
A Comparative Study of Glycopeptides Derived from … the isolation of a low molecular weight...
8
THE JOURNAL OF BIOLOGICAL CHE~IISTRY Vol.245, No. 19,Issue of October 10,~~. 5042-5048, 1970 Printed in U.S.A. A Comparative Study of Glycopeptides Derived from Selected Vertebrate Collagens A POSSIBLE ROLE OF THE CARBOHYDRATE IN FIBRIL lTORMATION* (Received for publication, May 22, 1970) PAUL H. MORGAN, H. GAYLE JACOBS, JERE P. SEGREST, AND LEON W. CUNNINGHAM From the Department of Biochemistry, Vanderbilt University School of Medicine, Nashville, Tennessee 37203 SUMMARY Glycopeptides from carp swim bladder, guinea pig skin, and human skin collagens were prepared by sequential colla- genase-trypsin digestion. Gel filtration on Sephadex G-25 permitted the isolation of a low molecular weight fraction con- taining most of the hydroxylysyl glycosides present in the parent collagen. Application of this fraction to a Dowex 50X-8 ion exchange column consistently resulted in distinc- tive glycopeptide patterns characteristic of each collagen. The glycohexapeptide, Gly-Met-Hyl (Gal-Glc)-Gly-His- Arg, which had been previously characterized in digests of soluble guinea pig skin collagen, was found in significant quantities in all three collagens examined, but the carp and human collagens each revealed an additional major glycopep- tide. The subtractive Edman procedure was used in estab- lishing amino acid sequences, and alkaline hydrolysis was used to confirm the composition and mode of attachment of the carbohydrate side chains. The predominant glycopep- tide isolated from carp swim bladder collagen was Gly-Ile- Hyl(Gal-Glc)-Gly-His-Arg, while the structure of the unique glycopeptide isolated from human skin collagen was Gly-Phe-Hyl(Gal-Glc)-Gly-lle-Arg. A possible role of the carbohydrate side chains in the or- ganization of fibrils is discussed. The biological function of carbohydrate prosthetic groups of glycoproteins has received much attention. It has been postu- lated, for example, that the rather large carbohydrate side chains of these proteins serve, in some way, to regulate their secretion (1, 2). Soluble collagen and salivary mucins (3) are, however, somewhat unique because of the relatively small molecular size of their carbohydrate moieties. We have previ- * This investigation was supported by Grant AM 11821, Train- ine Grant GM 00300. Health Sciences Advancement Award FR 06&7 from the National Institutes of Health, United States Public Health Service, and Grant GB 12940 from the National Science Foundation. This work was taken in part from the thesis submitted by Paul H. Morgan to the Graduate School of Vander- bilt University in partial fulfillment of the requirements for the degree of Doctor of Philosophy. ously shown that most of the hexose prosthetic groups of soluble collagen from guinea pig skin are di- and monosaccharides bound through an 0-glycosidic linkage to hydroxylysine (4). Two 0-hydroxylysyl glycosides, a disaccharide, Hyl-Gal-Glc, and a monosaccharide, Hyl-Gal, were isolated from alkaline hydroly- sates of soluble guinea pig skin collagen as well as directly from normal human urine. Butler and Cunningham (5), utilizing protease digests of soluble guinea pig skin collagen, have shown that a substantial portion of the disaccharide side chain is linked to hydroxylysine in the sequence, Gly-Met-Hyl(Gal-Glc)-Gly- His-Arg. Subsequently, Butler (6) demonstrated the presence of an identical glycopeptide region in rat skin collagen. The fact that methionine, histidine, and hydroxylysine are present in very small amounts in collagen, yet occur in glycopeptides from collagen of two different species of mammals, suggested that there might exist a high degree of specificity in the enzyme- catalyzed process leading to the hydroxylation and subsequent glycosylation of the hydroxylysine. In the present communication we present results of experi- ments designed to examine the possible relationship of this unusual sequence to fibril formation and the maturation of collagen by characterizing t,he degree of variation of the pre- dominant collagen glycopeptide sequence over a limited phylo- genetic range. Glycopeptides from collagen digests of two additional and phylogenetically distant species, the human and the carp, have therefore been prepared and studied. EXPERIMENTAL PROCEDURE Materials Icht’hyocol was prepared from carp swim bladders by the method of Gallop and Seifter (7). Insoluble collagen was obtained from guinea pig skin and adult human skin by a previ- ously described modification (3) of the method of Veis, Anesey, and Cohen (8). Purified Clostridium histolyticum collagenase, trypsin, Galacto- stat, and Glucostat were purchased from Worthington. Dowex 50X-8 (Spinco type 15A) was obtained from the Spinco Division of Beckman Instruments. Sephadex gels were purchased from Pharmacia and Bio-Gel P-2 from Bio-Rad. Orcinol and spectral quality pyridine (for use in the Edman degradation) were pur- chased from Matheson, Coleman, and Bell; phenylisothiocyanate and trifluoroacetic acid were from Eastman; ninhydrin and 5042 by guest on June 12, 2018 http://www.jbc.org/ Downloaded from
Transcript of A Comparative Study of Glycopeptides Derived from … the isolation of a low molecular weight...

THE JOURNAL OF BIOLOGICAL CHE~IISTRY Vol.245, No. 19,Issue of October 10,~~. 5042-5048, 1970
Printed in U.S.A.
A Comparative Study of Glycopeptides Derived from
Selected Vertebrate Collagens
A POSSIBLE ROLE OF THE CARBOHYDRATE IN FIBRIL lTORMATION*
(Received for publication, May 22, 1970)
PAUL H. MORGAN, H. GAYLE JACOBS, JERE P. SEGREST, AND LEON W. CUNNINGHAM
From the Department of Biochemistry, Vanderbilt University School of Medicine, Nashville, Tennessee 37203
SUMMARY
Glycopeptides from carp swim bladder, guinea pig skin, and human skin collagens were prepared by sequential colla- genase-trypsin digestion. Gel filtration on Sephadex G-25 permitted the isolation of a low molecular weight fraction con- taining most of the hydroxylysyl glycosides present in the parent collagen. Application of this fraction to a Dowex 50X-8 ion exchange column consistently resulted in distinc- tive glycopeptide patterns characteristic of each collagen.
The glycohexapeptide, Gly-Met-Hyl (Gal-Glc)-Gly-His- Arg, which had been previously characterized in digests of soluble guinea pig skin collagen, was found in significant quantities in all three collagens examined, but the carp and human collagens each revealed an additional major glycopep- tide. The subtractive Edman procedure was used in estab- lishing amino acid sequences, and alkaline hydrolysis was used to confirm the composition and mode of attachment of the carbohydrate side chains. The predominant glycopep- tide isolated from carp swim bladder collagen was Gly-Ile- Hyl(Gal-Glc)-Gly-His-Arg, while the structure of the unique glycopeptide isolated from human skin collagen was Gly-Phe-Hyl(Gal-Glc)-Gly-lle-Arg.
A possible role of the carbohydrate side chains in the or- ganization of fibrils is discussed.
The biological function of carbohydrate prosthetic groups of glycoproteins has received much attention. It has been postu- lated, for example, that the rather large carbohydrate side chains of these proteins serve, in some way, to regulate their secretion (1, 2). Soluble collagen and salivary mucins (3) are, however, somewhat unique because of the relatively small molecular size of their carbohydrate moieties. We have previ-
* This investigation was supported by Grant AM 11821, Train- ine Grant GM 00300. Health Sciences Advancement Award FR 06&7 from the National Institutes of Health, United States Public Health Service, and Grant GB 12940 from the National Science Foundation. This work was taken in part from the thesis submitted by Paul H. Morgan to the Graduate School of Vander- bilt University in partial fulfillment of the requirements for the degree of Doctor of Philosophy.
ously shown that most of the hexose prosthetic groups of soluble collagen from guinea pig skin are di- and monosaccharides bound through an 0-glycosidic linkage to hydroxylysine (4). Two 0-hydroxylysyl glycosides, a disaccharide, Hyl-Gal-Glc, and a monosaccharide, Hyl-Gal, were isolated from alkaline hydroly- sates of soluble guinea pig skin collagen as well as directly from normal human urine. Butler and Cunningham (5), utilizing protease digests of soluble guinea pig skin collagen, have shown that a substantial portion of the disaccharide side chain is linked to hydroxylysine in the sequence, Gly-Met-Hyl(Gal-Glc)-Gly- His-Arg. Subsequently, Butler (6) demonstrated the presence of an identical glycopeptide region in rat skin collagen. The fact that methionine, histidine, and hydroxylysine are present in very small amounts in collagen, yet occur in glycopeptides from collagen of two different species of mammals, suggested that there might exist a high degree of specificity in the enzyme- catalyzed process leading to the hydroxylation and subsequent glycosylation of the hydroxylysine.
In the present communication we present results of experi- ments designed to examine the possible relationship of this unusual sequence to fibril formation and the maturation of collagen by characterizing t,he degree of variation of the pre- dominant collagen glycopeptide sequence over a limited phylo- genetic range. Glycopeptides from collagen digests of two additional and phylogenetically distant species, the human and the carp, have therefore been prepared and studied.
EXPERIMENTAL PROCEDURE
Materials
Icht’hyocol was prepared from carp swim bladders by the method of Gallop and Seifter (7). Insoluble collagen was obtained from guinea pig skin and adult human skin by a previ- ously described modification (3) of the method of Veis, Anesey, and Cohen (8).
Purified Clostridium histolyticum collagenase, trypsin, Galacto- stat, and Glucostat were purchased from Worthington. Dowex 50X-8 (Spinco type 15A) was obtained from the Spinco Division of Beckman Instruments. Sephadex gels were purchased from Pharmacia and Bio-Gel P-2 from Bio-Rad. Orcinol and spectral quality pyridine (for use in the Edman degradation) were pur- chased from Matheson, Coleman, and Bell; phenylisothiocyanate and trifluoroacetic acid were from Eastman; ninhydrin and
5042
by guest on June 12, 2018http://w
ww
.jbc.org/D
ownloaded from

Issue of October 10, 1970 Morgan, Jacobs, Segrest, and Cunningham 5043
UL( +)-allo-&hydroxylysine hydrochloride was from Mann; triethylamine was from Pierce; and spectral grade butyl acetate was from Fischer.
Methods
Carbohydrate Analyses-The hexose content of column effluents was routinely analyzed by the automated orcinol procedure (9). Insoluble materials were manually analyzed for hexose by a slight modification of the method of Weimer and Moshin (10).
Glucose and galactose were measured by use of the Glucostat and Galactostat reagents (Worthington) as previously described (5). Following mild acid hydrolysis (4) of glycopeptides, glucose and galactose were identified by paper chromatography (1-butanol-pyridine-water, 6:4:3) with detection by the aniline- phthalate spray described by Partridge (11).
Amino Scid and Peptide Analyses-The presence of peptides in column effluents was detected by the ninhydrin procedure of Moore and Stein (12) adapted for the Technicon *4utoAnalyzer. Quantitative amino acid analyses (13) were obtained with the model 120 Beckman-Spinco automatic amino acid analyzer. Acid hydrolysis for amino acid determination was carried out in 6 N HCl at 110” for 20 hours.
Preparation of Glycopeptides-Each collagen was submitted to sequential 24.hour collagenase and trypsin digestions, a procedure designed to yield a reproducible preparation of low molecular weight glycopeptides. This procedure has been described in detail in a previous report from this laboratory (4).
Fractionation of Glycopeptides-Following t.he removal of a small amount of insoluble material by centrifugation, the colla- genase-trypsin digests were submitted to gel filtration on a Sephadex G-25 column (5 x 92 cm), equilibrated, and eluted with 0.1 M pyridine acetate buffer, pH 5.0.
The low molecular weight glycopeptide fraction (Pool II) was lyophilized and further fractionated on a column, 0.9 x 21 cm, of Dowex 50X-8 (Spinco type 15A) which had been equilibrated with 0.02 M pyridine acetate buffer, pH 3.1. Column elution proceeded sequentially with: 130 ml of 0.02 M pyridine acetate, pH 3.10; 200 ml of a linear gradient between 100 ml each of 0.02 M pyridine acetate, pH 3.10, and 0.02 M pyridine acetate, pH 3.10; 500 ml of a linear gradient between 250 ml each of 0.20 M
pyridine acetate, pH 3.10, and 1.40 M pyridine acetate, pH 4.4. In all cases, the molarities of pyridine acetate buffers refer to the concentration of pyridine. The column was pumped at a constant rate of 60 ml per hour. The column temperature was 55”; 2-ml fractions were collected.
Amino Acid Sequence Determination-The small quantities of purified glycopeptides available made the subtractive Edman procedure the method of choice for amino acid sequence deter- mination. The procedure of Dopheide, Moore, and Stein (14) was used with some minor modifications suggested by Dr. Stan- ford Moore. Briefly, this procedure involved drying 0.2 pmole of purified glycopeptide in a 12-ml calibrated conical centrifuge tube. The peptide was then dissolved in 0.30 ml of a solution containing SOY0 pyridine and 3% triethylamine. After the addition of 20 ~1 of phenylisothiocyanate (redistilled before use) the tube was thoroughly purged with nitrogen, covered with Parafilm, wrapped in aluminum foil, and maintained at 37” for 2 hours with occasional shaking. After lyophilization over- night, the dry residue was dissolved in 200 ~1 of trifluoroacetic acid, flushed with nitrogen, covered with Parafilm, and main-
tained at 37” for 30 min. Following lyophilization, the residue was dissolved in 0.5 ml of 0.40 M acetic acid and then extracted three times with 0.5 ml of butyl acetate. A suitable aliquot of the aqueous phase was removed for amino acid analysis while the remainder was lyophilized and then submitted to the next cycle of degradation.
Alkaline Hydrolysis of Ichthyocol, Guinea Pig Skin, and Human Skin Collagens and Glycopeptides-A lOO-mg sample of each collagen was hydrolyzed in 7 ml of 3 N NaOH for 16 hours at 90” in Vycor tubes. After centrifugation the hydrolysates were desalted on a Sephadex G-10 column (2.5 x 45 cm), equilibrated, and eluted with 0.1 M pyridine acetate, pH 5.0. The desalted sample was applied to a column, 0.9 x 21 cm, of Dowex 50X-8 (Spinco type 15A) which had been equilibrated with 0.24 N so- dium citrate, pH 3.25. After eluting with 60 ml of the starting buffer a change to 0.20 N sodium citrate, pH 4.25, was made. Normalities of sodium citrate buffers refer to sodium ion con- centration. Both buffers were 0.1 M in citrate and the pH was adjusted with HCl. The column temperature was maintained at 55” with a flow rate of 60 ml per hour. One-milliliter frac- tions were collected and analyzed for hexose. The column was calibrated with authentic samples of Hyl-Gal-Glc and Hyl-Gal which eluted at 58 ml and 106 ml, respectively, under these conditions.
Glycopeptides from human collagen (Peaks II, III, and V) were hydrolyzed for 10 hours in 2 N NaOH at 90” at a concen- tration of approximately 0.075 mg of hexose per ml. After neutralization the hydrolysates were desalted and submitted to Dowex chromatography as described above.
RESULTS
Fractionation of Glycopeptides
Collagens derived from carp swim bladder, guinea pig skin, and human skin were treated with collagenase and trypsin and the digests were submitted to gel filtration on Sephadex G-25 as indicated in Fig. 1. In each case the low molecular weight
FRACTION NUMBER
FIG. 1. Fractionation on Sephadex G-25 of peptides produced by sequential collagenase and trypsin digestion of (A) ichthyocol, (B) guinea pig skin-insoluble collagen, and (C) human skin-in- soluble collagen. The eluent was 0.1 M pyridine acetate, pH 5.0; 25-ml fractions were collected. --, hexose, reported as galac- tose equivalents; - - -, peptide, determined as absorbance at 570 rnp by the ninhydrin reaction.
by guest on June 12, 2018http://w
ww
.jbc.org/D
ownloaded from
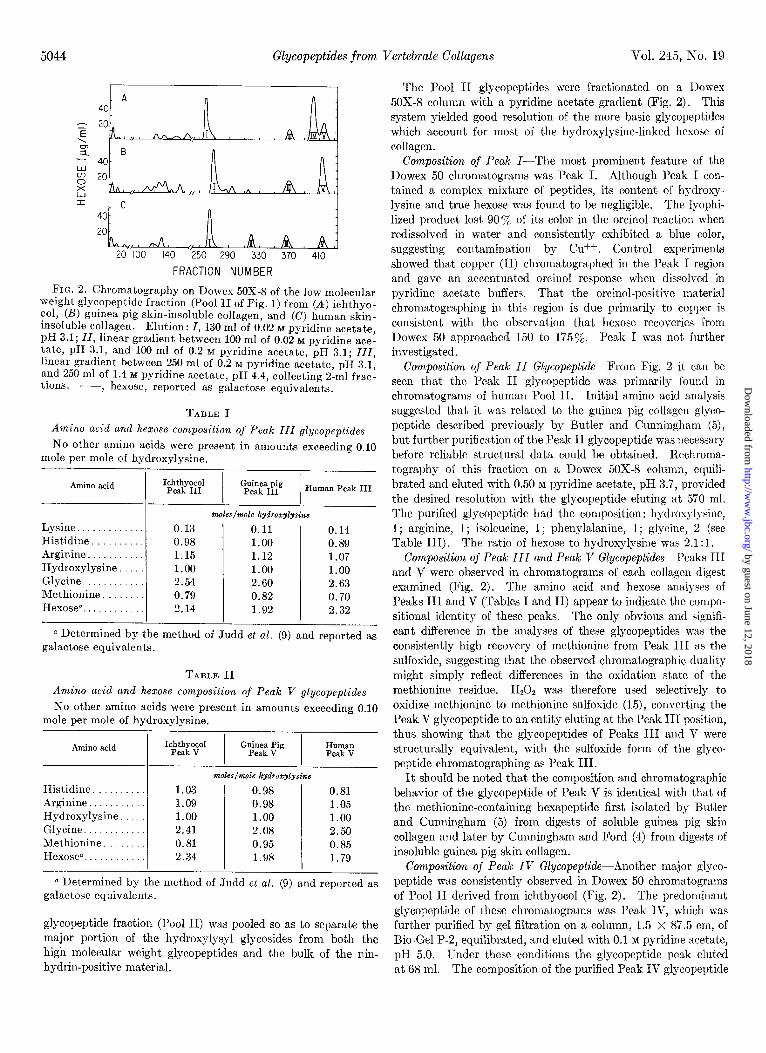
5044 Glycopeptides from Vertebrate Collagens Vol. 245, No. 19
20 100 140 250 290 330 370 410
FRACTION NUMBER
FIG. 2. Chromatography on Dowex 50X-8 of the low molecular weight glycopeptide fraction (l’ool II of Fig. 1) from (A) ichthyo- col, (B) guinea pig skin-insoluble collagen, and (C) human skin- insoluble collagen. Elution: I, 130 ml of 0.02 M pyridine acetate, pH 3.1; ZZ, linear gradient between 100 ml of 0.02 M pyridine ace- tate, pH 3.1, and 100 ml of 0.2 M pyridine acetate, pH 3.1; ZZZ, linear gradient between 250 ml of 0.2 M pyridine acetate, pH 3.1, and 250 ml of 1.4 M pyridine acetate, pH 4.4, collecting 2-ml frac- tions . p, hexose, reported as galactose equivalents.
TABLE I
Amino acid and hexose composition of Peak ZZZ glycopeptides
No other amino acids were present in amounts exceeding 0.10 mole per mole of hydroxylysine.
Lysine Histidine Arginine . Hydroxylysine Glycine Methionine . Hexosea.
I I
a Determined by the method of Judd et al. (9) and reported as galactose equivalents.
0.13 0.98 1.15 1.00 2.54 0.79 2.14
I I
moles/mole hydroxylysinc
0.11 0.14 1.00 0.89 1.12 1.07 1.00 1.00 2.60 2.63 0.82 0.70 1.92 2.32
TABLE II
Amino acid and hexose composition of Peak V glycopeptides
No other amino acids were present in amounts exceeding 0.10 mole per mole of hydroxylysine.
Histidine Arginine Hydroxylysine Glycine........... Methionine Hexosea.
moles/mole hydrozylysine
1.03 0.98 0.81 1.09 0.98 1.05 1.00 1.00 1.00 2.41 2.08 2.50 0.81 0.95 0.85 2.34 1.98 1.79
a Determined by the method of Judd et al. (9) and reported as galactose equivalents.
glycopeptide fraction (Pool II) was pooled so as to separate the major portion of the hydroxylysyl glycosides from both the high molecular weight glycopeptides and the bulk of the nin- hydrin-positive material.
The Pool II glycopeptides were fractionated on a 1)owex 50X-8 column with a pyridine acetate gradient (Fig. 2). This system yielded good resolution of the more basic glycopeptides which account for most of the hydroxylysine-linked hexose of collagen.
Composition of Peak Z-The most prominent feature of the Dowex 50 chromatograms was Peak I. Although Peak I con- tained a complex mixture of peptides, its content of hydroxy- lysine and true hexose was found to be negligible. The lyophi-
lized product lost 900/, of its color in the orcinol reaction when redissolved in water and consistently exhibited a blue color, suggesting contamination by Cu++. Control experiments showed that copper (II) chromatographed in the Peak I region and gave an accentuated orcinol response when dissolved in pyridine acetate buffers. That the orcmol-positive material chromatographing in this region is due primarily to copper is consistent with the observation that hexose recoveries from Dowex 50 approached 150 to 175%. Peak I was not further investigated.
Composition of Peak II Glycopeptide-From Fig. 2 it can be seen that the Peak II glycopeptide was primarily found in chromatograms of human Pool II. Initial amino acid analysis
suggested that it was related to the guinea pig collagen glyco- peptide described previously by Butler and Cunningham (5), but further purification of the Peak II glycopeptide was necessary before reliable structural data could be obtained. Rechroma- tography of this fraction on a Dowex 50X-8 column, equili- brated and eluted with 0.50 M pyridine acetate, pH 3.7, provided the desired resolution with the glycopeptide eluting at 570 ml. The purified glycopeptide had the composition: hydroxylysine, 1; arginine, I ; isoleucine, 1; phenylalanine, 1; glycine, 2 (see Table III). The ratio of hexose to hydroxylysine was 2.1 :l.
Composition of Peak III and Peak V Glycopeptides-Peaks III and V were observed in chromatograms of each collagen digest examined (Fig. 2). The amino acid and hexose analyses of Peaks III and V (Tables I and II) appear to indicate the compo- sitional identity of these peaks. The only obvious and signifi- cant difference in the analyses of these glycopeptides was the consistently high recovery of methionine from Peak III as the sulfoxide, suggesting that the observed chromatographic duality might simply reflect differences in the oxidation state of the methionine residue. H,O, was therefore used selectively to oxidize methionine to methionine sulfoxide (15), converting the Peak V glycopeptide to an entity eluting at the Peak III position, thus showing that the glycopeptides of Peaks III and V were structurally equivalent, with the sulfoxide form of the glyco- peptide chromatographing as Peak III.
It should be noted that the composition and chromatographic behavior of the glycopeptide of Peak V is identical with that of the methionine-containing hexapeptide first isolated by Butler and Cunningham (5) from digests of soluble guinea pig skin collagen and later by Cunningham and Ford (4) from digests of insoluble guinea pig skin collagen.
Composition of Peak IV Glycopeptidt-Another major glyco- peptide was consistently observed in Dowex 50 chromatograms of Pool II derived from ichthyocol (Fig. 2). The predominant glycopeptide of these chromatograms was Peak IV, which was further purified by gel filtration on a column, 1.5 x 87.5 cm, of Bio-Gel P-2, equilibrated, and eluted with 0.1 M pyridine acetate, pH 5.0. Under these conditions the glycopeptide peak eluted at 68 ml. The composition of the purified Peak IV glycopeptide
by guest on June 12, 2018http://w
ww
.jbc.org/D
ownloaded from

Issue of October 10, 1970 Morgan, Jacobs, Segrest, and Cunningham 5045
TABLE III
Subtractive Edman degradation of human Peak II glycopeptide
Proposed sequence Gly - Phe - Hyl - Cly - Ileu - Arg Composition” 1.14 1.04 0.95 1.14 0.99 1.00 First cycle - 1.14 - 1.10 1.00 1.00 Second cycle - 1.10 1.00 1.00 Third cycle - 1.17 1.00 1.00 Fourth cycle - 0.49 0.96 1.00 Fifth cycle - - - 0.60 0.60 1.00
a Composition is reported relative to arginine which is assumed to be unity. Boldface type indicates the residue removed in the corresponding cycle of degradation. No ot,her amino acids were present in quantities exceeding 0.10 mole per mole of arginine. Dashes (-) indicate <0.2 mole per mole of arginine.
was hydroxylysine, 1; histidine, 1; arginine, 1; glycine, 2; iso- leucine, 1 (see Table IV), with a hexose-hydroxylysine ratio of 1.8:1.
Amino Acid Xeguence of Peak II Glycopeptide-All of the major peptides obtained in this study contain 1 arginine residue which may tentatively be assigned a COOH-terminal position on the basis of the well defined specificity of trypsin. The sub- tractive Edman procedure was applied to the phenylalanine- containing glycohexapeptide (Peak II) obtained from human skin collagen yielding the results shown in Table III. The initial composition of the human Peak II glycopeptide and its proposed sequence are included for reference. The first degra- dative cycle resulted in the loss of 1 residue of glycine and also 1 residue of hydroxylysine. Phenylalanine was completely removed in the second cycle, while no losses were apparent during the third cycle. After this step, fractional losses were observed, respectively, in glycine and isoleucine relative to arginine.
Glycine is assigned an NH&erminal position despite the con- comitant first cycle loss of hydroxylysine. Phenylisothiocyanate appears to couple with the e-amino group of hydroxylysine, resulting in the apparent loss of a nonterminal residue through the formation of the acid-stable e-amino phenylthiocarbamyl derivative. Thus, the position of hydroxylysine would be indicated by a subsequent cycle in which no residue loss is appar- ent. This phenomenon is observed in the third cycle of the degradation of both the Peak II and Peak IV glycopeptides. It would appear that during this cycle the hydroxylysyl residue is actually removed by the coupling and cyclization of phenyl- isothiocyanat’e via the or-amino group. These analyses, despite fractional losses observed in the fourth and fifth cycles, allow unambiguous assignment of both hexapeptide sequences. These results are consistent with those that one would expect for a collagen peptide (glycine in every third position) obtained by a procedure which includes trypsin digestion. Glycosylation of hydroxylysyl residues posed no problem in the application or interpretation of the Edman procedure. It is of interest, how- ever, to note that the bond involving glycosylated hydroxylysine is resistant to trypsin, although Butler (16) has presented evi- dence that other hydroxylysyl bonds in collagen are cleaved by trypsin.
Amino Acid Sequence of Glycopeptide of Peaks III and V
It has already been noted that the composition of the glyco- peptide chromatographing as Peaks III and V is identical with that of the glycohexapeptide first reported by Butler and Cun-
TABLE IV
Subtractive Edman degradation of ichthyocol Peak IV glycopeptide
Proposed sequence Cly - Ile - 11~1 - Gly - His - Arg Composition” 1.11 1.02 1.04 1.11 1.01 1.00 First cycle - 0.87 - 1.12 0.96 1.00 Second cycle - 1.34 1.02 1.00 Third cycle - 1.20 1.04 1.00 Fourth cycle - - 0.66 1.05 1.00 Fift,h cycle - - 0.56 0.49 1.00
a Composition is reported relative to arginine which is assumed to be unity. Boldface type indicates t.he residue removed in the corresponding cycle of degradation. No other amino acids were present in quantities exceeding 0.10 mole per mole of aryi- nine. Dashes (-) indicate <0.2 mole per mole of arginine.
ningham (5) in digests of soluble guinea pig skin collagen. The weight of chromatographic and compositional data makes it highly probable that the sequence reported by these authors, Gly-Met-Hyl(Gal-Glcj-Gly-His-Arg, is common to the glyco- peptides of carp, guinea pig, and human Peaks III and V. Pre- liminary sequence studies, with the subtractive Edman procedure, confirm the NHz-terminal sequence, Gly-Met, in the human Peak V glycopeptide.
Amino Acid Sequence of Peak IV Glycopeptide-Results of the Edman degradation of ichthyocol Peak IV are given in Table IV. ,4s in the case of human Peak II, the first cycle resulted in the loss of 1 residue of glycine and 1 residue of hy- droxylysine. Isoleucine was lost in the second cycle and no losses were observed during the third cycle. Beyond this point losses were again fractional.
Identi$cation of Hydroxylysine-linked Hexoses of Carp, Guinea
Pig, and IIuman Collagens and Glycopeptides-The 2 : 1 ratio of hexose to hydroxylysine in each of the glycopeptides mentioned in this study suggests that the carbohydrate linkage and structure are identical with the Hyl-Gal-Glu found for the glycohexa- peptide from guinea pig skin collagen (4). Because of the known stability of these glycosides to alkaline hydrolysis, samples of the whole collagens and selected glycopeptides were submitted to this treatment in order to obtain the free 0-hydroxylysyl glycosides. The reproducible chromatographic resolution of Hyl-Gal-Glu and Hyl-Gal on Dowex 50X-8 permitted the identification of these compounds in the alkaline hydrolysates. Application of these procedures to the whole collagens indicated the presence of both Hyl-Gal-Glc and Hyl-Glc with the disac- charide constituting 88, 70, and 62% of the total hexose-linked hydroxylysine in carp, guinea pig, and human collagenn, respec- tively. The value for carp agrees well with that reported by Spiro (17).
Paper chromatography of acid hydrolysates of the Peak IV glycopeptide from ichthyocol indicated the presence of glucose and galactose as the only hexoses and in approximately equal amounts. These results, coupled with the knowledge that 88% of the hexose-linked hydroxylysine of ichthyocol is present as the disaccharide, indicate that the structure of the carbohydrate moiety of Peak IV is 2-O-ol-n-glucosyl-o-fi-n-galactosylhydroxy- lysine (17).
The human Peaks II, III, and V were subjected to alkaline hydrolysis, and Dowex 50 chromatography indicates the result- ing hydroxylysyl glycoside to be Hyl-Gal-Glu in each case. Mild acid hydrolysis of the Peak II glycopeptide followed by
by guest on June 12, 2018http://w
ww
.jbc.org/D
ownloaded from

5046 Glycopeptides from Vertebrate Collagen.3
glucose and galactose oxidase determinations confirmed the presence of equimolar quantities of glucose, galactose, and hydroxylysine.
have the reverse requirements. Further, the kidney galactosyl- transferase adds galactose to unsubstituted hydroxylysine of native collagen, but this reaction is not mediated by the con- nective tissue galactosyltransferase.
DISCUSSION
The glycopeptides of Peaks II, III, IV, and V all possess a hydroxylysine-linked disaccharide. The Hyl-Gal-Glc nucleus, first reported by Butler and Cunningham (5) in guinea pig skin collagen, is now recognized as a structural feature common to a wide variety of vertebrate collagens, although its functional role remains unclear. Cunningham, Ford, and Segrest (18) have
reported the isolation of t’his same hydroxylysine disaccharide directly from normal human urine. Spiro (19) has chemically characterized the Hyl-Gal-Glc unit derived from glomerular basement membrane and has deduced its structure to be 2-0-a-~- glucosyl-O-fi-n-galactosylhgdroxylysine. On the basis of similar work, the hydroxylysine disaccharides of ichthyocol (17) and lens capsule (20) were subsequently reported to have an identical structure. Since we have now shown the chromatographic identity of the Hyl-Gal-Glc units derived from carp, guinea pig, and human collagens, it seems highly probable that the carbo- hydrate side chains of Peaks II, III, IV, and V have the detailed structure described by Spiro. The three disaccharide glyco- peptide sequences now known are summarized in Table V. A general sequence can also be given: Gly-Y-Hyl(Gal-Glc)-Gly- Z-Arg. Residues Y and Z, respectively, represent: methionine and histidine in the glycopeptide of Peaks III and V from all three species; isoleucine and histidine in the ichthyocol Peak IV glycopeptide; phenylalanine and isoleucine in the human Peak II glycopeptide. It is possible that the invariant arrangement of hydroxylysine and arginine at positions 3 and 6 may represent an absolute requirement for glycosylation (i.e. the minimal sequence or conformation of the collagen chain (or both) consist- ent with the specificity of galactosyl or glucosyl transferase (or both)). Evidently, specificity requirements are less stringent for residues Y and Z, although it is likely that some restrictions apply since no acidic residues have been observed in either of these positions in skin collagens or ichthyocol. It should be recalled, however, that the compositions of the glycopeptides from glomerular basement membrane containing an identical hydroxylysine-disaccharide moiety suggest that, in that protein, glutamic and aspartic acids may occupy positions Y or Z (21).
Recognizing these differences in the glycosyltransferases isolated from different sources, one may tentatively analyze the specificity of enzymes from connective tissue. It would appear that the glucosyl- and galactosyltransferases are regulated by entirely separa.te specificity requirements. The hydroxylgsine- galactoside can be synthesized even with free hydroxylysine as substrate, but some of the hydroxylysines of collagen are appar- ently located in a sequence which will not permit the enzyme to act (23). On the other hand, the glucosyltransferase fails to recognize either native collagen or free galactose but requires a collagen-derived “receptor” lacking glucose (22). These findings have led to the assumption that the glucosyl transferase requires a special amino acid sequence, although it is not certain whether the hydroxylysine-galactoside alone might not also serve as substrate. Further investigation is needed in order to determine the absolute specificities of these two enzymes with regard to the peptide portion adjacent to “receptive” hydroxylysines of collagens.
Preliminary analyses of a monosaccharide glycopeptide isolated from collagenase-trypsin digests of human skin collagen indicate that it has the amino acid sequence: Gly-Pro-Hyl.1 This finding suggests that an imino acid residue at Y or Z (or both) may be incompatible with the specificity of skin glucosyltrans- ferase, thus restricting glycosylation at such sequences to mono- saccharide side chains. Cleavage by collagenase COOH-termi- nal to the hydroxylysine residue in this type of sequence (-Gly- Pro-X-Gly-) would appear to be consistent with the known specificity of bacterial collagenase (25) and furthermore would provide an explanation for the relatively smaller size of mono- saccharide glycopeptides first observed by Spiro (17).
The specificities of galactosyl- and glucosyltransferases have been studied with enzymes prepared from embryonic guinea pig skin and cartilage (22,23) and from rat kidney cortex (24). The transferases isolated from the former two sources exhibit signifi- cantly different properties from those found in kidney. For instance, with preparations from skin and cartilage, free hy- droxylysine acts as receptor for galactose, and glucosyltrans- ferase requires a specific galactosyl-hydroxylysine-peptide or tertiary structure, while the analogous enzymes from kidney
T~DLE V
Summary of major glycopeptide structures
Peak II Human Peaks III I%uman, guinea
and V pig, carp Peak IV Carp
Gly-Phe-Hyl (Gal-Glc)-Gly-Ile-Arg Gly-Met-Hyl(Gal-Glc)-Gly-His-Arg
Although the number of species examined is not yet large, it is impressive that the sequence Gly-Met-Hyl(Gal-Glc)-Gly-His- Arg is found as a major component of the digests of all which have been studied. In rat skin collagen (6) this particular glycosylated hydroxylysine is known to be located in the a-1 chain approximately 10% of the total length of the molecule from the NHz-terminal end. Furthermore, Piez et ~1.2 have recently shown that the location of this segment in the a-1 chain lies opposite the “hole” region (26) between two neighboring collagen molecules in a quarter-stagger array (27) (Fig. 3A).
Since the approximate diameter of a collagen molecule is of the order of 15 A (28) and the center to center distance between any 2 molecules in a hexagonal array of collagen in fibrils is about 17 A (29) (see Fig. 3B), it may be presumed that the projecting disaccharide prosthetic group, measuring at least 7 A by 15 ,4, would in all likelihood prevent or seriously impair regular hexag- onal packing in a quarter-stagger array unless holes existed in appropriate locations. Furthermore, it is known that these collagen molecules contain two a-1 chains, both having a disac- charide unit in the same location (6). If the polypeptide chains begin approximately in register at the NHp-terminal end, then the three-chain helical model requires that regular hexagonal packing of collagen molecules in a quarter-stagger array would be possible only if two hole regions occurred adjacent to each central collagen molecule, at the same short distance from the
Gly-Ile-Hyl (Gal-Glc)-Gly-His-Arg 1 H. G. Jacobs, unpublished data. 2 K. A. Piez et ul., in preparation; quoted by Butler (9).
Vol. 245, n-o. 19
by guest on June 12, 2018http://w
ww
.jbc.org/D
ownloaded from

Issue of October 10, 1970 Morgan, Jacobs, Segrest, and Cunningham
amino-terminal end of the molecule (Fig. 3A), and at an angle of 120’ from each other (Fig. 3B).
The nature of the three-dimensional packing of collagen into fibrils is not clearly understood. Many of the more recent suggestions have been summarized by Ramachandran (29) and by Veis, Spector, and Carmichael (30). It can, however, be shown that there are only three possible regular arrays of hexag- onally packed collagen molecules in which at least one plane exhib- its the quarter-stagger arrangement which is essential for the gen- eration of the typical electron microscopic banding pattern of collagen fibers (31). These arrays are defined by describing the degree of overlap or “stagger” in each of the three contact planes of the fibril: (1) (a) quarter. stagger, (b) half-stagger, (c) quarter- stagger; (2) (a) quarter-stagger, (b) full stagger (SLS), (c) quar- ter-stagger; (3) (a) quarter-stagger, (b) half-stagger, (c) three quarter-stagger.
The growth of a fibril composed of molecules in any of these three arrays can be seen as being initiated by association of 2 collagen molecules in quarter-, half-, or full stagger followed by the association of a 3rd molecule to complete the triangular segment of the developing hexagonal pattern. The pattern for indefinite longitudinal growth of a “minimum” fibril is then set, but growth in thickness of the fibril by hexagonal packing can occur only if the structure of the collagen molecules permits an individual molecule to be completely surrounded by 6 others. \ The steric problem posed by the disaccharide prosthetic group would appear to require that hole regions be appropriately
C &b 126
located before fibril thickness greater than 2 molecules can be FIG. 3. A, quarter-stagger array of collagen molecules along
attained. one nlane of a fibril. The annroximate location of the Glv-Met-
The characteristic pattern of holes surrounding a central Hyl<Gal-Glc)-Gly-His-Arg sequence in the a-1 chains is marked
molecule in each of the three packing arrangements can be l . Hole regions are the unshaded areas. B, diagrammatic representation of an end on view of hexagonally packed collagen
determined. Only the first possibility generates two holes next molecules, including an illustration of the angular distribution of
to a single locus on a common tropocollagen molecule at 120” individual chains (0) at a point within a single molecule, and of
from each other (Fig. 3C). The analysis further predicts that a the triangular array which constitutes the “minimum” fibril
disaccharide unit on the cr-2 chain cannot be located at precisely (shaded). C, diagrammatic representation of the location of
the same locus but could be situated at another locus within one hole regions about an interior collagen molecule in a fibril com- posed of a hexagonally packed, quarter-quarter-half array (see
of these same 400 A holes or at one of four other possible sites, all text).
further toward the COOH-terminal end of the molecule. It also requires that the several minor sites of attachment of hexose, droxylysine residues (including hydroxylation and subsequent mostly as monosaccharide, in these collagen molecules occur at glycosylation of additional lysine residues) would produce colla- these other loci, unless appropriately positioned monosaccharide gen molecules completely incapable of ordered fibril formation units can fit into the interstitial areas between the closely packed such as apparently exists in lens capsule and glomerular mem- molecules (Fig. 3B). brane (17).
It is interesting to consider the possibility that selective It is of interest also that, on the basis of our present knowledge glycosylation of specific hydroxylysine residues might inhibit of amino acid codons (32), the predominant carp sequence, fibril formation or dictate the production of any one of the three Gly-Ile-Hyl(Gal-Glc)-Gly-His-Arg, can be related to the more staggered arrays noted above. An extension of this concept generally found sequence, Gly-Met-Hyl(Gal-Glc)-Gly-His-Arg, coincides with an early observation by Dische (quoted in Refer- by a single base mutation, while the secondary human sequence, ence 3) that a high carbohydrate content was a characteristic of Gly-Phe-Hyl(Gal-Glc)-Gly-Ile-Arg, could only be so related collagen from tissues in which it did not exhibit the characteristic by a much more extensive and rather unlikely series of mutations. banding pattern. Spiro (17) has also noted the decreased A more reasonable hypothesis is that the latter sequence is not morphological organization in collagens having large amounts of related to the others by mutation but occurs in an entirely hydroxylysine-linked hexose. It can be seen that limited different part of the collagen molecule. The several possible glycosylation of “inappropriate” hydroxylysine residues would explanations for these similarities and differences will, however, create collagen molecules which could participate in ordered be susceptible to more detailed analysis only when additional packing only if they were located on the periphery of a fibril. information on the distribution of carbohydrate between the The proportion of such molecules in a population of collagen or-1 and cr-2 chains and the careful mapping of all sites of attach- molecules could be a factor in the control of fibril size in various ment within the chains is available. tissues. It should be emphasized, however, that complete regularity of packing is not to be expected nor required for fiber Acknowledgments-We would like to acknowledge the expert formation. More extensive glycosylation of inappropriate hy- technical assistance of Mr. John Ford and Miss Betty McGloth-
by guest on June 12, 2018http://w
ww
.jbc.org/D
ownloaded from

5048 Glycopeptides from Vertebrate Collagens Vol. 245, No. 19
len ill the perfornmce of these studies. We wish also to :LC- 15. S~:~.LI.\N~, N. I’. iu (:. II. W. IlIils (Editor), .Ilcthocln in enzy-
knowledge ~uluddc discussions with Dr. William T. Uut,ler ,wtolog?y, I’ol. Sl, .lcatlemic l’rcss, Se\\ \‘ork, l!l(ii, p. 485.
concerniilg the role of t.lie cxMlyclratc in coll:igen structure. 16. 13r~r.erc, W. T., ,S<.iencr. 161, 7%; (1968). 17. SPIRO, I!. G ., .I. IIiol. Thou ., 244, (iO2 (I%i!I). 18. C~~NNIS(;ll.\M. I,. W., I~oI~I). J. I.)., .\sI) SIX~ICMT, J. I’., .I. Ijioi.
REFERENCES Chem., 242, 2570 (1967). 19. SPIRO, R. G., J. Biol. Chem., 242, 4813 (1967).
1. EYL.IR, E. H., J. Theoret. Biol., 10, 89 (1965). 20. KEFALIDES, N. A., Proceedings of the Sixth International 2. CUNNINGHAM, L. W., Proceedings of the Fourth International
Conference on Cystic Fibrosis, Berne and Grindelwald. 1966. Congress on Diabetes Mellitus, Stockholm, 1967, Excerpta Medica Foundation. Amsterdam. 1969. n. 307.
3.
4.
5.
0. 7.
8.
9.
10.
11. 12. 13.
14.
Part II, S. Karaer, New York, 1968, p. 141. I I
21. SPIRO, R. G., J. BioZ.‘Chem., 242, 1923 (1967). GRAHAM, E. R. B., ~NII GOTTSCHALK, A., Biochim. Biophys. 22. BOSMANN, H. B., AND EYLAR, E. H., Biochem. Biophys. Res.
Acta, 38, 513 (1960). Commun., 30, 89 (1968). CUNNINGHAM, L. W., AND FORD, ‘J. D., J. Biol. Chem., 243, 23. BOSMANN, H. B.. AND EYLAR. E. H.. Biochem. Biovhus. Res.
2390 (1968) : BUTLER, W. T., ANII CUNNINGHAM, L. W., J. Biol. Chem., 241,
3882 (1966). BUTLER, W. T., Biochemistry, 9, 44 (1969). GALLOP, P. M., AND SEIFTER, S., in S. P. COLO~ICK AND N. 0.
KAPLAN (Editors), Methods in enzymology, VoZ. VI, Aca- demic Press, New York, 1963, p. 635.
VEIS, A., ANESEY, J., AND COHEN, J., J. Amer. Leather Chem. Ass., 66, 548 (1960).
JUDD, J., CLOUSE, W., FORD, J., VAN EYS, J., AND CUNNING- H.4M, L. W., Anal. Biochem., 4, 512 (1962).
WEIMER, H. E.. AND MOSHIN, J. R., Amer. Rev. Tuberc. Pul- monary Dis., 68, 594 (1953):
PARTRIDGE. S. M.. Nature. 164. 443 11949). MOORE, S.,‘AND STEIN, W.‘H., >. B&l. &em., 1’76, 367 (1948). SPACKMAN, D. H., STEIN, W. H., BND MOORE, S., Anal. Chem.,
30, 1190 (1958). DOPHEIDE, T. A. A., MOORE, S., AND STEIN, W. H., J. Biol.
Chem., 242, 1833 (1967).
24. 25.
26.
27.
28.
29.
30.
31.
32.
Commun., 33, 340 (1968). ’ ’ I Y
SPIRO, R. G., BND SPIRO, M. J., Fed. Proc., 27, 345 (1968). GRASSMAN. W.. H~RMANN. H.. NORDWIG. A.. AND WUNSCH.
E., Z. Physidl. Chem., 3i6, 287 (1959). ’ ’ PETRUSKA, J. A., AND HODGE, A. J., Proc. Nat. Acad. Sci.
U. 8. A., 61, 871 (1964). HODGE. A. J., SND SCHMITT. F. 0.. Proc. Nat. Acad. Sci.
U. S: A., 46; 186 (1960). BOEDTKER, H., BND DOTY, P., J. Amer. Chem. Sot., 77, 248
(1955). RAMACHANDHAN, G. N., in G. N. R~MACHANDRAN (Editor),
Treatise on collaaen. Vol. I. Academic Press. New York. 1967, p. 103. ” ’ ’
VEIS, A., SPECTOR, A. R., BND CARMICHAEL, D. J., Clin. or- thop. Related Res., 66, 188 (1969).
HILL, C. E., JAKERS, M. A., AND SCHMITT, F. O., J. Amer. Chem. Sot., 64, 1234 (1942).
SCHWEET, R., AND HEINTZ, R., Annu. Rev. Biochem., 36, 629 (1966).
by guest on June 12, 2018http://w
ww
.jbc.org/D
ownloaded from

Paul H. Morgan, H. Gayle Jacobs, Jere P. Segrest and Leon W. CunninghamFORMATION
Collagens: A POSSIBLE ROLE OF THE CARBOHYDRATE IN FIBRIL A Comparative Study of Glycopeptides Derived from Selected Vertebrate
1970, 245:5042-5048.J. Biol. Chem.
http://www.jbc.org/content/245/19/5042Access the most updated version of this article at
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
http://www.jbc.org/content/245/19/5042.full.html#ref-list-1
This article cites 0 references, 0 of which can be accessed free at
by guest on June 12, 2018http://w
ww
.jbc.org/D
ownloaded from


















