expression of TBP, TRFs, and TAFs we performed quantitative
RT-PCR using equal amounts of RNA from purified hepatoblasts and adult hepatocytes
and normalized to the GAPDH transcript (Figure 3A). Intriguingly TBP, TRF2 and the
majority of TAFs are significantly downregulated at the mRNA level in hepatocytes
when compared to hepatoblasts. Importantly, his includes the donwregulation of TAF3
which was previously shown to be unaltered in the myogenic differentiation program
(Deato and Tjian 2007). Conversely, TRF3 and TAF13 expression appear to be
relatively unchanged in the adult hepatocyte while the testes-specific TAF7 paralogue,
TAF7l is significantly unregulated. Additionally, expression of a core component of a
TBP-free TAF complex, PCAF is induced in hepatocytes.

Chapter 2: TBP, TRFs, and TAFs in Liver Development and Regeneration
The recent finding that components of the coactivator complex Mediator are
downregulated during myogenesis led us to ask how mediator is affected in the hepatic
program (Deato et al. 2008). Hence, we analyzed the expression of several key Mediator
subunits including components of the head, middle, tail, and CDK8 domains in
hepatoblast and hepatocytes by qPCR (Figure 3B). As with TFIID each mediator mRNA
examined is significantly downregulated in hepatocytes relative to hepatoblasts. While
approximately half of the known Mediator subunits were excluded in this analysis, none
of these proteins has been yet been isolated in a complex other than canonical Mediator
and those subunits which are shown to be downregulated are positioned throughout the
known complex (Myers and Kornberg 2000; Taatjes et al. 2004).
These analyses demonstrate a clear difference in the expression of TFIID and
Mediator subunits between progenitor hepatoblasts and adult hepatocytes. This led us to
ask when in hepatic development TFIID is downregulated and how its expression might
be altered following liver injury. While hepatocytes start to develop from hepatoblasts as
early as embryonic day 14, they may not become fully differentiated until postnatal day
21. Hence, neonatal hepatocytes are smaller and less binucleate than adult hepatocytes,
do not express all adult hepatic genes, and continue to proliferate. While not fully
understood neonatal hepatocyte maturation likely involves transcriptional changes which
are distinct from the embryonic or adult programs. Following toxic or mechanical injury
adult hepatocytes reenter the cell cycle and proliferate to regenerate the liver mass.
These regenerative hepatocytes undergo significant changes in gene expression without
dedifferentiation and are thus a unique cell type from either quiescent hepatocytes or

Chapter 2: TBP, TRFs, and TAFs in Liver Development and Regeneration
embryonic and neonatal progenitors. While liver-enriched transcription factors and
signaling molecules are known to be upregulated during liver injury, the transcriptional
programs which govern hepatocyte proliferation are not well understood. Thus, we
compared TBP, TRF, TAF, and Mediator mRNA levels from neonatal hepatocytes and
2/3 partial hepetectomy hepatocytes to hepatoblasts and adult hepatocytes by qPCR to
further understand the role of these complexes in maturation and regeneration (Figure 4).
Based on this analysis it appears the TFIID and Med expression in neonatal and
regenerative hepatocytes mirrors that of adult hepatocytes as TBP and most TAFs are
similarly downregulated relative to hepatocytes while TRF3, TAF13, and TAF7l are
unchanged. Hence, canonical TFIID and Mediator are likely downregulated and TAF7l
upregulated at the early stages of hepatocyte commitment prior to birth and their
expression does not appear to be significantly reinitiated following liver injury and
regeneration.
While spatial and temporal patterns of gene expression as determined by analysis
of mRNA abundance often correlate with developmental and cell-type-specific function,
differences in mRNA stability, translational efficiency, and protein turnover may cause
significant fluctuations between mRNA levels and protein abundance. For this reason
relative polypeptide levels are generally assumed to more accurately reflect the
developmental stage and cell-type-specific requirements for a given protein product.
Hence, we examined TBP, TRF and critical TAF subunit protein levels by western blot of
whole fetal, adult liver, and purified liver cells (Figure 5). This analysis demonstrates
that TBP and multiple TAFs are highly enriched in purified hepatoblasts and are possibly

Chapter 2: TBP, TRFs, and TAFs in Liver Development and Regeneration
expressed at higher levels than in C2C12 myoblasts or whole fetal liver. Conversely,
TBP and every TAF examined is significantly downregulated in adult hepatocytes when
compared to hepatoblasts including the critical structural subunits TAF1, TAF4 and
TAF5 (Wright et al. 2006). Furthermore, neonatal and regenerative hepatocytes appear
to have more TBP and TAF protein than do purified adult hepatocytes but clearly far less
than hepatoblasts. Intriguingly, several TAFs such as TAF10 appear to be more highly
expressed in the whole adult liver than they are in purified hepatocytes suggesting liver
cells other than hepatocytes may posses higher TAF levels. Additionally, protein levels
of TRF2 appear to be relatively unchanged between hepatoblasts and hepatocytes,
suggesting that this protein may partially substitute for TBP in adult hepatocytes. In the
case of TBP and several of the TAFs the fold reduction in protein levels may be greater
than that for mRNA, but this comparison is limited by the qualitative nature of western
blotting.
To further understand changes in Mediator expression during hepatic
development we also conducted western blots of multiple Med subunits in whole liver
and purified liver cells (Figure 6). As is the case with TFIID, every Mediator subunit
examined is enriched in fetal hepatoblasts and significantly downregulated in adult
hepatocytes while at reduced but intermediate levels in neonatal and regenerative
hepatocytes. Importantly, this is true for components of the distinct head, middle, and tail
domains, and the dissociable CDK8 module (Guglielmi et al. 2004). The apparent
donwregulation of Mediator protein during hepatic development closely mirrors what
was reported for the myogenic program (Deato et al. 2008).

Chapter 2: TBP, TRFs, and TAFs in Liver Development and Regeneration
Transcriptional Silencing of TBP and TAF Promoters During Hepatogenesis
While qPCR and western blot analysis clearly show that TFIID levels are
significantly reduced upon hepatic development, we wished to further confirm that TBP
and TAF gene expression is specifically silenced at the promoter level. The enrichment
or removal of highly defined histone marks is increasingly understood to correlate with
the level or gene expression or silencing (Bernstein et al. 2007). Among the many
recently identified histone modifications, promoter enrichment of histone H3 lysine 4
(H3K4Me3) trimethylation has been widely shown to correlate with gene activation,
while enrichment of histone H3 lysine 9 trimethylation (H3K9Me3) is associated with
transcriptional silencing (Li et al. 2007). Hence, we crosslinked E13.5 and adult livers
and performed chromatin immunoprecipitation with specific antibodies against these two
chromatin marks followed by qPCR of precipitated material with promoter-specific
primers (Figure 7). This analysis shows that the proximal promoters of TBP, TAF3, and
TAF5 are enriched for H3K4Me3 in the fetal liver sample and that levels of this mark are
subsequently reduced in the adult liver. Similarly and more strikingly, H3K9Me3
enrichment at these promoters is low in the fetal liver and greatly increased in the adult.
Importantly, the promoter of an adult hepatocyte-specific gene, CYP7A1, shows relative
enrichment of H3K9Me3 in the fetal liver and of H3K4Me3 in the adult. Hence, TFIID
promoters appear to be specifically upregulated in the fetal liver and transcriptionally
silenced in adult cells through a mechanism which involves directed modification of
histone tails in the proximal promoters of these genes.

Chapter 2: TBP, TRFs, and TAFs in Liver Development and Regeneration
Expression of TBP and TAFs in a Cell Culture Model of Hepatogenesis
Defined cell culture systems have contributed greatly to our understanding of
multiple metazoan differentiation programs and provide the greatest opportunity for
functional manipulation of cellular differentiation when a suitable whole animal model is
absent. Several groups have reported systems for both the short term ex vivo culture of
primary hepatoblasts and the long term maintenance of immortalized hepatoblast derived
cell lines (Strick-Marchand and Weiss 2003; Weiss and Strick-Marchand 2003). Hence,
fetal mouse hepatoblasts purified by both negative and positive selection have been
successfully cultured for multiple divisions on extracellular matrices in the presence of
growth factors which mimic the hepatic endoderm niche (Nitou et al. 2002; Tanimizu et
al. 2003). Furthermore, these cultures have been efficiently differentiated by defined
factors into both hepatocytes and cholangiocytes that recapitulate the metabolic functions
and gene expression patterns of in vivo isolates (Kamiya et al. 1999; Kinoshita et al.
1999; Tanimizu et al. 2007). While these ex vivo models have proven useful they require
frequent isolation of fresh hepatoblasts and result in significant experimental variability.
To date three groups have established defined immortalized hepatoblast cell lines which
abrogate the complexities of hepatoblast culture while faithfully recapitulating embryonic
liver development (Rogler 1997; Strick-Marchand and Weiss 2002; Tanimizu et al.
2004). Of these we chose to employ the Hepatic Progenitors Proliferating on Laminin
(HPPL) cell line, which mimics proliferative hepatoblasts in the presence of epidermal
growth factor (EGF) and hepatocyte growth factor (HGF) but is differentiated to
functional hepatocytes following incubation with oncostatin M (OSM) and Matrigel.

Chapter 2: TBP, TRFs, and TAFs in Liver Development and Regeneration
We maintained and expanded the HPPL cell line for multiple generations on
laminin-coated plates in the presence of EGF and HGF with no apparent change in
morphology, proliferative capacity, or marker gene expression consistent with published
reports. When confluent cultures were incubated for five days with 20ng/ml of
recombinant oncostatin M followed by five days with growth factor reduced Matrigel
cells took on an enlarged binucleate cuboidal morphology and acquired multiple
intracellular vacuolar structures. Reverse transcription and qPCR revealed a significant
decrease in the expression of bipotential and cholangiocytes markers by five days post
induction and a greater increase by ten days (Figure 8A). Conversely, adult hepatic gene
expression was significantly and rapidly upregulated. Similarly, western blot analysis
showed a reduction of bipotential marker protein levels and an increase in proteins
associated with terminal adult hepatocytes (Figure 8B). Thus the HPPL cell model
faithfully mimics the hepatoblast to hepatocyte transition based on morphology and
marker gene expression.
To assess changes in TBP and TAF expression during HPPL differentiation we
performed reverse transcription and qPCR on RNA isolated from proliferating and
differentiated HPPL cells (Figure 9A). These analyses clearly show that TBP, many
TAFs, and at least one Mediator subunit are significantly downregulated by five days
post induction with further downregulation by ten days. Conversely, TRF3, TAF13, and
TAF7l transcript levels are maintained or increased coincident with the downregulation
of other subunits. Importantly the extent of transcript changes closely mirrors that seen
between primary hepatoblasts and hepatocytes. Western blot analysis of TBP, TRF3, and

Chapter 2: TBP, TRFs, and TAFs in Liver Development and Regeneration
some TAFs displays a similar pattern with significant changes in TFIID but not TRF3
occurring by day five after induction. Hence, the HPPL culture system faithfully
recapitulates the downregulation of TFIID observed the primary hepatogenic program.
Discussion
A growing body of functional data suggests that specific TAFs and potentially holo-
TFIID may be dispensable for the activation of many promoters and the maintenance of
at least some differentiated cell types. This, combined with two recent studies that
demonstrate that TBP and multiple TAFs are specifically degraded during certain
differentiation programs, cast doubt on the idea that TFIID is universally expressed and
required for all cell-type-specific transcriptional programs (Brunkhorst et al. 2005; Deato
and Tjian 2007). Because of its central role in mammalian biology, the transcriptional
mechanisms that govern liver development and function are a subject of extensive study.
At least one previous study suggested that the adult mouse liver contains
particularly low levels of the key TFIID subunit TAF4 (Freiman et al. 2001). Indeed our
current data confirms that TAF4, along with TBP and TAF1, are significantly
downregulated in the adult liver relative to the embryo and other adult tissues, suggesting
that the hepatogenic program may also require the downregulation of canonical TFIID
and potentially its replacement with an alternative core promoter recognition complex.
While methods of normalization and differences in progenitor cell populations between
tissues complicate cross tissue analysis, preliminary data suggest that the significant
downregulation of TFIID at both the protein and RNA level that is observed in muscle

Chapter 2: TBP, TRFs, and TAFs in Liver Development and Regeneration
and liver may not be observed in all adult tissues and thus may be specific to a subset of
differentiation programs. As new protocols for the isolation of tissue progenitors and
fully committed cells become available, it will be interesting to explore how widely the
phenomenon of TFIID downregulation and core promoter complex switching is
employed or restricted throughout the mammalian body.
Because of the considerable cell type and functional complexity of most adult
organs, changes in transcription factor expression and function are clearly best observed
in highly defined pure cell populations. Hence, hepatocytes which represent sixty to
seventy percent of the adult liver cell population may exhibit differences in transcription
factor abundance from other liver cell types which would be masked in whole organ
experiments (Daoust and Cantero 1959). Importantly, our results suggest that the
hepatogenic downregulation of TFIID at both the protein and mRNA levels is most
pronounced between hepatoblast progenitors and purified hepatocytes. Intriguingly,
whole adult livers appear to retain at least some TAF subunits relative to hepatocytes,
consistent with previous literature reports (Tatarakis et al. 2008). While the many minor
cell types of the adult and developing liver are more difficult to purify and characterize, it
will be interesting for future studies to determine if at least some of these populations
retain and require canonical TFIID in their adult form, and if this accounts for at least
part of the difference in TFIID abundance between whole liver and purified hepatocytes
(Alpini et al. 1994). Specifically, it remains to be determined if bipotential hepatoblasts
downregulate TFIID when committing to both mature hepatocytes and cholangiocytes or
if selective downregulation is itself a determinant of this lineage split. While clear

Chapter 2: TBP, TRFs, and TAFs in Liver Development and Regeneration
differences in TFIID expression exist between hepatocytes and hepatoblasts, these two
cell types lie at the ends of a differentiation process which requires at least two weeks and
multiple intermediate stages in the mouse, making the exact timing of TFIID
downregulation unclear (Zaret 2002). The finding that TAFs are already significantly
degraded in neonatal hepatocytes suggests that this process primarily occurs early in
differentiation and most likely prenatally, however the observation that some protein
remains in late neonatal hepatocytes may imply that at least some TFIID is required for
final maturation. Additionally, differences in the relative levels of TAFs in purified
hepatocytes and the whole liver may indicate that some subunits are retained as part of
characterized or novel TBP free TAF containing complexes including TFTC and SAGA;
the role of these complexes in liver development and function remains to be determined.
The significant decrease in TBP and TAF mRNA levels and the active silencing of these
genes promoters supports the notion that long term reduction of TFIID in adult
hepatocytes is primarily a transcriptional phenomenon. However, more extensive time
course measurements of mRNA and protein levels during the differentiation process is
needed to determine if an active mechanism of subunit proteolysis is also employed. One
recent study suggests that in some differentiation programs, targeted ubiquitination and
proteasome mediated degradation of TFIID subunits occurs independently of
transcriptional changes (Perletti et al. 2001). Thus, it is possible that both transcriptional
silencing and ubiquitin E3 ligase mediated degradation of specific TFIID polypeptides
cooperate to establish immediate and prolonged reduction of TFIID levels. Most
interestingly, it remains to be understood how known and as yet undiscovered

Chapter 2: TBP, TRFs, and TAFs in Liver Development and Regeneration
extracellular signaling events trigger the transcriptional silencing of TBP and TAF
promoters or the targeted degradation of these subunits in precise spatial and temporal
patterns. Recent advances in the ex vivo study of these critical signaling events may be
fruitfully combined with existing transcriptional readouts to better understand these
connections (Wandzioch and Zaret 2009).
Importantly, a cell culture model of hepatic differentiation recapitulates the
downregulation of TFIID observed in purified primary cells. This model expands the
range of functional and cell signaling experiments that can be performed to better
understand the process of core promoter complex switching in hepatogenesis.
Additionally, these observations also suggest that because core promoter changes that are
observed in vivo are conserved in vitro, novel findings may be successfully employed to
improve the efficiency and fidelity of potentially therapeutic differentiation and
transplantation protocols.
Materials and Methods
Protein Extracts and RNA
Whole tissue lysates were generated by dounce homogenization in RIPA buffer of
fresh CD-1 mouse tissues dissected in cold PBS, or purchased from Protientech Group.
Tissue-specific total RNA was purchased from Ambion and used to generate cDNA by
reverse transcription with the High Capacity cDNA Kit (ABI). Purified mouse liver cells
were homogenized in RIPA buffer or used to generate cDNA by extraction with the
RNeasy Plus Mini Kit (Qiagen) and reverse transcription with the High Capacity cDNA

Chapter 2: TBP, TRFs, and TAFs in Liver Development and Regeneration
Kit. Protein samples were clarified by centrifugation and total protein concentration
normalized to BSA with the BCA Protein Assay reagents (Pierce).
Quantitative PCR
Relative qPCR of total cDNA was performed on an ABI 7300 with transcript
specific FAM labeled Taqman probes and Universal Master Mix (ABI) or verified
transcript specific primer sets and Power SYBR Green Master Mix (ABI). Input was
normalized to 18s for whole tissue samples and GAPDH for all other experiments.
Samples were quantified in triplicate and represent a minimum of two biological
replicates.
Western Blotting
Equal amounts of total protein were separated by denaturing polyacryalamide gel
electrophoresis and transferred to 0.22μm nitrocellulose, blocked with 5% milk or 5%
BSA in Tris-buffered saline/0.1% Tween (TBST) and incubated overnight with
appropriate antibodies. Membranes were washed extensively with TBST and probed
with species-specific horseradish peroxidase conjugated secondary antibodies (Jackson).
The following Antibodies were used: Rabbit anti-MED1 (Bethyl), Rabbit anti-MED6
(Bethyl), Rabbit anti-MED12 (Novus), Rabbit anti-MED18 (Novus), Rabbit anti-MED23
(Bethyl), Goat anti-CDK8 (Abcam), Mouse anti-TBP (Biodesign), Rabbit anti-TRF2
(Abcam), Rabbit anti-TRF3 (Deato and Tjian 2007), Goat anti-TAF1 (Santa Cruz),
Rabbit anti-TAF3 (Deato and Tjian 2007), Mouse anti-TAF4 (BD Bioscience), Mouse
anti-TAF5 (Eurogentec), Rabbit anti-TAF6 (Abcam), Rabbit anti-TAF7 (Abnova), Goat
anti-TAF9 (Santa Cruz), Mouse anti-TAF10 (Chemicon), Rabbit anti-TAF12

Chapter 2: TBP, TRFs, and TAFs in Liver Development and Regeneration
(Proteintech Group), Rabbit anti-GAPDH (Cell Signaling), Goat anti-Albumin (Bethyl),
Mouse anti-AFP (R&D Systems), Mouse anti-AQP1 (BD Bioscience), Rabbit anti-
CYP7A1 (Santa Cruz).
Histone ChIP
Indivitual E13.5 livers were dissected from the embryos of timed pregnant CD-1
females and adult livers were dissected from 8-10 week old CD-1 females. Crosslinked
chromatin was prepared essentially as described (Soutoglou and Talianidis 2002; Kyrmizi
et al. 2006). Embryonic and adult liver tissue was minced in PBS and homogenized by
10-15 strokes of an A pestle in the presence of 1% formaldehyde. Crosslinking was
continued for 10 minutes and quenched by addition of glycine to 125mM. Cells were
collected at 1,000g for 10 minutes and washed twice for 20 minutes in hypotonic lysis
buffer (25mM Hepes pH7.9, 10mM KCl, 0.250M Sucrose, 2mM EDTA, 1.5mM MgCl2,
0.1% NP-40, 1mM DTT, 0.5mM Spermidine, Protease Inhibitor Cocktail (Roche)).
Nuclei were collected at 1,000g and resuspended in an equal volume of nuclear lysis
buffer (50mM Hepes pH7.9, 140mM NaCl, 1mM EDTA, 1% Triton X-100, 0.1% sodium
deoxycholate, 0.1% SDS, Protease Inhibitor Cocktail (Roche)). Chromatin was sheared
to 300-600 bp fragments by sonication and cleared by centrifugation at 15,000g. Equal
amounts of cleared chromatin were precipitated by overnight incubation with rabbit IgG,
rabbit anti-H3K4Me3, or rabbit anti-H3K9Me3 (Active Motif). Immunocomplexes were
incubated for two hours with pre-blocked protein A and protein G magnetic beads
(Invitrogen). Samples were washed twice with nuclear lysis buffer, twice with nuclear
lysis buffer plus 500mM NaCl, twice with Li was buffer (20mM Tris-HCl pH7.9, 1mM

Chapter 2: TBP, TRFs, and TAFs in Liver Development and Regeneration
EDTA, 250mM LiCl, 0.5% NP-40, 0.5% sodium deoxycholate), and once with TE.
Chromatin was eluted by addition of 50mM Tris-HCl pH 7.9, 25mM EDTA, 1% SDS,
0.2mg/ml RNase A, 0.1mg/ml Proteinase K. After incubation at 37˚C for 2 hours NaCl
was added to 500mM and samples were incubated overnight at 65˚C to reverse the
crosslinking. DNA was purified using QIAquick spin columns (Qiagen) and used in
quantitative PCR with primers surrounding the relevant proximal promoter.
Liver Cell Purification
Adult, day 15 neonatal, and partial hepatectomy hepatocytes were purified by
two-step in situ perfusion with Liberase 3 (Roche) as described (Wang et al. 2003b).
Partial hepatectomy of 8-12 week old male CD-1 mice was performed as described
(Mitchell and Willenbring 2008). Indivitual E13.5 livers were dissected from 3-5 timed
pregnant females in PBS and dissociated by incubation for 10-15 minutes at 37˚C in the
presence of 0.9mg/ml Liberase 3 (Roche) and 0.07% DNAse I (Sigma). Digestion was
quenched by the addition of FBS to 3% and EDTA to 2mM. Cells were collected at 40g
and resuspended in 1x Pharmalyse (BD Bioscience) for hypotonic lysis of erythrocytes.
Cells were resuspended in PBS/10%FBS, passed through a 70μm filter and diluted to
1x108 cells/ml. Cells were incubated for 15 minutes at 4˚C with CD45 microbeads
(Miltenyi Biotec), and in the absence of hypotonic lysis TER119 micobeads (Miltenyi
Biotec), after which labeled cells were removed by passage over an LS column attached
to a MidiMacs (Miltenyi Biotec) separator according the manufacturer’s instructions.
HPPL Cell Culture and Differentiation
Hepatic Progenitors Proliferating on Laminin (HPPL) were cultured and

Chapter 2: TBP, TRFs, and TAFs in Liver Development and Regeneration
differentiated essentially as described (Tanimizu et al. 2004). Proliferating cultures were
maintained in DMEM/F12 (Invitrogen), 10% heat inactivated FBS, 1x
insulin/transferrin/selenium (ITS) (Invitrogen), 10mM nicotinamide (Sigma), 0.1μm
dexamethasone (Novagen), 5mM L-glutamine (Invitrogen), 20ng/ml HGF (Peprotech),
and 20ng/ml EGF (Peprotech) on laminin-coated plates (BD Bioscience). Confluent
cultures were differentiated by incubation for five days in the same media minus HGF
and EGF and containing 20ng/ml OSM (Peprotech). For complete differentiation OSM
media was removed and cells were overlaid with fresh media containing 0.350 mg/ml
growth factor reduced Matrigel (BD Bioscience) for an additional five days as reported
(Kamiya et al. 2002).

Chapter 2: TBP, TRFs, and TAFs in Liver Development and Regeneration
Figure 1: TBP, TRF3, and TAF Expression in Mouse Tissues
C2C12 myoblasts and myotubes, CD-1 embryos, and whole mouse tissues were
homogenized in RIPA buffer and equal amounts of total protein were analyzed by
western blot with TBP, TRF3, TAF1, and TAF4 specific antibodies. GAPDH is used as a
protein loading control.
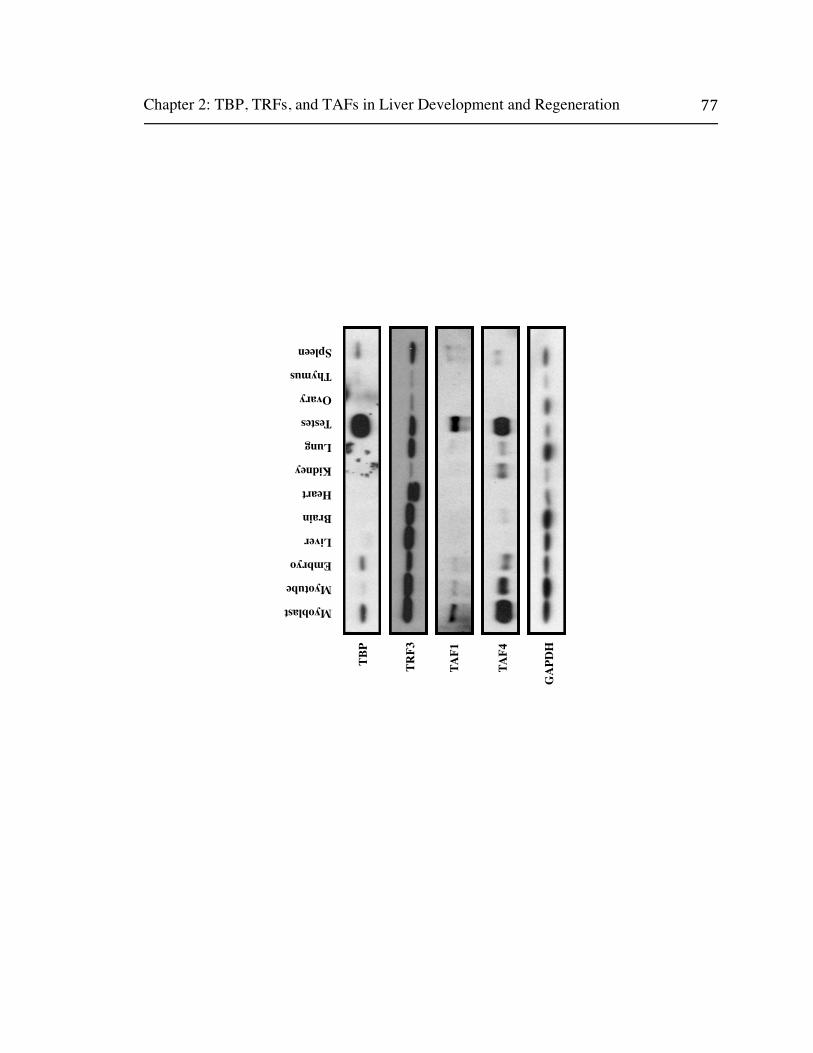
Chapter 2: TBP, TRFs, and TAFs in Liver Development and Regeneration
TBP
TRF3
TAF1
TAF4
GAPDH
Myoblast
Myotube
Embryo
Liver
Brain
Heart
Kidney
Lung
Testes
Ovary
Thymus
Spleen

Chapter 2: TBP, TRFs, and TAFs in Liver Development and Regeneration
Figure 2: TBP, TRF3, and TAF Expression in Mouse Tissues
Total RNA from diverse mouse tissues was reverse transcribed and used for
relative qPCR with transcript-specific Taqman probes. CT values were normalized to
18s ribosomal RNA (A) or GAPDH mRNA (B). Values for each tissue are expressed as
a percentage of the average value for all tissues combined.

Chapter 2: TBP, TRFs, and TAFs in Liver Development and Regeneration
Normalized to 18s
0
50
100
150
200
250
TBP TRF2 TRF3 TAF1 TAF3 TAF4 MED17
Embryo
Liver
Brain
Heart
Kidney
Lung
Testes
Ovary
Thymus
SpleenPer
cent
of A
vera
ge f
or A
ll T
issu
es
866%
A
Normalized to GAPDH
0
50
100
150
200
250
300
350
400
TBP TRF2 TRF3 TAF1 TAF3 TAF4 MED17
Embryo
Liver
Brain
Heart
Kidney
Lung
Testes
Ovary
Thymus
SpleenPer
cent
of A
vera
ge f
or A
ll T
issu
es
494%
896%
806%
B
452%

Chapter 2: TBP, TRFs, and TAFs in Liver Development and Regeneration
Figure 3: TBP, TRF, TAF, and Med Expression in Purified Liver Cells
Total RNA was purified from mouse E13.5 hepatoblasts and adult hepatocytes,
reverse transcribed, and used for qPCR. CT values were normalized to GAPDH and
expressed as a percentage of hepatoblast values. (A) TBP, TRFs and TAFs. (B)
Mediator subunits.

Chapter 2: TBP, TRFs, and TAFs in Liver Development and Regeneration
A
B
0
100
200
300
400
500
600
700
TB
PT
RF
2T
RF
3T
AF
1T
AF
2T
AF
3T
AF
4T
AF
4bT
AF
5T
AF
5LT
AF
6T
AF
6LT
AF
7T
AF
7LT
AF
8T
AF
9T
AF
9LT
AF
10T
AF
11T
AF
12T
AF
13T
AF
15G
CN
5P
CA
FB
TA
F1
Hepatoblast
Hepatocyte
0
20
40
60
80
100
120
Hepatoblast
Hepatocyte
Per
cent
of
Hep
atob
last
Per
cent
of
Hep
atob
last

Chapter 2: TBP, TRFs, and TAFs in Liver Development and Regeneration
Figure 4: TBP, TRF, TAF, and Med Expression in Neonatal and Regenerative
Hepatocytes.
Total RNA was purified from mouse E13.5 hepatoblasts, neonatal day 15
hepatocytes, regenerative hepatocytes following 2/3 partial hepatectomy, and adult
hepatocytes; reverse transcribed; and used for qPCR. CT values were normalized to
GAPDH and expressed as a percentage of hepatoblast values.

Chapter 2: TBP, TRFs, and TAFs in Liver Development and Regeneration
050100
150
200
250
300
350
Hep
atob
last
Neo
nata
l Hep
atoc
yte
Adu
lt H
epat
ocyt
e
Cyc
ling
Hep
atoc
yte
(2/3
par
tial
Hep
)
Percent of Hepatoblast

Chapter 2: TBP, TRFs, and TAFs in Liver Development and Regeneration
Figure 5: TBP, TRF and TAF Expression in Purified Liver Cells
Equal amounts of total protein from C2C12 cells, fetal and adult liver, and
purified liver cells were analyzed by western blotting with TBP, TRF2, and TAF-specific
antibodies.

Chapter 2: TBP, TRFs, and TAFs in Liver Development and Regeneration
TRF2
TBP
TAF1
TAF3
TAF4
TAF5
TAF7
TAF9
TAF10
TAF12
C2C
12
Fet
al L
iver
(E
13.5
)
Adu
lt L
iver
Hep
atob
last
Neo
nate
Hep
atoc
yte
Adu
lt H
epat
ocyt
e
Cyc
ling
Hep
atoc
yte

Chapter 2: TBP, TRFs, and TAFs in Liver Development and Regeneration
Figure 6: Med Expression in Purified Liver Cells
Equal amounts of total protein from C2C12 cells, fetal and adult liver, and
purified liver cells were analyzed by western blotting with antibodies against key
Mediator subunits.

Chapter 2: TBP, TRFs, and TAFs in Liver Development and Regeneration
MED26
MED18
MED23
MED16
MED17
MED12
MED14
MED7
MED1
MED6
CDK8
C2C
12
Fet
al L
iver
(E
13.5
)
Adu
lt L
iver
Hep
atob
last
Neo
nate
Hep
atoc
yte
Adu
lt H
epat
ocyt
e
Cyc
ling
Hep
atoc
yte

Chapter 2: TBP, TRFs, and TAFs in Liver Development and Regeneration
Figure 7: Transcriptional Silencing of TBP and TAF Promoters During
Hepatogenesis.
Individually dissected fetal and adult liver cells were crosslinked and used for
chromatin immunoprecipitation with histone mark-specific antibodies. Precipitated
chromatin was used in qPCR with primers surrounding the relevant proximal promoters
and values expressed as fold enrichment over an IgG control.

Chapter 2: TBP, TRFs, and TAFs in Liver Development and Regeneration
Fold Enrichment Fold Enrichment
E13
.5A
dult
051015202530354045
IgG
H3K
4Me3
H
3K9M
e3
TA
F3
Pro
mot
er
00.
511.
522.
533.
544.
55
IgG
H3K
4Me3
H
3K9M
e3
CY
P7A
1 P
rom
oter
05101520253035
IgG
H3K
4Me3
H
3K9M
e3
TA
F5
Pro
mot
er
0510152025
IgG
H3K
4Me3
H
3K9M
e3
TB
P P
rom
oter

Chapter 2: TBP, TRFs, and TAFs in Liver Development and Regeneration
Figure 8: Induction of Hepatic Gene Expression in a Cell Culture Model of
Development.
Hepatic Progenitors Proliferating on Laminin (HPPL) cells were differentiated by
incubating for five days in media containing 20ng/ml Oncostatin M and followed for five
days in media containing 0.350mg/ml Matrigel. Total RNA was reverse transcribed and
used in qPCR to analyze bipotential and hepatocyte marker gene expression (A). Values
are expressed as a percentage of undifferentiated control cells on a log scale. Whole cell
lysates were used in western blots with antibodies specific for -fetoprotein (AFP),
albumin (ALB), aquporin (AQP1), and CYP7A1 (B).

Chapter 2: TBP, TRFs, and TAFs in Liver Development and Regeneration
Day 0 5 10
AFP
ALB
AQP1
CYP7A1
A
B
1
10
100
1000
10000
100000
Day 0
Day 5
Day 10
Per
cent
Day
0 (
Log
Sca
le)

Chapter 2: TBP, TRFs, and TAFs in Liver Development and Regeneration
Figure 9: TBP, TRF, and TAF Expression in Differentiated HPPL Cells.
HPPLs were differentiated for five or ten days and total RNA was used for
quantitative PCR of TBP, TRF3, and TAF expression levels (A). CT values were
normalized to GAPDH and expressed as a percentage of an undifferentiated control.
Whole cell lysates were made and equal amounts of total protein analyzed by western
blot with antibodies specific to TBP, TRF3, TAF3, and TAF4 (A). GAPDH is used as a
protein loading control.

Chapter 2: TBP, TRFs, and TAFs in Liver Development and Regeneration
TBP
TAF4
TAF3
TRF3
GAPDH
Day 0 5 10
1
10
100
1000
10000
Day 0
Day 5
Day 10
A
B
Per
cent
Day
0

Chapter 2: TBP, TRFs, and TAFs in Liver Development and Regeneration
References
Alpini, G., Phillips, J.O., Vroman, B., and LaRusso, N.F. 1994. Recent advances in the
isolation of liver cells. Hepatology 20(2): 494-514.
Bernstein, B.E., Meissner, A., and Lander, E.S. 2007. The mammalian epigenome. Cell
128(4): 669-681.
Brunkhorst, A., Karlen, M., Shi, J., Mikolajczyk, M., Nelson, M.A., Metsis, M., and
Hermanson, O. 2005. A specific role for the TFIID subunit TAF4 and RanBPM in
neural progenitor differentiation. Mol Cell Neurosci 29(2): 250-258.
Cereghini, S. 1996. Liver-enriched transcription factors and hepatocyte differentiation.
FASEB J 10(2): 267-282.
Costa, R.H., Kalinichenko, V.V., Holterman, A.X., and Wang, X. 2003. Transcription
factors in liver development, differentiation, and regeneration. Hepatology 38(6):
1331-1347.
Daoust, R. and Cantero, A. 1959. The numerical proportions of cell types in rat liver
during carcinogenesis by 4-dimethylaminoazobenzene (DAB). Cancer Res 19:
757-762.
Deato, M.D., Marr, M.T., Sottero, T., Inouye, C., Hu, P., and Tjian, R. 2008. MyoD
targets TAF3/TRF3 to activate myogenin transcription. Mol Cell 32(1): 96-105.
Deato, M.D. and Tjian, R. 2007. Switching of the core transcription machinery during
myogenesis. Genes Dev 21(17): 2137-2149.
Freiman, R.N., Albright, S.R., Zheng, S., Sha, W.C., Hammer, R.E., and Tjian, R. 2001.
Requirement of tissue-selective TBP-associated factor TAFII105 in ovarian
development. Science 293(5537): 2084-2087.
Guglielmi, B., van Berkum, N.L., Klapholz, B., Bijma, T., Boube, M., Boschiero, C.,
Bourbon, H.M., Holstege, F.C., and Werner, M. 2004. A high resolution protein
interaction map of the yeast Mediator complex. Nucleic Acids Res 32(18): 5379-
5391.

Chapter 2: TBP, TRFs, and TAFs in Liver Development and Regeneration
Indra, A.K., Mohan, W.S., 2nd, Frontini, M., Scheer, E., Messaddeq, N., Metzger, D.,
and Tora, L. 2005. TAF10 is required for the establishment of skin barrier
function in foetal, but not in adult mouse epidermis. Dev Biol 285(1): 28-37.
Kamiya, A., Kinoshita, T., Ito, Y., Matsui, T., Morikawa, Y., Senba, E., Nakashima, K.,
Taga, T., Yoshida, K., Kishimoto, T. et al. 1999. Fetal liver development requires
a paracrine action of oncostatin M through the gp130 signal transducer. EMBO J
18(8): 2127-2136.
Kamiya, A., Kojima, N., Kinoshita, T., Sakai, Y., and Miyaijma, A. 2002. Maturation of
fetal hepatocytes in vitro by extracellular matrices and oncostatin M: induction of
tryptophan oxygenase. Hepatology 35(6): 1351-1359.
Kinoshita, T., Sekiguchi, T., Xu, M.J., Ito, Y., Kamiya, A., Tsuji, K., Nakahata, T., and
Miyajima, A. 1999. Hepatic differentiation induced by oncostatin M attenuates
fetal liver hematopoiesis. Proc Natl Acad Sci U S A 96(13): 7265-7270.
Kyrmizi, I., Hatzis, P., Katrakili, N., Tronche, F., Gonzalez, F.J., and Talianidis, I. 2006.
Plasticity and expanding complexity of the hepatic transcription factor network
during liver development. Genes Dev 20(16): 2293-2305.
Li, B., Carey, M., and Workman, J.L. 2007. The role of chromatin during transcription.
Cell 128(4): 707-719.
Martianov, I., Viville, S., and Davidson, I. 2002. RNA polymerase II transcription in
murine cells lacking the TATA binding protein. Science 298(5595): 1036-1039.
Mengus, G., Fadloun, A., Kobi, D., Thibault, C., Perletti, L., Michel, I., and Davidson, I.
2005. TAF4 inactivation in embryonic fibroblasts activates TGF beta signalling
and autocrine growth. EMBO J 24(15): 2753-2767.
Mitchell, C. and Willenbring, H. 2008. A reproducible and well-tolerated method for 2/3
partial hepatectomy in mice. Nat Protoc 3(7): 1167-1170.
Mohan, W.S., Jr., Scheer, E., Wendling, O., Metzger, D., and Tora, L. 2003. TAF10
(TAF(II)30) is necessary for TFIID stability and early embryogenesis in mice.
Mol Cell Biol 23(12): 4307-4318.

Chapter 2: TBP, TRFs, and TAFs in Liver Development and Regeneration
Muller, F., Lakatos, L., Dantonel, J., Strahle, U., and Tora, L. 2001. TBP is not
universally required for zygotic RNA polymerase II transcription in zebrafish.
Curr Biol 11(4): 282-287.
Myers, L.C. and Kornberg, R.D. 2000. Mediator of transcriptional regulation. Annu Rev
Biochem 69: 729-749.
Nitou, M., Sugiyama, Y., Ishikawa, K., and Shiojiri, N. 2002. Purification of fetal mouse
hepatoblasts by magnetic beads coated with monoclonal anti-e-cadherin
antibodies and their in vitro culture. Exp Cell Res 279(2): 330-343.
Perletti, L., Dantonel, J.C., and Davidson, I. 1999. The TATA-binding protein and its
associated factors are differentially expressed in adult mouse tissues. J Biol Chem
274(22): 15301-15304.
Perletti, L., Kopf, E., Carre, L., and Davidson, I. 2001. Coordinate regulation of
RARgamma2, TBP, and TAFII135 by targeted proteolysis during retinoic acid-
induced differentiation of F9 embryonal carcinoma cells. BMC Mol Biol 2: 4.
Persengiev, S.P., Robert, S., and Kilpatrick, D.L. 1996. Transcription of the TATA
binding protein gene is highly up-regulated during spermatogenesis. Mol
Endocrinol 10(6): 742-747.
Rogler, L.E. 1997. Selective bipotential differentiation of mouse embryonic hepatoblasts
in vitro. Am J Pathol 150(2): 591-602.
Schrem, H., Klempnauer, J., and Borlak, J. 2002. Liver-enriched transcription factors in
liver function and development. Part I: the hepatocyte nuclear factor network and
liver-specific gene expression. Pharmacol Rev 54(1): 129-158.
Soutoglou, E. and Talianidis, I. 2002. Coordination of PIC assembly and chromatin
remodeling during differentiation-induced gene activation. Science 295(5561):
1901-1904.
Strick-Marchand, H. and Weiss, M.C. 2002. Inducible differentiation and morphogenesis
of bipotential liver cell lines from wild-type mouse embryos. Hepatology 36(4 Pt
1): 794-804.

Chapter 2: TBP, TRFs, and TAFs in Liver Development and Regeneration
Suzuki, A., Zheng, Y., Kondo, R., Kusakabe, M., Takada, Y., Fukao, K., Nakauchi, H.,
and Taniguchi, H. 2000. Flow-cytometric separation and enrichment of hepatic
progenitor cells in the developing mouse liver. Hepatology 32(6): 1230-1239.
Taatjes, D.J., Marr, M.T., and Tjian, R. 2004. Regulatory diversity among metazoan co-
activator complexes. Nat Rev Mol Cell Biol 5(5): 403-410.
Tanimizu, N., Miyajima, A., and Mostov, K.E. 2007. Liver progenitor cells develop
cholangiocyte-type epithelial polarity in three-dimensional culture. Mol Biol Cell
18(4): 1472-1479.
Tanimizu, N., Nishikawa, M., Saito, H., Tsujimura, T., and Miyajima, A. 2003. Isolation
of hepatoblasts based on the expression of Dlk/Pref-1. J Cell Sci 116(Pt 9): 1775-
1786.
Tanimizu, N., Saito, H., Mostov, K., and Miyajima, A. 2004. Long-term culture of
hepatic progenitors derived from mouse Dlk+ hepatoblasts. J Cell Sci 117(Pt 26):
6425-6434.
Tatarakis, A., Margaritis, T., Martinez-Jimenez, C.P., Kouskouti, A., Mohan, W.S., 2nd,
Haroniti, A., Kafetzopoulos, D., Tora, L., and Talianidis, I. 2008. Dominant and
redundant functions of TFIID involved in the regulation of hepatic genes. Mol
Cell 31(4): 531-543.
Thomas, M.C. and Chiang, C.M. 2006. The general transcription machinery and general
cofactors. Crit Rev Biochem Mol Biol 41(3): 105-178.
Voss, A.K., Thomas, T., Petrou, P., Anastassiadis, K., Scholer, H., and Gruss, P. 2000.
Taube nuss is a novel gene essential for the survival of pluripotent cells of early
mouse embryos. Development 127(24): 5449-5461.
Wandzioch, E. and Zaret, K.S. 2009. Dynamic signaling network for the specification of
embryonic pancreas and liver progenitors. Science 324(5935): 1707-1710.
Wang, X., Foster, M., Al-Dhalimy, M., Lagasse, E., Finegold, M., and Grompe, M.
2003a. The origin and liver repopulating capacity of murine oval cells. Proc Natl
Acad Sci U S A 100 Suppl 1: 11881-11888.

Chapter 2: TBP, TRFs, and TAFs in Liver Development and Regeneration
Wang, X., Willenbring, H., Akkari, Y., Torimaru, Y., Foster, M., Al-Dhalimy, M.,
Lagasse, E., Finegold, M., Olson, S., and Grompe, M. 2003b. Cell fusion is the
principal source of bone-marrow-derived hepatocytes. Nature 422(6934): 897-
901.
Wauthier, E., Schmelzer, E., Turner, W., Zhang, L., LeCluyse, E., Ruiz, J., Turner, R.,
Furth, M.E., Kubota, H., Lozoya, O. et al. 2008. Hepatic stem cells and
hepatoblasts: identification, isolation, and ex vivo maintenance. Methods Cell Biol
86: 137-225.
Weiss, M.C. and Strick-Marchand, H. 2003. Isolation and characterization of mouse
hepatic stem cells in vitro. Semin Liver Dis 23(4): 313-324.
Wright, K.J., Marr, M.T., 2nd, and Tjian, R. 2006. TAF4 nucleates a core subcomplex of
TFIID and mediates activated transcription from a TATA-less promoter. Proc
Natl Acad Sci U S A 103(33): 12347-12352.
Zaret, K.S. 2002. Regulatory phases of early liver development: paradigms of
organogenesis. Nat Rev Genet 3(7): 499-512.
Zaret, K.S. and Grompe, M. 2008. Generation and regeneration of cells of the liver and
pancreas. Science 322(5907): 1490-1494.

Chapter 3: TRF3, TAF7l, and TAF13 in Hepatic Gene Regulation
Chapter 3: TRF3, TAF7l, and TAF13 in Hepatic Gene Regulation
Summary
The near infinite diversity of cell types and biological functions intrinsic to the
mammalian body plan is primarily determined at the level of transcriptional regulation.
In the mammalian liver, a diverse set of tissue-enriched and extensively studied
sequence-specific activators are known to regulate embryonic and adult gene expression
programs. While the spatial and temporal patterns of liver-enriched transcription factor
expression have been determined, and their requirements in hepatic gene expression
demonstrated, the exact mechanisms by which multiple factors collaborate to tune global
patterns of expression are not well understood. In contrast, the molecular mechanisms of
gene activation by canonical core promoter recognition and coactivator complexes such
as TFIID and Mediator are well established in cell culture and in vitro systems, but have
not yet extended to a tissue-specific context. Now that tissue-specific TAF paralogs,
TBP-related factors, and alternative core promoter recognition complexes have been
identified, it is critical to determine how these assemblies alter gene expression patterns
to specify tissue and cell-type-specific functions in collaboration with known activators.
To date, liver-specific core promoter recognition and coactivator complex subunits have
not been specifically characterized. If canonical TFIID is indeed absent from the adult
liver, than the isolation of such liver-enriched subunits and an understanding of their
coordination with existing liver-enriched transcription factors would greatly elucidate our
picture of liver development, disease, and regeneration.

Chapter 3: TRF3, TAF7l, and TAF13 in Hepatic Gene Regulation
Introduction
While TBP and the canonical TAFs which comprise TFIID were historically
assumed to be invariant between tissues, cell types and developmental time points, tissue-
specific TAF paralogs have since their discovery been known to exhibit highly restricted
expression patterns and transcriptional functions. Though a variety of TAF paralogs
were previously identified by bioinformatic means, the first concrete evidence of a tissue-
specific TFIID subunit came with the discovery of a substoichiometric TAF4 paralog in
cultured B cells (Dikstein et al. 1996). Subsequently, TAF4b was found to be highly
enriched in mouse testes and ovary and to be an intrinsic component of an altered ovary-
specific TFIID (Freiman et al. 2001). TAF4b knockout females, while viable, are
infertile as a result of deficient folliculogenesis, and knockout males show age-dependent
infertility resulting from a defect in spermatogenic maintenance (Falender et al. 2005).
Consistently, gene expression analysis revealed a requirement for TAF4b in regulating a
subset of ovarian granulosa cell-specific genes and genes required for spermatogonial
stem cell maintenance (Geles et al. 2006). Furthermore, TAF4b containing TFIID does
not necessarily require unique tissue-specific activators, but rather induces the expression
of and cooperates with ubiquitous general activators such as c-Jun to regulate cell-type-
specific gene expression (Geles et al. 2006). More detailed developmental analysis
confirmed that TAF4b is the primary transcriptional integrator of extracellular signals
that specify the developmental and proliferative program of granulosa cells (Voronina et
al. 2007). While not explicitly required in B cells, TAF4b cooperates with known
regulators of B cell-specific transcription such as OCA-B and NF B, and may serve a

Chapter 3: TRF3, TAF7l, and TAF13 in Hepatic Gene Regulation
redundant function to TAF4 in this context (Yamit-Hezi and Dikstein 1998; Wolstein et
al. 2000; Freiman et al. 2002). Promoter-specific differences in both basal and activated
transcription by canonical and TAF4b containing TFIID were demonstrated in vitro, and
these differences shown to correlate with TAF4b target genes identified in vivo (Liu et al.
2008). Intriguingly, a TAF4b homologue is enriched in Xenopus ovaries but absent from
lower metazoans, suggesting that unlike canonical TAFs, tissue-specific paralogs may
have evolved to direct species-specific transcriptional programs (Xiao et al. 2006)
The importance of tissue-specific TAFs was further validated by the identification
of five testis-restricted (tTAF) subunits in Drosophila, no hitter (paralog of TAF4),
cannonball (paralog of TAF5), meiosis I arrest (paralog of TAF6), spermatocyte arrest
(paralog of TAF8), and ryan express (paralogue of TAF12) (Lin et al. 1996). Disrupting
each of these proteins alters expression of testis-specific genes, and together they form a
stable complex, which is required for meiotic cell cycle progression and normal
spermatid differentiation (Hiller et al. 2001; Hiller et al. 2004). Intriguingly, the tTAF
complex mediates spermatid gene activation at least in part by displacing the repressive
polycomb complex and promoting its sequestration to the nucleolus (Chen et al. 2005);
hence, a derepressive tTAF strategy may represent a novel cell-type-specific mechanism
of TAF dependent coactivation.
While tTAFs are restricted to Drosophila, a testis-linked paralogue of TAF7
appears unique to vertebrates. High level expression of mouse TAF7l in late
spermatocytes and haploid spermatids is coincident with a reduction in TAF7 expression,
and as spermatogenesis proceeds TAF7l, is relocalized to the nucleus where it associates

Chapter 3: TRF3, TAF7l, and TAF13 in Hepatic Gene Regulation
with TBP and other TAFs to form an alternative TFIID- like complex (Pointud et al.
2003). Viable TAF7l knockout mice display altered patterns of spermatocyte gene
expression and defects in spermiogenesis which lead to reduced fertility, consistent with
a testes function (Cheng et al. 2007). Additional TAF paralogs, including TAF9b, while
shown to associate with TFIID and promote alternative gene expression programs, are
just beginning to be characterized, and the full repertoire of tissue-specific TAFs remains
to be discovered (Frontini et al. 2005).
While noncanonical TAF paralogs were originally identified in a subset of tissues,
TBP-related factors or TRFs were originally assumed to be widely expressed but are
now known to exhibit tissue-specific expression patterns and regulatory functions
(Hochheimer and Tjian 2003). Hence TRF1, the first identified TBP-related factor to be
discovered, is restricted to Drosophila, where it is enriched in the nervous system and
gonads and thus seems to have evolved to suite a specific purpose in these insect tissues
(Hansen et al. 1997). Polytene chromosome staining and genome-wide promoter
occupancy experiments suggest that TRF1 regulates a highly restricted set of genes
relative to TBP, and that these loci are primarily associated with RNA PolIII transcription
(Isogai et al. 2007b). However, TRF1 has been isolated in association with BRF and a
larger complex containing novel proteins and been shown to direct both RNA PolII and
PolII transcription in vitro and in vivo, much like TBP (Holmes and Tjian 2000; Takada
et al. 2000). Hence, TRF1 represents an evolutionarily restricted alternative core
promoter recognition complex which directs promoter-selective transcription by multiple
polymerases in a subset of tissues.

Chapter 3: TRF3, TAF7l, and TAF13 in Hepatic Gene Regulation
TBP-related factor 2 (TRF2, also called TLF, TLP, or TRP), though more widely
distributed evolutionarily from C. elegans to humans, also exhibits tissue-specific
expression patterns and possibly species-specific transcriptional specialization (Dantonel
et al. 1999; Rabenstein et al. 1999). While TRF2 binds TFIIA and TFIIB and displays
high conservation of the TBP DNA binding domain, it does not bind TATA box
containing DNA and fails to complement TFIID in basal or activated in vitro
transcription assays (Moore et al. 1999; Teichmann et al. 1999). Biochemically, TRF2
associates with the DRE binding factor (DREF) and components of the NURF chromatin
remodeling complex, and has been shown to selectively regulate the transcription of
DRE-containing promoters of cell cycle and proliferation genes in coordination with
these proteins (Hochheimer et al. 2002). Intriguingly, several genes have been shown to
contain tandem transcriptional start sites whose promoters are specifically and
differentially regulated by TRF2 and TFIID. Disruption experiments in C. elegans,
Drosophila, and Xenopus demonstrate a requirement for TRF2 in early embryonic
development, but also confirm that TRF2 regulates a subset of developmental genes
responsible for these phenotypes, while being dispensable for the expression of others
(Dantonel et al. 2000; Kaltenbach et al. 2000; Veenstra et al. 2000; Kopytova et al. 2006).
Conversely, mouse TRF2 is highly enriched in the testis, and while knockout animals
display spermiogenesis defects and altered patterns of testes-specific gene expression,
they show no discernable developmental defects, consistent with an evolutionarily
divergent function in mammals (Martianov et al. 2001; Zhang et al. 2001). Recent
genome-wide localization of Drosophila TRF2 showed that it occupies over a thousand

Chapter 3: TRF3, TAF7l, and TAF13 in Hepatic Gene Regulation
promoters, most of which are not bound by TBP and are enriched for DREs but lack a
canonical TATA box (Isogai et al. 2007a). Intriguingly, the Drosophila histone H1
promoter is specifically regulated by TRF2 while the reminder of the histone gene cluster
is TFIID dependent, suggesting that differences in core promoter recognition complex
activity may alter histone ratios. Hence, TRF2 exhibits evolutionarily diverse roles, and
clearly extends the diversity of core promoter recognition complexes in its regulation of
both tissue-specific and promoter-selective gene expression patterns. Additionally, the
existence of separate TRF2 and TFIID-dependent promoters within related gene clusters,
or in some cases single genes, may allow a limited set of promoters and core promoter
recognition complexes to produce dramatically varied expression patterns in response to
signaling events or between tissues. This alternative regulation likely results from
TRF2’s association with novel transcription factors through its divergent N and C-
terminal protein interaction domains and its recruitment to novel core promoter elements
not occupied by TBP.
The most recently identified and perhaps most intriguing member of the TBP-
related factor family is the vertebrate-specific TRF3 (also called TBP2). Initially
identified based on homology, mammalian TRF3 shows near identity to TBP in its C-
terminus including, the TATA box, TFIIA, and TFIIB binding domains, and considerable
divergence from all known TBPs in its N-terminus (Persengiev et al. 2003). Preliminary
characterization showed TRF3 to be widely expressed in adult mammalian tissues,
though at varying levels relative to TBP, and to be primarily contained in a larger 150-
200kD complex. Many studies of TRF3 function have focused on oogenesis and early

Chapter 3: TRF3, TAF7l, and TAF13 in Hepatic Gene Regulation
embryonic development. Hence, murine oocyte TRF3 expression has been shown to
increase rapidly during oocyte growth from primordial follicle formation until
preovulation at the same time that TBP expression is effectively eliminated (Yang et al.
2006; Gazdag et al. 2007). However, these observational reports reach conflicting
conclusions on the role of TRF3 in murine embryogenesis, as one shows a steady
increase in both TRF3 and TBP expression following fertilization, while the other
concludes that TRF3 remains silent as TBP expression is induced post fertilization; this
conflict in immunofluorescence data will likely be resolved by more extensive studies
and additional approaches.
The role of TRF3 in early development has been most extensively investigated in
Xenopus and zebrafish. Hence, Xenopus and zebrafish TRF3 bind TFIIA, TFIIB, and
TATA containing DNA, and Xenopus TRF3 support TATA-dependent transcription in
egg extracts (Bartfai et al. 2004; Jallow et al. 2004). While highly expressed in the
Xenopus oocyte, TRF3 is expressed at lower levels in the developing embryo and is
recruited to a subset of genes not occupied by TBP in a time-dependent manner.
Importantly, TRF3 depletion causes defects in Xenopus gastrulation and blastopore
closure and alters expression of many oocyte and embryo-specific genes, a subset of
which also require TBP. Intriguingly, Xenopus TRF3 overexpression can partially rescue
TBP knockdown phenotypes and gene expression patterns, consistent with a partial
redundancy. Further genome wide analysis found that Xenopus TBP primarily regulates
the synthesis of maternally deposited transcripts while a majority of embryonically
transcribed genes require TRF2 or TRF3 (Jacobi et al. 2007). Additionally, TBP

Chapter 3: TRF3, TAF7l, and TAF13 in Hepatic Gene Regulation
dependent Xenopus genes are widely conserved from yeast to humans while TRF3-
specific genes are generally restricted to vertebrates. Xenopus knockdown and ChIP
experiments demonstrate that many promoters while exclusively dependent on either
TRF3 or TFIID are occupied by both, suggesting that differences in complex-specific
transcription may occur at the level of activation rather than recruitment. Expression of
zebrafish TRF3 increases steadily from fertilization to gastrulation, after which it declines
rapidly but remains detectable in all adult tissues and enriched in the ovary. As with
Xenopus, knockdown inhibits normal gastrulation and results in mesodermal patterning
defects. Intriguingly, both species show an enrichment of TRF3 in the ventral embryo
consistent with a role in dorsoventral patterning. Additional knockdown studies in later
stage zebrafish embryos demonstrate a specific requirement for TRF3 in the initiation of
hematopoiesis, and partially define a lineage-specific transcriptional cascade in which
TRF3 is the master regulator of this particular differentiating program (Verdorfer et al.
2007). Thus, multiple studies in lower vertebrates reveal a requirement for TRF3 in early
development and elucidate both redundant and overlapping functions with TBP in
embryonic transcription. However, it is intriguing to note that the striking embryonic
phenotypes of TRF2 depletion in Xenopus and zebrafish are not conserved in the
knockout mice and that the important embryonic functions of TRF3 in lower vertebrates
likewise may not be preserved in mammals.
A study investigating the differentiation of mouse C2C12 cells found that while
TBP, along with several TAFs, are degraded upon differentiation, at least some TRF3 is
retained in myotubes, some of which is found in a complex with TAF3 (Deato and Tjian

Chapter 3: TRF3, TAF7l, and TAF13 in Hepatic Gene Regulation
2007). Depletion of TRF3 and TAF3 by RNAi suggested that this putative core promoter
recognition complex may be required for myogenesis. Consistent with a requisite role in
differentiation, ChIP analysis found TRF3/TAF3 occupying a key muscle-specific gene
promoter. Further biochemical analysis of the complex suggested that TRF3/TAF3 is
sufficient for modest activation of the myogenin promoter in vitro by the myogenic
transcription factors MyoD and E47 (Deato and Tjian 2008). Protein-protein interaction
studies, along with additional in vitro transcription assays, revealed that MyoD directly
targets TAF3, and that this interaction is necessary for in vitro activity. It will be
interesting to determine how widely the TRF3/TAF3 complex may be used in other
tissues.
The Mediator coactivator complex was also recently found to display
considerable subunit diversity, which ultimately may be shown to correlate with tissue
and cell-type-specific transcriptional programs. Hence, biochemical fractionation of
HeLa nuclear extracts yielded an altered CRSP/Med2 complex lacking two key canonical
Mediator subunits previously shown to be critical for nuclear hormone receptor (NHR)
dependent transcriptional activation (Taatjes and Tjian 2004). This complex, while
unable to support NHR dependent transcription, supports transcription by activators
which target other Mediator subunits and therefore may be responsible for limiting
coactivation to a subset of extracellular signals or promoters. Similarly, the ARC-L/Med
complex which contains most canonical Mediator subunits in addition to a dissociable
CDK8/cyclin C/Med230/Med240 submodule may fine tune Mediator activity in response
to environmental or developmental cues (Taatjes et al. 2002). Importantly, reversible

Chapter 3: TRF3, TAF7l, and TAF13 in Hepatic Gene Regulation
association of the ARC-L/Med submodule transitions C/EBPb dependent Mediator
activity from a stimulatory to a repressive state (Mo et al. 2004). While ARC-L/Med has
generally been associated with transcriptional repression it appears to stimulate a limited
subset of promoters, and thus the ratio of ARC-L/Med to Mediator may lead to the
activation of some genes and subsequent repression of others (Akoulitchev et al. 2000;
Zhou et al. 2002). Furthermore, the finding that the ARC-L/Med submodule enhances
Mediator’s association with a ubiquitin ligase and may enhance its proteasome-dependent
degradation suggests that submodule abundance may regulate coactivation globally by
influencing Mediator turnover (Brower et al. 2002) Thus, combinatorial control of
subunit composition could allow for cell-type and promoter-specific transcription
dependent on distinct classes of activators. However, it remains to be seen whether these
complexes are utilized in a cell-type-specific way.
Additional precedent for the idea of tissue and developmental stage-specific
changes in transcription factor complexes that were previously held to be invariant comes
from the recent observation that Swi/Snf chromatin remodeling complexes also vary
greatly between tissues. Hence, the finding that specific subunits of the mammalian
PBAF complex are enriched in cardiac progenitors and required for normal heart
maturation provided the earliest support for the idea of cell-type-specific chromatin
remodelers (Wang et al. 2004; Yan et al. 2005). Conversely, knockout studies of unique
subunits from the related BAF-A and BAF-B complexes demonstrated overlapping
deficiencies in ES cell pluripotency and lineage-specific differentiation, while disruption
of PBAF showed no specific ES cell phenotype, consistent with distinct developmental

Chapter 3: TRF3, TAF7l, and TAF13 in Hepatic Gene Regulation
requirements for each complex (Gao et al. 2008; Huang et al. 2008; Yan et al. 2008).
The first evidence of a requisite developmental stage-specific switch in subunit
composition came with the discovery that homologous BAF45 and BAF53 subunits must
be exchanged during the transition from neuronal progenitors to postmitotic neurons
(Lessard et al. 2007; Wu et al. 2007). Moreover, this exchange is now known to depend
on microRNA-mediated repression of the progenitor-specific subunits, adding an even
greater level of regulatory complexity to the neuronal transition (Yoo et al. 2009). Most
recently, a novel esBAF complex was isolated from embryonic stem cells which has a
unique subunit composition, is required for pluripotency and self-renewal, and is
downregulated upon differentiation (Ho et al. 2009b). Further genome-wide localization
experiments demonstrate the existence of a cooperative transcriptional network between
esBAF and known ES cell-specific transcription factors which defines self-renewal and
pluripotency (Ho et al. 2009a). The extensive evidence that TAF1 can bind and modify
histones, combined with the more recent finding that TAF3 shows a specific affinity for
active chromatin marks, further suggests that diverse cell or developmental stage-specific
chromatin remodelers and core promoter recognition complexes may cooperate with
specific chromatin states to direct highly specialized transcriptional programs (Mizzen et
al. 1996; Jacobson et al. 2000; Vermeulen et al. 2007).

Chapter 3: TRF3, TAF7l, and TAF13 in Hepatic Gene Regulation
Results
Hepatic Expression of TRF3, TAF7l, and TAF13
Because our previous qPCR analysis suggested that TRF3, TAF7l, and TAF13
expression may not be as dramatically downregulated as TBP and other TAFs in adult
hepatocytes or may in fact be induced upon differentiation, we chose to further explore
the expression and function of these specific subunits in hepatic transcription (Chapter 2,
Figure 3). Hence, we performed western blots of purified hepatoblasts and hepatocytes
with multiple antibodies specific for these three subunits (Figure 1). At least two
separate anti-TRF3 antibodies recognize strong expression of TRF3 expression in both
developing hepatoblasts and committed hepatocytes (Figure 1A). Within the quantitative
limits of western blot analysis, TRF3 expression in liver cells does not appear to differ
greatly from that in the whole ovary, a tissue in which TRF3 was previously shown to be
highly enriched (Gazdag et al. 2009). While qPCR data suggest that TRF3 expression
may be slightly greater in hepatocytes than hepatoblasts, quantitative western blotting or
other techniques need to be employed to determine if this is true at the protein level.
Similarly, western blots with two different TAF7l antibodies suggest that this
paralog is expressed at similar levels in hepatocytes and developing hepatoblasts and at
comparable levels to the testes, the tissue from which TAF7l was first isolated and shown
to have a specific function (Figure 1B). Again, more quantitative proteomic methods will
need to be employed to determine if the significant upregulation of TAF7l transcript
observed in adult hepatocytes leads to an increase in protein levels. Initial western blot
comparison of TAF13 expression in developing hepatoblasts and mature hepatocytes also

Chapter 3: TRF3, TAF7l, and TAF13 in Hepatic Gene Regulation
suggests that this subunit may be retained or induced in the adult liver, in contrast to the
majority of canonical TFIID components (Figure 1C). While further quantitation of
TAF13 polypeptide is needed, initial data suggest that this subunit warrants further
investigation.
RNAi Mediated Depletion of TRF3, TAF7l, and TAF13 Attenuates Hepatic Gene
Induction in HPPLs
While spatial and temporal patterns of transcription factor expression often
correlate with tissue-specific function, this is not always the case, and some factors may
be inactive or dispensable even where they are highly abundant. To explore the
possibility that TRF3, TAF7l, and TAF13 may play a role in adult hepatic gene
transcription, we depleted these proteins by lentiviral RNAi in the HPPL cell line.
pLKO.1 vectors which are part of the Broad Institute RNAi consortium and express short
hairpin RNAs targeting these three transcripts under the U6 promoter, along with a
puromycin resistance gene, were used to generate replication-incompetent lentiviral
particles by transient transfection of HEK293T cells (Figure 2A). Proliferating HPPLs
were infected for 24 hours with viral supernatants and then selected for four days in fresh
growth medium containing puromycin to eliminate uninfected cells. Cells were then
harvested to determine the extent of gene knockdown by qPCR or differentiated in the
presence of Oncostatin M. By this approach we consistently achieved knockdown of
TRF3, TAF7l, and TAF13 with sixty to eighty percent efficiency (Figure 2B).
Importantly, infection with a control lentivirus targeting GFP had no discernible affect on

Chapter 3: TRF3, TAF7l, and TAF13 in Hepatic Gene Regulation
the levels of these three transcripts, and reduction of any one transcript did not influence
the expression of another, suggesting that off target effects and lentivirally induced
cytotoxicity are minimal in this system.
Because the most significant changes in both TFIID downregulation and hepatic
gene induction in the HPPL cell model were observed after five days of differentiation,
we reasoned that this represented the most critical period for transcription factor activity
in this particular system (Chapter 1, Figure 8 and 9). Following four days of selection,
confluent lentivirally transduced HPPLs were differentiated for five days in the presence
of Oncostatin M and RNA harvested for qPCR analysis. This data shows that even
moderate depletion of TRF3, TAF7l, and TAF13 has a significant impact on the
induction of multiple adult hepatic genes in the HPPL model, as judged by transcript
abundance (Figure 3). Qualitatively, differentiated TRF3, TAF7l, and TAF13 shRNA
expressing HPPLs contained fewer large cuboidal, binucleate, and vacuolated cells than
the GFP control, suggesting that these cultures failed to fully differentiate. Though it
cannot be concluded from this data that TRF3, TAF7l, or TAF13 specifically regulate the
hepatic genes in question, this approach suggests that these factors have some influence
on the HPPLs’ competence to differentiate towards a mature hepatic cell and that in their
absence, hepatic maturation is at least partially compromised.

Chapter 3: TRF3, TAF7l, and TAF13 in Hepatic Gene Regulation
TRF3, TAF7l, and TAF13 are Enriched at Hepatic Promoters in the Adult Liver
and Differentiated HPPLs.
The physical recruitment of sequence-specific activators to their target enhancers
and of the general machinery to a gene’s proximal promoters is a requisite step in
transcriptional activation (Thomas and Chiang 2006). Hence, the ability to localize
proteins and complexes at the regulatory elements of a given gene supports a role for
such factors in that gene’s transcriptional regulation. Chromatin immunoprecipitation is
a powerful technique for investigating the relative occupancy of defined DNA domains
by specific proteins and has been extensively employed to elucidate the functions of
single transcription factors and the complexities of large regulatory networks (Massie and
Mills 2008; Park 2009). Specifically, this approach has been invaluable in unraveling the
role of liver-enriched activators and general transcription factors in liver development
and maintenance (Kyrmizi et al. 2006; Tatarakis et al. 2008).
To investigate the role of TRF3, TAF7l, and TAF13 in hepatic transcription we
isolated formaldehyde crosslinked chromatin from individually dissected E13.5 and adult
mouse livers, and subjected it to immunoprecipitation with antibodies against these three
factors. Precipitated material was analyzed by qPCR with primers surrounding the
promoters of adult hepatic genes (Figure 4). Intriguingly, TRF3, TAF7l, and TAF13
show overlapping and distinct patterns of promoter occupancy on multiple hepatocyte-
specific genes. Most notably all three factors are significantly and specifically enriched
on the promoters of 1-antitrypsin (AAT) and tryptophan-2,3-dioxygenase (TDO2), two
genes which are completely silent in the embryo and are highly induced late postnatally.

Chapter 3: TRF3, TAF7l, and TAF13 in Hepatic Gene Regulation
Importantly, these promoters show no enrichment of the factors at day E13.5, a time
when they are known to be transcriptionally silenced. Intriguingly, a second class of
promoters, including apolipoprotein C3 (APOCIII) and possibly apolipoprotein B
(APOB) and albumin (ALB), are occupied by TRF3, TAF7l, and possibly TAF13 in both
the embryo and adult. In contrast to AAT and TDO2, these promoters are expressed in
the embryo and continue to be expressed at high levels postnatally. Recruitment of
TRF3, TAF7l, and TAF13 is less pronounced and varies between factors on the other
promoters examined; however the extent of factor recruitment is significant and specific
and its timing is consistent with the known developmental upregulation for these
promoters.
The primary drawback of chromatin immunoprecipitation is its dependence on the
specificity and efficiency of available antibodies for the transcription factor in question.
Because of this, overexpression of exogenous epitope-tagged proteins followed by
precipitation with well characterized antibodies against those tags has proven valuable to
corroborate data obtained with antibodies against the endogenous factor. To this end, we
generated recombinant TRF3, TAF7l, and TAF13 constructs with N-terminal FLAG tags
in the doxycycline-inducible pTRIPZ vector; these vectors were used to produce
lentiviral particles which were used to transduce proliferative HPPLs. Two days post
infection confluent cells were differentiated for five days by the addition of oncostatin M,
and pTRIPZ expression was induced by the addition of doxycycline to 2μg/ml on the last
two days of differentiation. Cells were crosslinked and purified chromatin was
immunoprecipitated with IgG or anti-FLAG polyclonal antibodies. Precipitated material

Chapter 3: TRF3, TAF7l, and TAF13 in Hepatic Gene Regulation
was analyzed by qPCR and results expressed as fold enrichment over the IgG control
(Figure 5).
While the promoter enrichment of FLAG-tagged TRF3, TAF7l, and TAF13 in
differentiated HPPLs does not exactly mirror that seen in adult liver, this data clearly
demonstrates that all three factors are specifically recruited to adult hepatic genes (Figure
5). The efficiency of FLAG-tagged protein immunoprecipitation may be impacted by the
presentation of the N-terminal FLAG epitope on these proteins in a native environment;
likewise the extent of promoter occupancy may be impacted by the ability of these
exogenous proteins to associate with other factors required for their recruitment or the
presence of these factors in not fully differentiated HPPL. In particular, the reduced
recruitment of TAF7l likely results from the relatively low expression and cytotoxicity
that we observe with this construct and that may inhibit differentiation or adequate
promoter binding. However, the data show that all three factors are enriched at multiple
hepatic promoters by an approach that is independent of antibody specificity.
Furthermore, the relative fold enrichment is similar to that seen in whole liver and
previously reported for TAFs, as well as general transcription factors on hepatic genes in
whole liver and liver cell line samples (Tatarakis et al. 2008).
TRF3 Associates with One or More High Molecular Weight Complexes in the Adult
Liver.
The finding that TRF3, TAF7l, and TAF13 are expressed in the adult liver, are
enriched at overlapping hepatic promoters, and may be required for hepatic gene

Chapter 3: TRF3, TAF7l, and TAF13 in Hepatic Gene Regulation
expression raised the possibility that they interact to form a coordinated multisubunit
regulatory complex. Indeed, one of the defining characteristics of known core promoter
recognition and coactivator complexes including TFIID and Mediator is their existence as
stable high molecular weight assemblies under native conditions (Albright and Tjian
2000; Myers and Kornberg 2000). Additionally, TRF3 has previously been shown to
associate with a 150kD complex in HeLa cells and to form a 150kD complex with TAF3
in myotubes (Persengiev et al. 2003; Deato and Tjian 2007).
To explore this possibility, we prepared nuclear extracts from adult mouse livers
and subjected them to size exclusion chromatography on a Superdex-200 column
previously calibrated with known sizing standards. Western blot analysis of those
fractions corresponding to molecular weights from 440 to 13.7kD reveal that TRF3
migrates at approximately 150kD as previously reported in the HeLa and myotube
systems. However, TAF7l and TAF13 do not appear, under these conditions, to
significantly associate with slower migrating TRF3, as they both migrate at or near their
calculated monomeric molecular weights of 42 and 18kD respectively. Our finding that
TAF3 is downregulated transcriptionally during hepatic development and seemingly
absent from adult hepatocytes at the protein level led us to believe that the TRF3/TAF3
complex previously identified in myotubes would be absent in the hepatic system
(Chapter 2, Figure 3 and 5). However, the observation that TRF3 remains associated
with a 150kD complex in liver nuclear extracts suggests that either trace amounts of
TAF3 remain, or that TRF3 is associated with other as yet to be identified subunits.
While the slower migration of TRF3 clearly does not depend on its association with the

Chapter 3: TRF3, TAF7l, and TAF13 in Hepatic Gene Regulation
majority of TAF7l or TAF13 under these conditions, this does not rule out the possibility
that these three proteins interact more transiently in vivo or that minor amounts of TAF7l
and TAF13, which avoid detection in this setting, are associated with higher molecular
weight assemblies. It is important to note that neither TAF7l nor TAF13 is observed in
the void fraction that would contain canonical TFIID. Though many general
transcription factor assemblies survive biochemical fractionation under the conditions
employed, activator-coactivator interactions are typically weaker, more transient, and
refractory to efficient biochemical copurification. Clearly, further investigation is needed
to determine the full range of protein-protein interactions by TAF7l and TAF13.
Core Promoter Recognition Complex Changes in Human ES Cell Derived
Hepatocytes.
Canonical TFIID structure and function has been a subject of almost equally
intensive study in both the mouse and human systems, and its role in transcription has
been shown to be highly conserved between these and lower vertebrates. In contrast,
investigation of TBP-related factors and TAF paralogs, including TRF3 and TAF7l, has
been largely restricted to the murine, Xenopus, and zebrafish models. Furthermore,
significant differences in the mechanisms and processes of liver development and
regeneration have been identified between mice and humans (Wauthier et al. 2008).
Hence, novel mechanisms of hepatic regulation uncovered by murine experimentation
must be recapitulated and confirmed in the human system for their therapeutic potential
to be realized. The directed in vitro differentiation of human embryonic stem cells to

Chapter 3: TRF3, TAF7l, and TAF13 in Hepatic Gene Regulation
functioning hepatocytes with the potential for therapeutic transplantation has been
reported by several groups and is a subject of intensive ongoing investigation (Duan et al.
2007; Hay et al. 2007; Hay et al. 2008a). Thus, we wished to determine if the changes
we observe in core promoter recognition complex expression during murine liver
development are conserved in an ES cell-based model of human hepatic commitment.
Human H9 ES cells were differentiated using a three step, sixteen day protocol
which mimics in vivo signaling events and has a reported efficiency of 90% (Hay et al.
2008a). In the first stage, proliferating ESCs are committed to the definitive endoderm
by incubation for five days with Activin A and Wnt3A, in the second stage they are
differentiated to hepatoblast like cells in defined growth factor free media, and in the
final stage matured to hepatocyte-like cells by the addition of hepatocyte growth factor
and oncostatin M (Figure 7A). Analysis of known marker gene expression in these four
cell populations by qPCR mirrors which is observed in vivo and is consistent with
published reports of directed hepatocyte differentiation, suggesting that the procedure
does faithfully model the hepatogenic process (Hay et al. 2008a; Hay et al. 2008b).
Intriguingly, qPCR analysis of TBP, three critical TAF subunits, and MED17 in these
four cell populations shows that TFIID and Mediator levels are held constant or induced
during the first two stages and reduced dramatically during hepatogenic maturation
(Figure 7B). Thus, as is observed in the purified mouse liver cell samples and HPPL
protocol, this model suggests that human hepatogenesis requires significant changes in
TFIID and Mediator expression or composition.

Chapter 3: TRF3, TAF7l, and TAF13 in Hepatic Gene Regulation
Discussion
Intensive and ongoing study of liver-enriched sequence-specific activators has
contributed greatly to our understanding of the transcriptional changes that govern
hepatic development and regeneration (Schrem et al. 2002; Costa et al. 2003). However,
the molecular mechanisms by which unique combinations of activators alter global
patterns of gene expression in response to developmental inputs or hepatic injury remains
to be elucidated. Core promoter recognition and coactivator complexes are a critical link
between activators, chromatin modifiers, core promoter DNA, and the general PolII
machinery. Despite this, the composition and function of these complexes in liver
development, regeneration, and homeostasis has not been thoroughly explored.
Our observation that canonical TFIID, including TBP and multiple TAFs, is
significantly downregulated during hepatic development raised the possibility that
alternative core promoter recognition proteins exist in the adult liver and at least partially
replace the critical functions of TFIID (Chapter 2). This hypothesis was informed by
several recent studies that demonstrate a role for TBP-related factors and TAF paralogs is
defined differentiation programs or cell types. Specifically, muscle cell development and
embryonic hematopoiesis were independently shown to require a novel TRF3/TAF3 core
promoter recognition complex which at least partially replaces TFIID in the regulation of
developmental stage-specific genes (Deato and Tjian 2007; Hart et al. 2009).
Additionally, TRF3 was found to be a critical regulator of oocyte-specific genes during
primary folliculogenesis, while TAF7l was shown to be essential for spermiogenesis,
potentially as part of a novel TBP containing complex (Cheng et al. 2007; Gazdag et al.

Chapter 3: TRF3, TAF7l, and TAF13 in Hepatic Gene Regulation
2009). Our qPCR analysis of known TRF and TAF transcripts in hepatoblasts and
hepatocytes suggests that, unlike TBP and the majority of TAFs, TRF3, TAF7l, and
TAF13, expression is maintained or induced in committed liver cells and that the role of
these factors in hepatic transcription therefore warrants further investigation.
While more accurate quantitation of TRF3, TAF7l, and TAF13 polypeptide is
required, initial data confirms expression of these three proteins in adult hepatocytes and
suggests that they are therefore unique from the majority of TFIID subunits which are
significantly downregulated. A more thorough understanding of liver organogenesis
should include expression studies of TFIID and these three factors in the other major cell
types, including Cholangiocytes and Kupffer cells, to determine if retention of these
factors is specific to the hepatocyte lineage or a hallmark of multiple liver cells. Initial
RNAi experiments demonstrate that depletion of TRF3, TAF7l, and TAF13 alters
patterns of hepatic gene induction, suggesting that their presence is in fact functionally
relevant. More detailed HPPL studies will be needed to determine if RNAi of these
factors alters cell proliferation or competence prior to hepatocyte commitment,
suggestive of a role in hepatoblast transcription, or if the observed phenotype reflects a
specific requirement for these factors solely during the process of hepatic gene induction.
While gene expression is an important indirect readout of hepatic development,
differentiated HPPLs have been shown to exhibit hepatocyte like metabolic functions
such that it will be interesting to determine if RNAi impacts more functional readouts,
including albumin secretion and cytochrome P450 metabolism (Kamiya et al. 2002).
Additionally rescue experiments are needed to confirm the specificity of shRNA

Chapter 3: TRF3, TAF7l, and TAF13 in Hepatic Gene Regulation
targeting. Mouse knockout and knockdown experiments have helped to confirm the in
vivo relevance of both liver-enriched transcription factors and TFIID components in other
tissues (Lee et al. 2005; Mengus et al. 2005; Tatarakis et al. 2008) Whole animal
knockdown studies are currently underway and should help to corroborate and expand the
findings of TRF3, TAF7l, and TAF13 RNAi experiments conducted in the HPPL system.
Morphological and functional phenotypes resulting from gene ablation
experiments often reflect secondary or tertiary effects and thus fail to prove that a given
promoter is the direct target of the transcription factor in question. The chromatin
immunoprecipitation data presented demonstrates that at least some adult hepatic genes
are specifically occupied by TRF3, TAF7l, and TAF13 and suggests that the observed
RNAi phenotype is not exclusively the product of secondary effects. In light of these
results, ChIP and Re-ChIP analysis should be extended to determine more accurately the
timing and potential co-occupancy of additional adult and embryonic promoters in
multiple liver cell types. A major goal of hepatology research has been to uncover the
overlapping and autoregulatory actions of all known liver-enriched transcription factors
throughout development, and to use this data in defining an all inclusive regulatory
network (Odom et al. 2004; Kyrmizi et al. 2006). If novel core promoter recognition
complexes prove to be global regulators of hepatic transcription, then chromatin
immunoprecipitation combined with recent deep sequencing techniques has the potential
to identify all promoters regulated by these factors and refine or expand considerably our
understanding of the hepatogenic transcription factor network (Park 2009). Reconstituted
in vitro transcription using defined factors was recently employed to confirm the

Chapter 3: TRF3, TAF7l, and TAF13 in Hepatic Gene Regulation
regulation of a key myogenic promoter by the TRF3/TAF3 complex in coordination with
a known myogenic activator, and was previously shown to be a valuable technique in the
mechanistic study of a tissue-specific noncanonical TAF4 paralogs (Deato et al. 2008;
Liu et al. 2008). Hence, if liver-specific core promoter recognition proteins and their
target promoters can be sufficiently defined, then in vitro assays with known liver-
enriched transactivators may prove valuable in further uncovering the mechanisms of
hepatocyte-specific transcriptional activation.
While our data confirm the association of TRF3 with a high molecular weight
complex in adult liver as previously reported for other cell types, initial experiments
suggest that it does not physically associate with either TAF7l or TAF13 under the
conditions employed (Hart et al. 2009). Clearly, further investigation is needed to
determine if TAF7l and TAF13 possess unique or overlapping binding partners in
committed hepatocytes and if such proteins are required for their promoter recruitment or
coactivator function. One outstanding question is whether either of these factors
associates with canonical TFIID in the fetal or adult liver and if this association is
strengthened or eliminated during hepatogenic differentiation. While specific TAFs have
been shown to bind known transcriptional activators and thus facilitate recruitment of
holo-TFIID to the core promoter, no specific interaction has yet been reported between
canonical TFIID subunits and liver-specific activators (Albright and Tjian 2000). It will
be interesting for future studies to determine if known liver-specific activators exhibit
preferential binding to TFIID or novel hepatocyte enriched core promoter recognition

Chapter 3: TRF3, TAF7l, and TAF13 in Hepatic Gene Regulation
proteins and if these interactions are consistent with previously identified target genes
and patterns of expression for the activator in question.
Ongoing efforts to develop therapeutically relevant transplantable hepatocytes
from human embryonic stem cells have benefitted greatly from basic advances in our
understanding of liver development (Hay et al. 2007; Hay et al. 2008a). Our initial
finding that hepatogenic in vitro differentiation of human ESC involves significant
downregulation of TFIID as first observed in the mouse system, suggests that core
promoter recognition complex changes are not unique to the murine system and may
prove useful in the refinement of ESC differentiation protocols. In light of these
observations and the recent generation of iPS cells from mature hepatocytes, it will be
interesting to determine the function of TFIID and potentially novel core promoter
recognition factors, not only in hepatocyte commitment but in the process of iPS
induction, pluripotency, and self-renewal (Aoi et al. 2008).
It is now clear that TFIID, Mediator, and potentially other general transcription
factor complexes, which were once thought to be a hallmark of all cell types and gene
expression programs, are dispensable in at least some developmental strategies and
tissue-specific cell types. The role of TBP-related factors, TAF paralogs, and
noncanonical core promoter recognition or coactivator complexes have become
increasingly important as new techniques have allowed us to study rarer and more
defined populations of cells. The recent finding that myogenesis and early hematopoiesis
employ a stripped down TRF3/TAF3 complex in place of canonical TFIID, and that germ
cell development relies on multiple TFIID independent factors including TRF2, TRF3

Chapter 3: TRF3, TAF7l, and TAF13 in Hepatic Gene Regulation
and TAF7l raises the possibility that additional differentiation programs will require these
and as yet unidentified proteins which supplant TFIID at critical developmental junctions.
Our current data suggest that the commitment of hepatogenic precursors to mature
hepatocytes likewise involves the downregulation of canonical TFIID subunits and their
possible replacement with TRF3, TAF7l, or TAF13 and potentially other unknown core
promoter recognition elements.
Materials and Methods
Antibodies
Fresh TRF3, TAF7l, and TAF13 antibodies were generated by multiple strategies.
Rabbits, Guinea pigs, and chickens were injected with an N-terminal TRF3 peptide
coupled to Keyhole Limpet Hemocyanin (Pierce) or a recombinant TRF3-Pseudomonas
exotoxin fusion protein expressed from pVCH6 and purified under denaturing conditions.
Rabbit and chicken anti-TAF7l antibodies were raised against a Pseudomonas exotoxin
fusion-TAF7l fusion protein purified under denaturing conditions. Mouse anti-TAF13
antibodies were raised against full length recombinant Drosophila TAF13 purified under
denaturing conditions. Rabbit and mouse polyclonal antibodies were purified on Protein
A sepharose (Amersham) and chicken antibodies were purified using the IgY Purification
Kit (Pierce). Mouse monoclonal anti-TBP/TRF3 antibody 58C9 was used for western
blotting at a dilution of 1:1,000 (Sigma, (Hoey et al. 1993).

Chapter 3: TRF3, TAF7l, and TAF13 in Hepatic Gene Regulation
Plasmids
shRNA constructs in the pLKO.1 plasmid are part of the Broad Institute RNAi
library and were purchased from Open Biosystems. The lentiviral packaging plasmids
pSPAX2 and pMD2.G were purchased from Addgene. The TRF3, TAF7l, and TAF13
open reading frames were PCR amplified to contain N-Terminal FLAG tags and C-
Terminal HA, V5, and MYC tags respectively. PCR products were cloned into the AgeI
and MluI sites of the Doxycycline inducible pTRIPZ vector (Open Biosystems). All
constructs were sequenced and purified using Endotoxin Free Maxi or Giga kits
(Qiagen).
Size-Exclusion Chromatography
Nuclear extracts were prepared from 8-10 week old CD-1 males as reported
(Conaway et al. 1996) and fractionated on a 24ml Superdex-200 column (GE Healthcare)
using an AKTAexplorer. Fractions were analyzed by western blot and calibrated using
the Gel Filtration Calibration Kit according the manufacturer’s instructions (GE
Healthcare).
Lentiviral RNAi
Recombinant lentivirus was generated by transient cotransfection of 293T cells
with pLKO.1 or pTRIPZ (Openbiosystems) and pSPAX2/pMD2.G (Addgene) using
Lipofectamine 2000 (Invitrogen). Viral supernatants were used to infect proliferating
HPPL cells in the presence of 8ug/ml hexadimethrine bromide and 24 hours post
infection cells were selected with 1.5ug/ml Puromycin (Sigma). RNA from

Chapter 3: TRF3, TAF7l, and TAF13 in Hepatic Gene Regulation
lentivirus infected cells was isolated using RNeasy kits (Qiagen) and used for RT-
qPCR.
Chromatin Immunoprecipitation
Fetal and adult liver chromatin was prepared essentially as reported (Kyrmizi et
al. 2006). Individually dissected adult and E13.5 livers were minced with razor blades in
PBS and homogenized in 1% formaldyde by 10-15 strokes of an A pestle. Crosslinking
was continued for 10 minutes at room temperature and quenched by addition of glycine
to 125mM. Cells were washed twice in PBS and twice for 20 minutes in cell lysis buffer
(25mM Hepes pH7.9, 10mM KCl, 0.250M Sucrose, 2mM EDTA, 1.5mM MgCl2, 0.1%
NP-40, 1mM DTT, 0.5mM Spermidine, Protease Inhibitor Cocktail (Roche) followed by
passage through a 100μm filter. Nuclei were collected by centrifugation resuspended in
two pellet volumes of nuclear lysis buffer (50mM Hepes pH7.9, 140mM NaCl, 1mM
EDTA, 1% Triton X-100, 0.1% sodium deoxycholate, 0.1% SDS, Protease Inhibitor
Cocktail (Roche)), and chromatin was sonicated for 10 minutes (30 second on cycles) on
high with a Bioruptor (Diagenode). Chromatin was clearly by centrifugation at 12,000g
for 10 minutes and equal A260 units were precipitated overnight with protein A purified
rabbit anti-TRF3, TAF7l, and TAF13 antibodies or Rabbit IgG (Jackson
Immunoresearch). Protein A and Protein G beads were blocked overnight with 5mg/ml
BSA and 5mg/ml herring sperm DNA (Invitrogen), and added to the chromatin
immuncomplexes for 1 hour. Beads were washed three times with IP buffer, once with
IP buffer plus 500mM NaCl, twice with LiCl wash buffer (20mM Tris-HCl pH7.9, 1mM
EDTA, 250mM LiCl, 0.5% NP-40, 0.5% sodium deoxycholate), and once with TE.

Chapter 3: TRF3, TAF7l, and TAF13 in Hepatic Gene Regulation
Complexes were eluted by incubation for 30 minutes at 37˚C in elution buffer plus
10ug/ml RNase A (Qiagen) followed by two hours at 65˚C in the presence of 10ug/ml
Proteinase K (Invitrogen). Crosslinking was reversed overnight at 65 and DNA purified
using Qiagen spin columns before qPCR with promoter-specific primers. Chromatin
from pTRIPZ infected cell lines was prepared by addition of formaldehyde to the cell
culture media to a final concentration of 1% for 10 minutes at room temperature followed
the addition of glycine to 125mM. Chromatin was prepared as above and precipitated
with rabbit IgG or rabbit anti-FLAG polyclonal antibody (Sigma).
H9 hESC Differentiation
Human H9 hESC cells were maintained on irradiated mouse embryonic
fibroblasts (MEFs) following WICell protocols and differentiated as reported (Hay et al.
2007; Hay et al. 2008a). Cells were passaged to Matrigel (BD Bioscience) coated plates
in 10% MEF conditioned media and at 70% confluence induced to differentiate by the
addition of priming media containing RPMI1640 (Invitrogen), 1xB27 (Invitrogen),
50ng/ml Wnt3a (R&D Systems), and 100ng/ml Activin A (Peprotech). After five days
cells were split 1:2 onto fresh Matrigel and cultured in differentiation media containing
knockout-DMEM (Invitrogen), 20% Serum Replacement (Invitrogen), 1mM glutamine,
1% nonessential amino acids (Invitrogen), 0.1mM -mercaptoethanol, and 1% DMSO.
Finally, cells were matured in L15 medium (Invitrogen) supplemented with 8.3% FBS,
8.3% tryptose phosphate broth (Sigma), 10μM hydrocortisone 21-hemisuccinate (Sigma),
1μM insulin, 2mM glutamine, 10ng/ml hepatocyte growth factor (Peprotech), and

Chapter 3: TRF3, TAF7l, and TAF13 in Hepatic Gene Regulation
20ng/ml Oncostatin M. After seven days RNA was harvested for reverse transcription
and qPCR with validated gene-specific primers.

Chapter 3: TRF3, TAF7l, and TAF13 in Hepatic Gene Regulation
Figure 1: Expression of TRF3, TAF7l, and TAF13 in purified liver cells.
Recombinant TRF3, whole ovary lysate, adult liver lysate, hepatoblasts, and
hepatocytes were subjected to western blotting with mouse monoclonal antibody 58C9
and rabbit anti-TRF3 antibodies (A). Recombinant TAF7l, testes lysate, fetal and adult
liver lysate, hepatoblasts and hepatocytes were subjected to western blotting with two
different polyclonal anti-TAF7l antibodies (B). Hepatoblast and hepatocyte lysates were
subjected to western blotting with mouse anti-TAF13 antibody (C).

Chapter 3: TRF3, TAF7l, and TAF13 in Hepatic Gene Regulation
rTR
F3
Ova
ry
Adu
lt L
iver
Hep
tobl
ast
Hep
atoc
yte
Mouse Anti-TRF3(58C9)
Rabbit Anti-TRF3(22598)
A
rTA
F7l
Tes
tes
Fet
al L
iver
(E
13.5
)
Adu
lt L
iver
Hep
tobl
ast
Hep
atoc
yte
Rabbit Anti-TAF7l(HYRB1)
Rabbit Anti-TAF7l(23266)
B
Hep
tobl
ast
Hep
atoc
yte
Mouse Anti-TAF13(31)
C

Chapter 3: TRF3, TAF7l, and TAF13 in Hepatic Gene Regulation
Figure 2: shRNA Mediated Depletion of TRF3, TAF7l, and TAF13 in HPPLs.
The pLKO.1 vector, which contains lentiviral 5’ and 3’ leader and other
regulatory sequences, expresses short hairpin RNAs from the human U6 promoter and
puromycin N-acetyl-transferase from the PKG promoter (A). Proliferating HPPLs were
infected for 24 hours with pLKO.1 viral supernatants expressing the indicated shRNA,
selected for four days with 1.5μg/ml Puromycin and knockdown assessed by qPCR (B).

Chapter 3: TRF3, TAF7l, and TAF13 in Hepatic Gene Regulation
5’ LTR 3’ LTRRRE cPPT PGK-PuroU6 shRNA
A
B
0
20
40
60
80
100
120
TRF3 TAF7l TAF13
shGFP
shTRF3
shTAF7l
shTAF13
Per
cent
of
shG
FP
Con
trol

Chapter 3: TRF3, TAF7l, and TAF13 in Hepatic Gene Regulation
Figure 3: Expression of Adult Hepatic Genes in Differentiated and pLKO.1
Tranduced HPPLs.
Following four days of selection, five days post infection, HPPLs were
differentiated for five days in the presence of 20ng/ml Oncostatin M and the induction of
hepatic genes was analyzed by qPCR.

Chapter 3: TRF3, TAF7l, and TAF13 in Hepatic Gene Regulation
020406080100
120
Cyp
2F2
Cyp
7B1
Fas
nIG
F1
Mup
3Sl
co1a
1F
oxA
2H
NF
1aH
NF
4aG
6PC
TA
TT
DO
2
shG
FP
shT
RF
3
shT
AF
7l
shT
AF
13
Percent of shGFP Control

Chapter 3: TRF3, TAF7l, and TAF13 in Hepatic Gene Regulation
Figure 4: Chromatin Immunoprecipitation in Fetal and Adult Liver
E13.5 and adult livers were individually dissected, crosslinked, and chromatin
was immunoprecipitated with IgG or TRF3, TAF7l, and TAF13 antibodies. Crosslinking
was reversed and precipitated DNA was analyzed by qPCR. Signals were normalized to
input and expressed as fold enrichment over the IgG control.

Chapter 3: TRF3, TAF7l, and TAF13 in Hepatic Gene Regulation
0
1
2
3
4
5
6
7
8
E13.5-IgG
Adult-IgG
E13.5-TRF3
Adult-TRF3
AF
old
Enr
ichm
ent
02468
101214161820
E13.5-IgG
Adult-IgG
E13.5-TAF7l
Adult-TAF7l
B
Fol
d E
nric
hmen
t
0
10
20
30
40
50
60
E13.5-IgG
Adult-IgG
E13.5-TAF7l
Adult-TAF7l
C
Fol
d E
nric
hmen
t

Chapter 3: TRF3, TAF7l, and TAF13 in Hepatic Gene Regulation
Figure 5: Chromatin Immunoprecipitation in Differentiated HPPLs.
Proliferating HPPLs were infected for 24 hours with pTRIPZ based lentivirus
expressing FLAG tagged TRF3, TAF7l or TAF13 and differentiated for five days in the
presence of 20 ng/ml Oncostatin M. Cells were crosslinked and chromatin was subjected
to immunoprecipitation with IgG or anti-FLAG polyclonal antibody. Crosslinking was
reversed and precipitated DNA analyzed by qPCR. Signals were normalized to input and
expressed as fold enrichment of the IgG control.

Chapter 3: TRF3, TAF7l, and TAF13 in Hepatic Gene Regulation
0510152025
IgG
FL
AG
-TR
F3
FL
AG
-TA
F7l
FL
AG
-TA
F13
Fold Enrichment

Chapter 3: TRF3, TAF7l, and TAF13 in Hepatic Gene Regulation
Figure 6: Size Exclusion Chromatography of Adult Liver Nuclear Extract.
Adult mouse liver nuclear extract was fractionated on a Superdex-200 column and
fractions analyzed by western blot with TRF3, TAF7l, and TAF13 antibodies. Relative
molecular weights were calibrated with the standards in HMW and LMW Gel Filtration
Calibration Kit.

Chapter 3: TRF3, TAF7l, and TAF13 in Hepatic Gene Regulation
0 1 2 3 4 5 6 7 8 9 10 11 12
Inpu
t (N
E)
Voi
d44
0 kD
158
kD
43 k
D
13.7
kD
TRF3
TAF7l
TAF13

Chapter 3: TRF3, TAF7l, and TAF13 in Hepatic Gene Regulation
Figure 7: Hepatic Differentiation of Human H9 ES Cells.
Human H9 ES cells were differentiated to mature hepatocyte-like cells by a three
stage protocol that mimics in vivo signaling events (A). Analysis of known marker genes
for ES cells, definitive endoderm, hepatoblasts, and mature hepatocytes by qPCR shows
that this protocol faithfully mimics human hepatogenesis (B).

Chapter 3: TRF3, TAF7l, and TAF13 in Hepatic Gene Regulation
110100
1000
1000
0
1000
00
1000
000
1000
0000
Oct
4
Sox1
7C
XC
R4
Met
Krt
19A
qp1
Pro
x1A
fpA
lbG
6PC
HN
F3b
AA
TT
DO
2C
PS1
hESC
(H9)
Stag
e 1
Stag
e 2
Stag
e 3
Percent of hESC(H9) (Log Scale)
hESC
(H9)
Stag
e 1
Stag
e 2
Stag
e 3
A B

Chapter 3: TRF3, TAF7l, and TAF13 in Hepatic Gene Regulation
Figure 8: Expression of TBP, TRFs, and TAFs in H9 Differentiation
Expression of TBP, TAF1, TAF3, TAF5, and MED17 as judged by qPCR,
remains stable during definitive endoderm and hepatoblast commitment, but decreases
significantly during hepatocyte maturation in the H9 system.

Chapter 3: TRF3, TAF7l, and TAF13 in Hepatic Gene Regulation
0
50
100
150
200
250
TBP TAF1 TAF3 TAF5 MED17
hESC(H9)
Stage 1
Stage 2
Stage 3Per
cent
of
hESC
(H9)

Chapter 3: TRF3, TAF7l, and TAF13 in Hepatic Gene Regulation
References
Akoulitchev, S., Chuikov, S., and Reinberg, D. 2000. TFIIH is negatively regulated by cdk8-containing mediator complexes. Nature 407(6800): 102-106.
Albright, S.R. and Tjian, R. 2000. TAFs revisited: more data reveal new twists and confirm old ideas. Gene 242(1-2): 1-13.
Aoi, T., Yae, K., Nakagawa, M., Ichisaka, T., Okita, K., Takahashi, K., Chiba, T., and Yamanaka, S. 2008. Generation of pluripotent stem cells from adult mouse liver and stomach cells. Science 321(5889): 699-702.
Bartfai, R., Balduf, C., Hilton, T., Rathmann, Y., Hadzhiev, Y., Tora, L., Orban, L., and Muller, F. 2004. TBP2, a vertebrate-specific member of the TBP family, is required in embryonic development of zebrafish. Curr Biol 14(7): 593-598.
Brower, C.S., Sato, S., Tomomori-Sato, C., Kamura, T., Pause, A., Stearman, R., Klausner, R.D., Malik, S., Lane, W.S., Sorokina, I. et al. 2002. Mammalian mediator subunit mMED8 is an Elongin BC-interacting protein that can assemble with Cul2 and Rbx1 to reconstitute a ubiquitin ligase. Proc Natl Acad Sci U S A 99(16): 10353-10358.
Chen, X., Hiller, M., Sancak, Y., and Fuller, M.T. 2005. Tissue-specific TAFs counteract Polycomb to turn on terminal differentiation. Science 310(5749): 869-872.
Cheng, Y., Buffone, M.G., Kouadio, M., Goodheart, M., Page, D.C., Gerton, G.L., Davidson, I., and Wang, P.J. 2007. Abnormal sperm in mice lacking the Taf7l gene. Mol Cell Biol 27(7): 2582-2589.
Conaway, R.C., Reines, D., Garrett, K.P., Powell, W., and Conaway, J.W. 1996. Purification of RNA polymerase II general transcription factors from rat liver. Methods Enzymol 273: 194-207.
Costa, R.H., Kalinichenko, V.V., Holterman, A.X., and Wang, X. 2003. Transcription factors in liver development, differentiation, and regeneration. Hepatology 38(6): 1331-1347.

Chapter 3: TRF3, TAF7l, and TAF13 in Hepatic Gene Regulation
Dantonel, J.C., Quintin, S., Lakatos, L., Labouesse, M., and Tora, L. 2000. TBP-like factor is required for embryonic RNA polymerase II transcription in C. elegans. Mol Cell 6(3): 715-722.
Dantonel, J.C., Wurtz, J.M., Poch, O., Moras, D., and Tora, L. 1999. The TBP-like factor: an alternative transcription factor in metazoa? Trends Biochem Sci 24(9): 335-339.
Deato, M.D., Marr, M.T., Sottero, T., Inouye, C., Hu, P., and Tjian, R. 2008. MyoD targets TAF3/TRF3 to activate myogenin transcription. Mol Cell 32(1): 96-105.
Deato, M.D. and Tjian, R. 2007. Switching of the core transcription machinery during myogenesis. Genes Dev 21(17): 2137-2149.
-. 2008. An unexpected role of TAFs and TRFs in skeletal muscle differentiation: switching core promoter complexes. Cold Spring Harb Symp Quant Biol 73: 217-225.
Dikstein, R., Zhou, S., and Tjian, R. 1996. Human TAFII 105 is a cell type-specific TFIID subunit related to hTAFII130. Cell 87(1): 137-146.
Duan, Y., Catana, A., Meng, Y., Yamamoto, N., He, S., Gupta, S., Gambhir, S.S., and Zern, M.A. 2007. Differentiation and enrichment of hepatocyte-like cells from human embryonic stem cells in vitro and in vivo. Stem Cells 25(12): 3058-3068.
Falender, A.E., Freiman, R.N., Geles, K.G., Lo, K.C., Hwang, K., Lamb, D.J., Morris, P.L., Tjian, R., and Richards, J.S. 2005. Maintenance of spermatogenesis requires TAF4b, a gonad-specific subunit of TFIID. Genes Dev 19(7): 794-803.
Freiman, R.N., Albright, S.R., Chu, L.E., Zheng, S., Liang, H.E., Sha, W.C., and Tjian, R. 2002. Redundant role of tissue-selective TAF(II)105 in B lymphocytes. Mol Cell Biol 22(18): 6564-6572.
Freiman, R.N., Albright, S.R., Zheng, S., Sha, W.C., Hammer, R.E., and Tjian, R. 2001. Requirement of tissue-selective TBP-associated factor TAFII105 in ovarian development. Science 293(5537): 2084-2087.

Chapter 3: TRF3, TAF7l, and TAF13 in Hepatic Gene Regulation
Frontini, M., Soutoglou, E., Argentini, M., Bole-Feysot, C., Jost, B., Scheer, E., and Tora, L. 2005. TAF9b (formerly TAF9L) is a bona fide TAF that has unique and overlapping roles with TAF9. Mol Cell Biol 25(11): 4638-4649.
Gao, X., Tate, P., Hu, P., Tjian, R., Skarnes, W.C., and Wang, Z. 2008. ES cell pluripotency and germ-layer formation require the SWI/SNF chromatin remodeling component BAF250a. Proc Natl Acad Sci U S A 105(18): 6656-6661.
Gazdag, E., Rajkovic, A., Torres-Padilla, M.E., and Tora, L. 2007. Analysis of TATA-binding protein 2 (TBP2) and TBP expression suggests different roles for the two proteins in regulation of gene expression during oogenesis and early mouse development. Reproduction 134(1): 51-62.
Gazdag, E., Santenard, A., Ziegler-Birling, C., Altobelli, G., Poch, O., Tora, L., and Torres-Padilla, M.E. 2009. TBP2 is essential for germ cell development by regulating transcription and chromatin condensation in the oocyte. Genes Dev 23(18): 2210-2223.
Geles, K.G., Freiman, R.N., Liu, W.L., Zheng, S., Voronina, E., and Tjian, R. 2006. Cell-type-selective induction of c-jun by TAF4b directs ovarian-specific transcription networks. Proc Natl Acad Sci U S A 103(8): 2594-2599.
Hansen, S.K., Takada, S., Jacobson, R.H., Lis, J.T., and Tjian, R. 1997. Transcription properties of a cell type-specific TATA-binding protein, TRF. Cell 91(1): 71-83.
Hart, D.O., Santra, M.K., Raha, T., and Green, M.R. 2009. Selective interaction between Trf3 and Taf3 required for early development and hematopoiesis. Dev Dyn 238(10): 2540-2549.
Hay, D.C., Fletcher, J., Payne, C., Terrace, J.D., Gallagher, R.C., Snoeys, J., Black, J.R., Wojtacha, D., Samuel, K., Hannoun, Z. et al. 2008a. Highly efficient differentiation of hESCs to functional hepatic endoderm requires ActivinA and Wnt3a signaling. Proc Natl Acad Sci U S A 105(34): 12301-12306.

Chapter 3: TRF3, TAF7l, and TAF13 in Hepatic Gene Regulation
Hay, D.C., Zhao, D., Fletcher, J., Hewitt, Z.A., McLean, D., Urruticoechea-Uriguen, A., Black, J.R., Elcombe, C., Ross, J.A., Wolf, R. et al. 2008b. Efficient differentiation of hepatocytes from human embryonic stem cells exhibiting markers recapitulating liver development in vivo. Stem Cells 26(4): 894-902.
Hay, D.C., Zhao, D., Ross, A., Mandalam, R., Lebkowski, J., and Cui, W. 2007. Direct differentiation of human embryonic stem cells to hepatocyte-like cells exhibiting functional activities. Cloning Stem Cells 9(1): 51-62.
Hiller, M., Chen, X., Pringle, M.J., Suchorolski, M., Sancak, Y., Viswanathan, S., Bolival, B., Lin, T.Y., Marino, S., and Fuller, M.T. 2004. Testis-specific TAF homologs collaborate to control a tissue-specific transcription program. Development 131(21): 5297-5308.
Hiller, M.A., Lin, T.Y., Wood, C., and Fuller, M.T. 2001. Developmental regulation of transcription by a tissue-specific TAF homolog. Genes Dev 15(8): 1021-1030.
Ho, L., Jothi, R., Ronan, J.L., Cui, K., Zhao, K., and Crabtree, G.R. 2009a. An embryonic stem cell chromatin remodeling complex, esBAF, is an essential component of the core pluripotency transcriptional network. Proc Natl Acad Sci U S A 106(13): 5187-5191.
Ho, L., Ronan, J.L., Wu, J., Staahl, B.T., Chen, L., Kuo, A., Lessard, J., Nesvizhskii, A.I., Ranish, J., and Crabtree, G.R. 2009b. An embryonic stem cell chromatin remodeling complex, esBAF, is essential for embryonic stem cell self-renewal and pluripotency. Proc Natl Acad Sci U S A 106(13): 5181-5186.
Hochheimer, A. and Tjian, R. 2003. Diversified transcription initiation complexes expand promoter selectivity and tissue-specific gene expression. Genes Dev 17(11): 1309-1320.
Hochheimer, A., Zhou, S., Zheng, S., Holmes, M.C., and Tjian, R. 2002. TRF2 associates with DREF and directs promoter-selective gene expression in Drosophila. Nature 420(6914): 439-445.

Chapter 3: TRF3, TAF7l, and TAF13 in Hepatic Gene Regulation
Hoey, T., Weinzierl, R.O., Gill, G., Chen, J.L., Dynlacht, B.D., and Tjian, R. 1993. Molecular cloning and functional analysis of Drosophila TAF110 reveal properties expected of coactivators. Cell 72(2): 247-260.
Holmes, M.C. and Tjian, R. 2000. Promoter-selective properties of the TBP-related factor TRF1. Science 288(5467): 867-870.
Huang, X., Gao, X., Diaz-Trelles, R., Ruiz-Lozano, P., and Wang, Z. 2008. Coronary development is regulated by ATP-dependent SWI/SNF chromatin remodeling component BAF180. Dev Biol 319(2): 258-266.
Isogai, Y., Keles, S., Prestel, M., Hochheimer, A., and Tjian, R. 2007a. Transcription of histone gene cluster by differential core-promoter factors. Genes Dev 21(22): 2936-2949.
Isogai, Y., Takada, S., Tjian, R., and Keles, S. 2007b. Novel TRF1/BRF target genes revealed by genome-wide analysis of Drosophila Pol III transcription. EMBO J 26(1): 79-89.
Jacobi, U.G., Akkers, R.C., Pierson, E.S., Weeks, D.L., Dagle, J.M., and Veenstra, G.J. 2007. TBP paralogs accommodate metazoan- and vertebrate-specific developmental gene regulation. EMBO J 26(17): 3900-3909.
Jacobson, R.H., Ladurner, A.G., King, D.S., and Tjian, R. 2000. Structure and function of a human TAFII250 double bromodomain module. Science 288(5470): 1422-1425.
Jallow, Z., Jacobi, U.G., Weeks, D.L., Dawid, I.B., and Veenstra, G.J. 2004. Specialized and redundant roles of TBP and a vertebrate-specific TBP paralog in embryonic gene regulation in Xenopus. Proc Natl Acad Sci U S A 101(37): 13525-13530.
Kaltenbach, L., Horner, M.A., Rothman, J.H., and Mango, S.E. 2000. The TBP-like factor CeTLF is required to activate RNA polymerase II transcription during C. elegans embryogenesis. Mol Cell 6(3): 705-713.
Kamiya, A., Kojima, N., Kinoshita, T., Sakai, Y., and Miyaijma, A. 2002. Maturation of fetal hepatocytes in vitro by extracellular matrices and oncostatin M: induction of tryptophan oxygenase. Hepatology 35(6): 1351-1359.

Chapter 3: TRF3, TAF7l, and TAF13 in Hepatic Gene Regulation
Kopytova, D.V., Krasnov, A.N., Kopantceva, M.R., Nabirochkina, E.N., Nikolenko, J.V., Maksimenko, O., Kurshakova, M.M., Lebedeva, L.A., Yerokhin, M.M., Simonova, O.B. et al. 2006. Two isoforms of Drosophila TRF2 are involved in embryonic development, premeiotic chromatin condensation, and proper differentiation of germ cells of both sexes. Mol Cell Biol 26(20): 7492-7505.
Kyrmizi, I., Hatzis, P., Katrakili, N., Tronche, F., Gonzalez, F.J., and Talianidis, I. 2006. Plasticity and expanding complexity of the hepatic transcription factor network during liver development. Genes Dev 20(16): 2293-2305.
Lee, C.S., Friedman, J.R., Fulmer, J.T., and Kaestner, K.H. 2005. The initiation of liver development is dependent on Foxa transcription factors. Nature 435(7044): 944-947.
Lessard, J., Wu, J.I., Ranish, J.A., Wan, M., Winslow, M.M., Staahl, B.T., Wu, H., Aebersold, R., Graef, I.A., and Crabtree, G.R. 2007. An essential switch in subunit composition of a chromatin remodeling complex during neural development. Neuron 55(2): 201-215.
Lin, T.Y., Viswanathan, S., Wood, C., Wilson, P.G., Wolf, N., and Fuller, M.T. 1996. Coordinate developmental control of the meiotic cell cycle and spermatid differentiation in Drosophila males. Development 122(4): 1331-1341.
Liu, W.L., Coleman, R.A., Grob, P., King, D.S., Florens, L., Washburn, M.P., Geles, K.G., Yang, J.L., Ramey, V., Nogales, E. et al. 2008. Structural changes in TAF4b-TFIID correlate with promoter selectivity. Mol Cell 29(1): 81-91.
Martianov, I., Fimia, G.M., Dierich, A., Parvinen, M., Sassone-Corsi, P., and Davidson, I. 2001. Late arrest of spermiogenesis and germ cell apoptosis in mice lacking the TBP-like TLF/TRF2 gene. Mol Cell 7(3): 509-515.
Massie, C.E. and Mills, I.G. 2008. ChIPping away at gene regulation. EMBO Rep 9(4): 337-343.
Mengus, G., Fadloun, A., Kobi, D., Thibault, C., Perletti, L., Michel, I., and Davidson, I. 2005. TAF4 inactivation in embryonic fibroblasts activates TGF beta signalling and autocrine growth. EMBO J 24(15): 2753-2767.

Chapter 3: TRF3, TAF7l, and TAF13 in Hepatic Gene Regulation
Mizzen, C.A., Yang, X.J., Kokubo, T., Brownell, J.E., Bannister, A.J., Owen-Hughes, T., Workman, J., Wang, L., Berger, S.L., Kouzarides, T. et al. 1996. The TAF(II)250 subunit of TFIID has histone acetyltransferase activity. Cell 87(7): 1261-1270.
Mo, X., Kowenz-Leutz, E., Xu, H., and Leutz, A. 2004. Ras induces mediator complex exchange on C/EBP beta. Mol Cell 13(2): 241-250.
Moore, P.A., Ozer, J., Salunek, M., Jan, G., Zerby, D., Campbell, S., and Lieberman, P.M. 1999. A human TATA binding protein-related protein with altered DNA binding specificity inhibits transcription from multiple promoters and activators. Mol Cell Biol 19(11): 7610-7620.
Myers, L.C. and Kornberg, R.D. 2000. Mediator of transcriptional regulation. Annu Rev Biochem 69: 729-749.
Odom, D.T., Zizlsperger, N., Gordon, D.B., Bell, G.W., Rinaldi, N.J., Murray, H.L., Volkert, T.L., Schreiber, J., Rolfe, P.A., Gifford, D.K. et al. 2004. Control of pancreas and liver gene expression by HNF transcription factors. Science 303(5662): 1378-1381.
Park, P.J. 2009. ChIP-seq: advantages and challenges of a maturing technology. Nat Rev Genet 10(10): 669-680.
Persengiev, S.P., Zhu, X., Dixit, B.L., Maston, G.A., Kittler, E.L., and Green, M.R. 2003. TRF3, a TATA-box-binding protein-related factor, is vertebrate-specific and widely expressed. Proc Natl Acad Sci U S A 100(25): 14887-14891.
Pointud, J.C., Mengus, G., Brancorsini, S., Monaco, L., Parvinen, M., Sassone-Corsi, P., and Davidson, I. 2003. The intracellular localisation of TAF7L, a paralogue of transcription factor TFIID subunit TAF7, is developmentally regulated during male germ-cell differentiation. J Cell Sci 116(Pt 9): 1847-1858.
Rabenstein, M.D., Zhou, S., Lis, J.T., and Tjian, R. 1999. TATA box-binding protein (TBP)-related factor 2 (TRF2), a third member of the TBP family. Proc Natl Acad Sci U S A 96(9): 4791-4796.

Chapter 3: TRF3, TAF7l, and TAF13 in Hepatic Gene Regulation
Schrem, H., Klempnauer, J., and Borlak, J. 2002. Liver-enriched transcription factors in liver function and development. Part I: the hepatocyte nuclear factor network and liver-specific gene expression. Pharmacol Rev 54(1): 129-158.
Taatjes, D.J., Naar, A.M., Andel, F., 3rd, Nogales, E., and Tjian, R. 2002. Structure, function, and activator-induced conformations of the CRSP coactivator. Science 295(5557): 1058-1062.
Taatjes, D.J. and Tjian, R. 2004. Structure and function of CRSP/Med2; a promoter-selective transcriptional coactivator complex. Mol Cell 14(5): 675-683.
Takada, S., Lis, J.T., Zhou, S., and Tjian, R. 2000. A TRF1:BRF complex directs Drosophila RNA polymerase III transcription. Cell 101(5): 459-469.
Tatarakis, A., Margaritis, T., Martinez-Jimenez, C.P., Kouskouti, A., Mohan, W.S., 2nd, Haroniti, A., Kafetzopoulos, D., Tora, L., and Talianidis, I. 2008. Dominant and redundant functions of TFIID involved in the regulation of hepatic genes. Mol Cell 31(4): 531-543.
Teichmann, M., Wang, Z., Martinez, E., Tjernberg, A., Zhang, D., Vollmer, F., Chait, B.T., and Roeder, R.G. 1999. Human TATA-binding protein-related factor-2 (hTRF2) stably associates with hTFIIA in HeLa cells. Proc Natl Acad Sci U S A 96(24): 13720-13725.
Thomas, M.C. and Chiang, C.M. 2006. The general transcription machinery and general cofactors. Crit Rev Biochem Mol Biol 41(3): 105-178.
Veenstra, G.J., Weeks, D.L., and Wolffe, A.P. 2000. Distinct roles for TBP and TBP-like factor in early embryonic gene transcription in Xenopus. Science 290(5500): 2312-2315.
Verdorfer, I., Hollrigl, A., Strasser, U., Susani, M., Hartmann, A., Rogatsch, H., and Mikuz, G. 2007. Molecular-cytogenetic characterisation of sex cord-stromal tumours: CGH analysis in sertoli cell tumours of the testis. Virchows Arch 450(4): 425-431.

Chapter 3: TRF3, TAF7l, and TAF13 in Hepatic Gene Regulation
Vermeulen, M., Mulder, K.W., Denissov, S., Pijnappel, W.W., van Schaik, F.M., Varier, R.A., Baltissen, M.P., Stunnenberg, H.G., Mann, M., and Timmers, H.T. 2007. Selective anchoring of TFIID to nucleosomes by trimethylation of histone H3 lysine 4. Cell 131(1): 58-69.
Voronina, E., Lovasco, L.A., Gyuris, A., Baumgartner, R.A., Parlow, A.F., and Freiman, R.N. 2007. Ovarian granulosa cell survival and proliferation requires the gonad-selective TFIID subunit TAF4b. Dev Biol 303(2): 715-726.
Wang, Z., Zhai, W., Richardson, J.A., Olson, E.N., Meneses, J.J., Firpo, M.T., Kang, C., Skarnes, W.C., and Tjian, R. 2004. Polybromo protein BAF180 functions in mammalian cardiac chamber maturation. Genes Dev 18(24): 3106-3116.
Wauthier, E., Schmelzer, E., Turner, W., Zhang, L., LeCluyse, E., Ruiz, J., Turner, R., Furth, M.E., Kubota, H., Lozoya, O. et al. 2008. Hepatic stem cells and hepatoblasts: identification, isolation, and ex vivo maintenance. Methods Cell Biol 86: 137-225.
Wolstein, O., Silkov, A., Revach, M., and Dikstein, R. 2000. Specific interaction of TAFII105 with OCA-B is involved in activation of octamer-dependent transcription. J Biol Chem 275(22): 16459-16465.
Wu, J.I., Lessard, J., Olave, I.A., Qiu, Z., Ghosh, A., Graef, I.A., and Crabtree, G.R. 2007. Regulation of dendritic development by neuron-specific chromatin remodeling complexes. Neuron 56(1): 94-108.
Xiao, L., Kim, M., and DeJong, J. 2006. Developmental and cell type-specific regulation of core promoter transcription factors in germ cells of frogs and mice. Gene Expr Patterns 6(4): 409-419.
Yamit-Hezi, A. and Dikstein, R. 1998. TAFII105 mediates activation of anti-apoptotic genes by NF-kappaB. EMBO J 17(17): 5161-5169.
Yan, Z., Cui, K., Murray, D.M., Ling, C., Xue, Y., Gerstein, A., Parsons, R., Zhao, K., and Wang, W. 2005. PBAF chromatin-remodeling complex requires a novel specificity subunit, BAF200, to regulate expression of selective interferon-responsive genes. Genes Dev 19(14): 1662-1667.

Chapter 3: TRF3, TAF7l, and TAF13 in Hepatic Gene Regulation
Yan, Z., Wang, Z., Sharova, L., Sharov, A.A., Ling, C., Piao, Y., Aiba, K., Matoba, R., Wang, W., and Ko, M.S. 2008. BAF250B-associated SWI/SNF chromatin-remodeling complex is required to maintain undifferentiated mouse embryonic stem cells. Stem Cells 26(5): 1155-1165.
Yang, Y., Cao, J., Huang, L., Fang, H.Y., and Sheng, H.Z. 2006. Regulated expression of TATA-binding protein-related factor 3 (TRF3) during early embryogenesis. Cell Res 16(7): 610-621.
Yoo, A.S., Staahl, B.T., Chen, L., and Crabtree, G.R. 2009. MicroRNA-mediated switching of chromatin-remodelling complexes in neural development. Nature 460(7255): 642-646.
Zhang, D., Penttila, T.L., Morris, P.L., Teichmann, M., and Roeder, R.G. 2001. Spermiogenesis deficiency in mice lacking the Trf2 gene. Science 292(5519): 1153-1155.
Zhou, R., Bonneaud, N., Yuan, C.X., de Santa Barbara, P., Boizet, B., Schomber, T., Scherer, G., Roeder, R.G., Poulat, F., and Berta, P. 2002. SOX9 interacts with a component of the human thyroid hormone receptor-associated protein complex. Nucleic Acids Res 30(14): 3245-3252.


















