
519 Mechanically Dependent Changes in Action Potentials...
11
519 Mechanically Dependent Changes in Action Potentials Recorded from the Intact Frog Ventricle MAX J. LAB SUMMARY The wall of the ventricle contracts inhomogeneously during an isovolumic beat of an isolated, intact frog ventricle. Some epicardial segments actually lengthen while the pressure is rising. Almost simultaneously, the early repolarization phase of the monophasic action potential recorded from such a segment is accelerated, compared to the same phase for an isotonic beat in which the segment shortens. Segment lengthening during the isovolumic beat also may be seen during the late repolarization phase when, in contrast to the above, it produces an afterdepolari- zation. These electrical changes disappear when isotonic contraction is restored. Corroborative findings were ob- tained from microelectrode and insulated gap recordings from isolated frog ventricular strip. Both electrical changes can be seen clearly when the segment is lengthened by intraventricular injections of Ringer's solution. There also is a short transition period toward the end of the action potential plateau when lengthening produces neither depolari- zation nor repolarization. The accelerated repolarization is manifest as a shortening of the Q-T interval in the ven- tricular electrogram. In all experimental preparations, the afterdepolarizations reached threshold for a propagated action potential. This mechanism may explain the generation of extrasystoles in myocardial ischemia. IT IS WIDELY accepted that during a heart beat there is a progression of events, directionally oriented, which begins with the depolarizing action potential and, via a process of "excitation-contraction coupling," leads to con- traction of the muscle. 1 ' 2 From the observations in this report and elsewhere, 3 " 8 it is suggested that the process is not strictly undirectional. When heart muscle undergoes active or passive mechanical stress or strain, electrical records from the muscle change. Thus the observations on intact frog ventricle presented here, supplemented by studies on isolated ventricular strips, indicate that the mechanics of contraction can influence action potentials and ECG. Furthermore, the mechanism(s) involved can induce opposite polarizing effects, depending on the time of occurrence of an epicardial distortion during the cardiac cycle. These experiments may have some clinical rele- vance in that the mechanically induced changes may alter the T wave repolarizing vector and be related to the generation of ectopic beats in ventricular myocardium. Methods Intact Isolated Ventricle Frog hearts, obtained from animals that were killed by pithing, during winter and summer, were used in the experiments. The ventricular chamber was cannulated and perfused via the aorta. The cannula was connected to a tap constructed to allow isovolumic or free-loaded con- traction, the continuous recording of intraventricular pres- sure, and rapid externally imposed changes in ventricular From the Department of Physiology, Charing Cross Hospital Medical School, Fulham Palace Road, London, England. Presented in part at the XXV International Congress of Physiological Sciences, Munich, 1971. Address for reprints: Max J. Lab, Ph.D. M.B.BCh., Department of Physiology, Charing Cross Hospital Medical School, Fulham Palace Road, London, England W6 8RF. Received September 7, 1976; accepted for publication December 2. 1977. volume. Monophasic action potentials from the epicar- dium and the relative movement of two points on either side of the electrode, monitored by a caliper, were re- corded simultaneously (Fig. 1). Perfusion System The ventricles of frogs (Xenopus laevis) were cannu- lated via the aorta and perfused with continually oxygen- ated Ringer's solution (NaCl, 111.3 ITIM; KC1, 1.8 ITIM; CaCl 2 , 1.08 HIM; Na HCO 3 , 2.4 mM) at room temperature which varied between 20 and 22°C. The cannula was in two sections: a small cannulating section which connected the ventricle to a piston tap and a second vertical section glued to the other end of the tap. A pressure head of 2-3 cm in the cannula provided adequate diastolic filling of the ventricle. The diameter of the ventricle was adjusted by varying the pressure head against which it worked, to be approximately the same as that observed in the intact frog with the chest open. The volume moving up and down the single perfusion cannula with each beat was between 0.1 and 0.3 ml. The contractions were thus free- loaded unless the cannula was closed by the tap, as described below, in which case the contractions were isovolumic. The atria usually were left intact and the heart allowed to beat spontaneously at between 20 and 30 beats/min, but in some cases the atria were tied off and the ventricles electrically stimulated at 24 beats/min. Piston Tap Construction. The piston tap consisted of an outer cylinder with a piston inside. The cannula from the ventricle was inserted into a sealing ring in the wall of the outer cylinder. There were holes running through the piston in different directions so that the position of the piston could determine to which of the resevoirs of fluid the ventricle was connected. There were three basic positions of the tap: for free-loaded (auxotonic) contrac- tion, for isovolumic contraction, and for rapid, externally by guest on June 28, 2018 http://circres.ahajournals.org/ Downloaded from
Transcript of 519 Mechanically Dependent Changes in Action Potentials...

519
Mechanically Dependent Changes in ActionPotentials Recorded from the Intact Frog Ventricle
MAX J. LAB
SUMMARY The wall of the ventricle contracts inhomogeneously during an isovolumic beat of an isolated, intactfrog ventricle. Some epicardial segments actually lengthen while the pressure is rising. Almost simultaneously, theearly repolarization phase of the monophasic action potential recorded from such a segment is accelerated, comparedto the same phase for an isotonic beat in which the segment shortens. Segment lengthening during the isovolumicbeat also may be seen during the late repolarization phase when, in contrast to the above, it produces an afterdepolari-zation. These electrical changes disappear when isotonic contraction is restored. Corroborative findings were ob-tained from microelectrode and insulated gap recordings from isolated frog ventricular strip. Both electrical changescan be seen clearly when the segment is lengthened by intraventricular injections of Ringer's solution. There also isa short transition period toward the end of the action potential plateau when lengthening produces neither depolari-zation nor repolarization. The accelerated repolarization is manifest as a shortening of the Q-T interval in the ven-tricular electrogram. In all experimental preparations, the afterdepolarizations reached threshold for a propagatedaction potential. This mechanism may explain the generation of extrasystoles in myocardial ischemia.
IT IS WIDELY accepted that during a heart beat there isa progression of events, directionally oriented, whichbegins with the depolarizing action potential and, via aprocess of "excitation-contraction coupling," leads to con-traction of the muscle.1'2 From the observations in thisreport and elsewhere,3"8 it is suggested that the process isnot strictly undirectional. When heart muscle undergoesactive or passive mechanical stress or strain, electricalrecords from the muscle change. Thus the observations onintact frog ventricle presented here, supplemented bystudies on isolated ventricular strips, indicate that themechanics of contraction can influence action potentialsand ECG. Furthermore, the mechanism(s) involved caninduce opposite polarizing effects, depending on the timeof occurrence of an epicardial distortion during the cardiaccycle. These experiments may have some clinical rele-vance in that the mechanically induced changes may alterthe T wave repolarizing vector and be related to thegeneration of ectopic beats in ventricular myocardium.
Methods
Intact Isolated Ventricle
Frog hearts, obtained from animals that were killed bypithing, during winter and summer, were used in theexperiments. The ventricular chamber was cannulated andperfused via the aorta. The cannula was connected to atap constructed to allow isovolumic or free-loaded con-traction, the continuous recording of intraventricular pres-sure, and rapid externally imposed changes in ventricular
From the Department of Physiology, Charing Cross Hospital MedicalSchool, Fulham Palace Road, London, England.
Presented in part at the XXV International Congress of PhysiologicalSciences, Munich, 1971.
Address for reprints: Max J. Lab, Ph.D. M.B.BCh., Department ofPhysiology, Charing Cross Hospital Medical School, Fulham Palace Road,London, England W6 8RF.
Received September 7, 1976; accepted for publication December 2.1977.
volume. Monophasic action potentials from the epicar-dium and the relative movement of two points on eitherside of the electrode, monitored by a caliper, were re-corded simultaneously (Fig. 1).
Perfusion System
The ventricles of frogs (Xenopus laevis) were cannu-lated via the aorta and perfused with continually oxygen-ated Ringer's solution (NaCl, 111.3 ITIM; KC1, 1.8 ITIM;CaCl2, 1.08 HIM; Na HCO3, 2.4 mM) at room temperaturewhich varied between 20 and 22°C. The cannula was intwo sections: a small cannulating section which connectedthe ventricle to a piston tap and a second vertical sectionglued to the other end of the tap. A pressure head of 2-3cm in the cannula provided adequate diastolic filling ofthe ventricle. The diameter of the ventricle was adjustedby varying the pressure head against which it worked, tobe approximately the same as that observed in the intactfrog with the chest open. The volume moving up anddown the single perfusion cannula with each beat wasbetween 0.1 and 0.3 ml. The contractions were thus free-loaded unless the cannula was closed by the tap, asdescribed below, in which case the contractions wereisovolumic. The atria usually were left intact and the heartallowed to beat spontaneously at between 20 and 30beats/min, but in some cases the atria were tied off andthe ventricles electrically stimulated at 24 beats/min.
Piston Tap
Construction. The piston tap consisted of an outercylinder with a piston inside. The cannula from theventricle was inserted into a sealing ring in the wall of theouter cylinder. There were holes running through thepiston in different directions so that the position of thepiston could determine to which of the resevoirs of fluidthe ventricle was connected. There were three basicpositions of the tap: for free-loaded (auxotonic) contrac-tion, for isovolumic contraction, and for rapid, externally
by guest on June 28, 2018http://circres.ahajournals.org/
Dow
nloaded from

520 CIRCULATION RESEARCH VOL. 42, No. 4, APRIL 1978
imposed changes in ventricular volume. Ventricular pres-sure could be monitored at any of the positions.
Operation of the Tap. The relevant position of thepiston and thus alignment of the appropriate holes werechosen by setting a mechanical stop onto which the pistonwas driven by gas under pressure. As shown in Figure 1,the three-way solenoid valve releasing the gas underpressure was opened by the closing of gated relay, 2,controlled by a "Digitimer" (Devices). The piston wasspring-loaded and returned, aided by vacuum, when thevalve switched back. The "Digitimer" was triggered by aSchmitt Trigger (B) into which the action potential wasfed.
Injecting System
Intraventricular injections and withdrawals were accom-plished by a motor-driven syringe which was operatedwhen another relay, 1, closed at times preset by the"Digitimer." The relay allowed current to flow first in onethen in the other direction to drive the motor forward andthen in reverse.
Recording System
Monophasic action potentials from the epicardium wererecorded by means of suction electrodes (C) after themethod of Hoffman, et al.9 These authors have drawnattention to some of the pitfalls in the interpretation ofthe records obtained by this method of recording. How-ever, interest in the present studies is centered on the
A — »•!'
H
FIGURE 1 Diagram of experimental arrangement. The frog ven-tricle was cannulated and attached to the cylinder of the piston. Themotor driven syringe was operated by Relay 1 and a "Digitimer."A three-way solenoid valve was connected to a pressure cylinderoperated by Relay 2. A, vacuum; B, Schmidt-trigger; C, suctionelectrode; D, caliper; E, electrocardiogram; F, pressure transducer;G, photoelectric circuit to monitor plunger of syringe;' H, multi-channel pen recorder; I, storage oscilloscope; J, marker.
slower time course changes, for example, the repolariza-tion phase of the monophasic action potential which is notqualitatively different from the repolarization phase foundin transmembrane action potentials recorded through amicroelectrode.9 The suction electrode recording was fedinto one input of a high input impedance differentialamplifier. The other input was a wick electrode on theventricle. The precise location of the wick did not affectthe nature of the recording obtained.
The relative movements of two points on either side ofthe electrode were monitored by a photoelectric caliper(D) placed either longitudinally or transversely. Thiscaliper was also attached by suction and recorded roughlyunidimensional changes in the chord length of the segmentstudied.10 The construction of the caliper did not allow for"shearing" between the points of attachment of the caliperto the epicardium. A bipolar electrogram (E, Fig. 1) wasrecorded from two epicardial wick electrodes, soaked inRinger's solution, or with one electrode in the intraven-tricular perfusing fluid. Intraventricular pressure wasmonitored with a pressure transducer (F), and the volumesinjected into the ventricle were monitored by means of agraded density film attached to the plunger of the motor-driven syringe. The film interrupted a light source to aphototransistor (G) ." The signals from transducers andelectrodes were fed into a multichannel pen recorder (H)(Devices) and storage oscilloscope (I) (Tektronix).
Confirmatory Records from Frog Ventricular Strips
lntracellular Recordings
A ring of muscle was cut from the base of the frogventricles (Rana pipiens), connective and atrial tissueswere cut away, and the ring was converted into a stripabout 1 mm2 and 4-8 mm long. The muscle was clampedhorizontally in a perfusion chamber and a conventionalmicroelectrode recording system for cardiac muscle wasused.12 The electrodes were floating free from earth andtheir resistances were not less than 20 Mft. Records of allthe attempted impalements were stored on magnetic tapeso that those which were successful could be analyzedlater. The number of successful impalements was ratherlow because frog ventricular cells are small and the musclelength was changing. Tension was measured by an RCAtransducer.
Insulated Cap Recordings
The method described by Niedergerke13 was used forthese recordings. The strip of ventricle, prepared asdescribed above, was mounted vertically and clampednear one end in a canal 0.5 mm long and 0.5 mm indiameter. The canal was formed by two sliding plates ofacrylicic plastic which created a partition between twochambers. Silicone grease in and around the canal com-pleted the separation between the two chambers. Themuscle traversed the gap and lay in the chambers; thefirst chamber containing Ringer's solution and the secondisotonic KCL or 0 .5% procaine in 7% sucrose. Actionpotentials recorded across the insulating gap, between thefirst and second chambers, were 20-60 mV in amplitude.14
by guest on June 28, 2018http://circres.ahajournals.org/
Dow
nloaded from
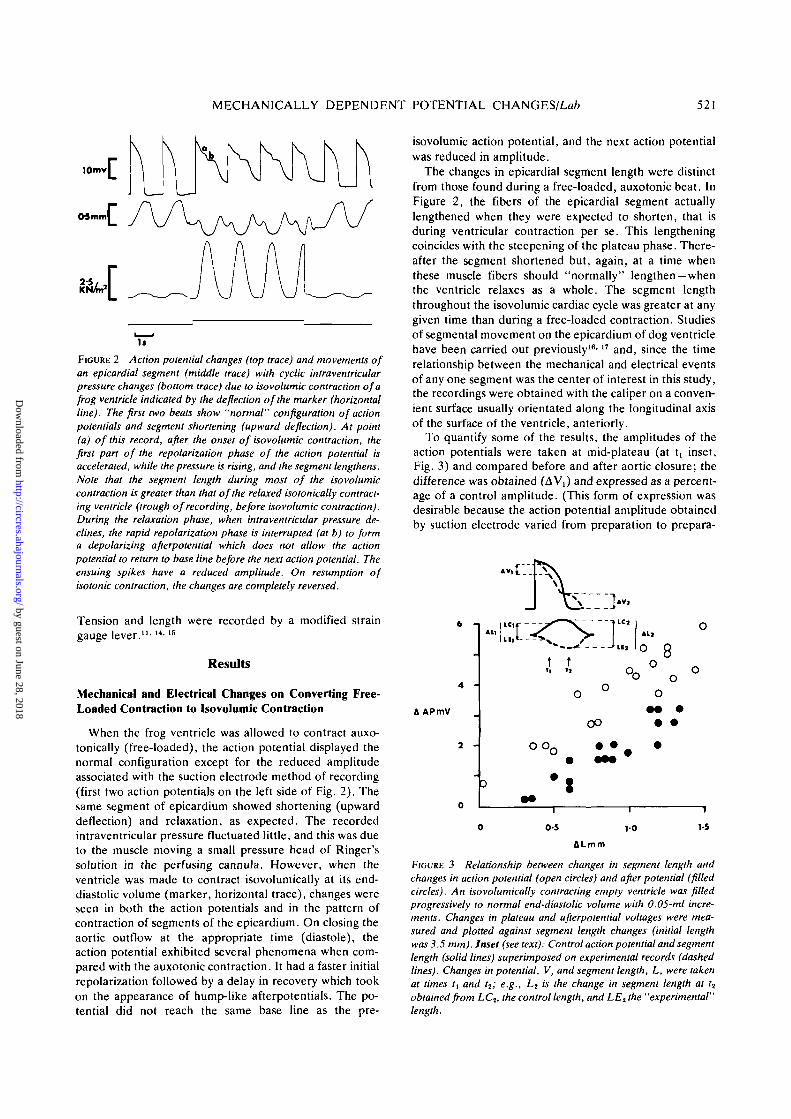
MECHANICALLY DEPENDENT POTENTIAL CHANGES/Lafc 521
10mv
05 mm
Is
FIGURE 2 Action potential changes (top trace) and movements ofan epicardial segment (middle trace) with cyclic intraventricularpressure changes (bottom trace) due to isovolumic contraction of afrog ventricle indicated by the deflection of the marker (horizontalline). The first two beats show "normal" configuration of actionpotentials and segment shortening (upward deflection). At point(a) of this record, after the onset of isovolumic contraction, thefirst part of the repolarization phase of the action potential isaccelerated, while the pressure is rising, and the segment lengthens.Note that the segment length during most of the isovolumiccontraction is greater than that of the relaxed isotonically contract-ing ventricle (trough of recording, before isovolumic contraction).During the relaxation phase, when intraventricular pressure de-clines, the rapid repolarization phase is interrupted (at b) to forma depolarizing afterpotential which does not allow the actionpotential to return to base line before the next action potential. Theensuing spikes have a reduced amplitude. On resumption ofisotonic contraction, the changes are completely reversed.
Tension and length were recorded by a modified straingauge lever."'14-15
Results
Mechanical and Electrical Changes on Converting Free-Loaded Contraction to Isovolumic Contraction
When the frog ventricle was allowed to contract auxo-tonically (free-loaded), the action potential displayed thenormal configuration except for the reduced amplitudeassociated with the suction electrode method of recording(first two action potentials on the left side of Fig. 2). Thesame segment of epicardium showed shortening (upwarddeflection) and relaxation, as expected. The recordedintraventricular pressure fluctuated little, and this was dueto the muscle moving a small pressure head of Ringer'ssolution in the perfusing cannula. However, when theventricle was made to contract isovolumically at its end-diastolic volume (marker, horizontal trace), changes wereseen in both the action potentials and in the pattern ofcontraction of segments of the epicardium. On closing theaortic outflow at the appropriate time (diastole), theaction potential exhibited several phenomena when com-pared with the auxotonic contraction. It had a faster initialrepolarization followed by a delay in recovery which tookon the appearance of hump-like afterpotentials. The po-tential did not reach the same base line as the pre-
isovolumic action potential, and the next action potentialwas reduced in amplitude.
The changes in epicardial segment length were distinctfrom those found during a free-loaded, auxotonic beat. InFigure 2, the fibers of the epicardial segment actuallylengthened when they were expected to shorten, that isduring ventricular contraction per se. This lengtheningcoincides with the steepening of the plateau phase. There-after the segment shortened but, again, at a time whenthese muscle fibers should "normally" lengthen —whenthe ventricle relaxes as a whole. The segment lengththroughout the isovolumic cardiac cycle was greater at anygiven time than during a free-loaded contraction. Studiesof segmental movement on the epicardium of dog ventriclehave been carried out previously16-17 and, since the timerelationship between the mechanical and electrical eventsof any one segment was the center of interest in this study,the recordings were obtained with the caliper on a conven-ient surface usually orientated along the longitudinal axisof the surface of the ventricle, anteriorly.
To quantify some of the results, the amplitudes of theaction potentials were taken at mid-plateau (at t, inset,Fig. 3) and compared before and after aortic closure; thedifference was obtained (AV,) and expressed as a percent-age of a control amplitude. (This form of expression wasdesirable because the action potential amplitude obtainedby suction electrode varied from preparation to prepara-
4 -
A APmV
AllICi• • .
o o o
•
tu
o
•
t
r.J°
oo
A l l
oo
o
8o
9
o
o
0-5 10
ALmm
1-5
FIGURE 3 Relationship between changes in segment length andchanges in action potential (open circles) and after potential (filledcircles). An isovolumically contracting empty ventricle was filledprogressively to normal end-diastolic volume with 0.05-ml incre-ments. Changes in plateau and afterpotential voltages were mea-sured and plotted against segment length changes (initial lengthwas 3.5 mm). Inset (see text): Control action potential and segmentlength (solid lines) superimposed on experimental records (dashedlines). Changes in potential, V, and segment length, L, were takenat times (, and t2; e.g., L2 is the change in segment length at t2
obtained from LC2, the control length, and LE2the "experimental"length.
by guest on June 28, 2018http://circres.ahajournals.org/
Dow
nloaded from

522 CIRCULATION RESEARCH VOL. 42, No. 4, APRIL 1978
tion.) The control base-line was established, using action-potentials before and after aortic closure, and the mea-surements were made from this line drawn through thegreatest diastolic potentials. The peak height of the after-potential produced on closing the aorta was measured att2 (inset, Fig. 3) and the percent difference obtained (AV2)was again expressed as a percentage of control plateauheight. The time t2 corresponded in most cases with themiddle of the fall in intraventricular pressure and, wherethe peak of the after potential was not clear, t2 was put atthis mid-fall point. Changes in segment length were ex-pressed in strains AL/L at t, and t2 (where, for example,AL is the difference in length at t, between pre- andpostaortic closure (LE-LC), and L is the control segmentlength at tt (LC). Correlation coefficients (r) betweenpotentials and lengths at t, and t2 were 0.52 and 0.235,respectively, with P < 0.001, and between 0.05 and 0.01,respectively.
In one experiment, the ventricle was emptied andrefilled in 0.05-ml steps to its end-diastolic volume. Ateach volume the same measurements as above were made,but at each volume the figures obtained were related tothe control values for the empty ventricle. The results areplotted in Figure 3. Clearly, the greater the length changethe greater the change in potential. For a given lengthchange, the change in the plateau was greater than thechange in afterpotential. Not all the changes need bepresent at the same time. Figure 4 shows an unaltered
650 N/m2
Is
FIGURE 4 The same type of experiment as in Figure 2 but with noobvious accelerated repolarization of the plateau phase of theaction potential (top trace) when the ventricle is made isovolumic(marker). In this case, the predominant lengthening of the epicar-dial segment (middle trace) takes place during the decline ofintraventricular pressure (bottom trace), and the length attained isgreater than the isotonic diastolic length —left side of picture. Atroughly the same time as the segment lengthens, during theisovolumic contraction, the rapid repolarization phase of the actionpotential is interrupted by a depolarizing afterpotential.
lOmv
2-SKN/m
Is
FIGURE 5 Action potentials (top trace) and intraventricular pres-sures (middle trace) of frog ventricle during isovolumic contraction(upward deflection of the marker on the bottom trace). In part A,isovolumic contraction resulted in an increased intraventricularpressure, and the accelerated repolarization (a) followed by depo-larizing afterpotential (b). In part B of this figure (the samepreparation), the afterpotential is accompanied by new propagatedaction potentials (c).
plateau phase with the beginnings of a normal (shortening)contraction pattern of the segment of epicardial muscle.Thereafter the segment shows an exaggerated lengtheningcompared with the same period during isotonic contrac-tion. Associated with the latter, the hump-like after-po-tential and the rise in base line are observed. Althoughaction potential changes occasionally were seen withoutsegment length changes, only once was a significant lengthchange observed with little accompanying action potentialchange. These last two observations could be explained ifepicardial contraction were inhomogeneous, with the elec-trode on a segment having a mechanical behavior differentfrom that of the "caliper" segment. Fisher et al.,IB in fact,found no distinct regular pattern of epicardial lengthchanges in their experiments. It is likely that epicardialsegments can lengthen in a direction not monitored by thecaliper, no length change being apparent, but with theaction potential still changing. The reverse is less likelybut possible. An electrical recording which is obtainedfrom a relatively unstrained bit of myocardium could thenexplain the isolated observation of a segment lengthchange with no change in potential.
Electrical Effects Related to the Recorded Changes inPotential
Threshold Depolarization
If the depolarizing afterpotential observed during iso-volumic contraction is nonartifactual, it may be expectedto be capable of reaching threshold to initiate a propa-gated action potential. Figure 5A again shows the steep-ening of the plateau phase with whole ventricle contrac-tion and the depolarizing afterpotential with whole ventri-cle relaxation (as designated by the decline in intraven-tricular pressure). In Figure 5B, this depolarizing potentialreaches threshold and produces an action potential capa-ble of initiating a contraction.
Although the afterpotentials or the points at whichthreshold are reached in Figure 5B are somewhat largerthan in Figure 5A, the last potential, d, was the largest ofall. Yet no action potential has propagated. The explana-tion for the last observation, made in several preparations,
by guest on June 28, 2018http://circres.ahajournals.org/
Dow
nloaded from

MECHANICALLY DEPENDENT POTENTIAL CHANGES/Lafr 523
is speculative at present. It is possible, first, that ionicfluxes accompanying mechanically induced multiple extra-systoles alter the immediate extracellular environmentand thus produce conditions which change the threshold.Second, the cells that actually reached threshold may havebeen distant from the electrode and many have shown, atthe time of recording potential d, a potential of reducedamplitude.
Microelectrode Recordings
The action potentials obtained by suction electrodes areproportional to intracellular potentials but are reliable infollowing slow changes in potential.10 However, duringthe changes in length of the epicardial segments there isinevitably distortion at the tissue-electrode interface. Thusthe current flowing into the electrode may vary either bya reduction in the number of cells contributing to thecurrent or by a change in the extracellular shuntingresistance. Some confirmatory studies were thereforethought necessary and were done on isolated strips ofmyocardium using microelectrodes.
It was technically difficult to keep an electrode impaledin a single cell and record reliably while imposing rapidmechanical changes on the muscle. (A success rate of 1 in60 attempts has been quoted in cat papillary muscle withlarger cells".) However, the measurement of the durationof action potentials was attempted in these experiments,for even if the full resting potential was not recorded, theduration of the action potential at 80% repolarization wasidentical with that of an action potential obtained with agood impalement. Action potential durations could thusby compared reliably at two different lengths: at slacklengths and at lengths at which near maximum tension wasproduced (Lmax)- Using several different impalements,making 16 observations in two preparations, action poten-tial duration at a slack length was 864 ± 106 (mean ± SD)msec and, at near Lmax, 790 ± 140 msec (P < 0.001 byStudent's Mest).
A good example of a change in action potential durationassociated with extension of a muscle from a length atwhich little active tension is produced to a length less thanLn,ax is seen in Figure 6. The two action potentials areconsecutive and superimposed, being obtained from thesame impalement, and the action potential duration isclearly reduced with stretch.
Although changes in action potential plateau, withalterations in length, were clearly demonstrable, consist-ent changes in resting potential or the production ofafterpotentials following mechanical changes were lessobvious. It was extremely difficult to stretch a muscle atrest and keep the electrode impaled so that, when releas-ing the muscle, the control, full resting potential wasreestablished. With resting potentials more negative thanabout -40 mV, a depolarization was the predominantresponse with a transient stretch (Fig. 7). This responsecould be explained by the electrode coming out andreimpaling the cell. However, even with short durationstretches confined to less than Lmax, a n e w propagatedaction potential that produced an active tension could begenerated (Fig. 7), which is in keeping with a depolariza-
IOO mV
0-45 G
0-5 sFIGURE 6 Superimposed traces of action potentials, obtainedwith microelectrodes, and tensions (two middle traces) duringstretch. Same impalement in an isolated strip of frog ventricle. Themuscle first contracted (I) at a short length, producing a smalltension, and was associated with the action potential with the longduration. On stretching before the second contraction (2), themuscle passive and active tension increased and the action potentialduration simultaneously shortened. Stimulation frequency, 30/min; temperature, 18°C; muscle slack length, 2.0 mm. Theserecords were obtained in Dr. B.R. Jewell's laboratory at UniversityCollege London, with the assistance of Dr. M. Boyett. Inset:Monophasic action potentials, obtained with an insulated gaptechnique (upper traces, calibration bar, 20 mV), tension (middletraces, calibration bar, 4 mN/mm2), and length (lower traces,calibration bar, downward deflection, Loto Lmax) recorded froman isolated strip of frog ventricle. In the first contraction (I) at theshort length the muscle produced almost no active tension and wasassociated with the longer action potential. During the secondcontraction (2), stretching the muscle to Lmax resulted in a largertension and the shorter action potential. Stimulation frequency 30/min; temperature, 20°C; muscle slack length, 5 mm.
tion being real and reaching threshold to activate contrac-tion. A stretch induced "extrasystole" was seen in everyisolated strip in which it was sought (n > 40).
Insulated Gap Recordings
In view of the limitations in interpreting the recordswith suction electrode and some of the microelectroderecordings, particularly the depolarization on stretch, athird method of recording was used.13 In this method, oneend of the muscle was firmly held in an insulation gapwhile the mechanical changes were obtained via the otherend. The electrical recordings were transgap potentials.Changes in action potential duration, recorded across theinsulated gap (inset, Fig. 6) were again confirmed withlength changes of the same magnitude as used with themicroelectrode studies. With this technique of recording,several transient stretches could be given at different timesduring consecutive action potentials. Brief repolarizationswere seen during the action potential, and transient de-polarizations were again produced with stretch which also
by guest on June 28, 2018http://circres.ahajournals.org/
Dow
nloaded from
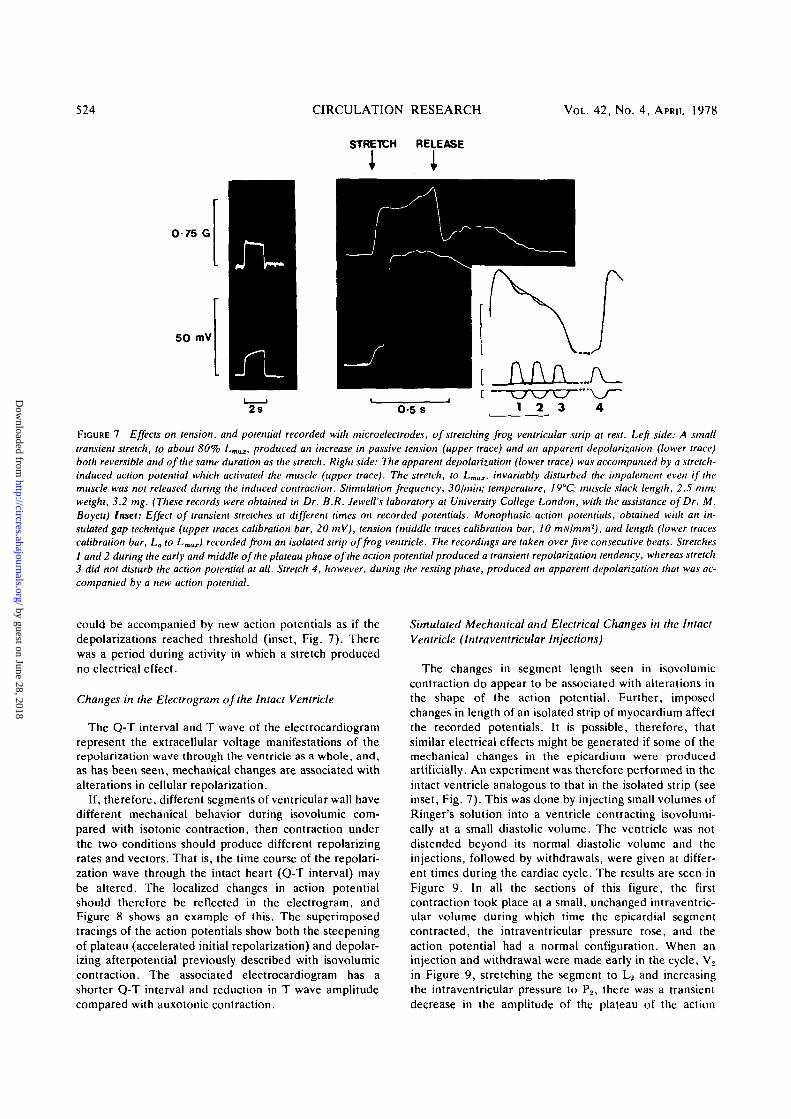
524 CIRCULATION RESEARCH VOL. 42, No. 4, APRIL 1978
STRETCH RELEASE
1 I
075 G
50 mV
2 s
FIGURE 7 Effects on tension, and potential recorded with microelectrodes, of stretching frog ventricular strip at rest. Left side: A smalltransient stretch, to about 80% Lmax, produced an increase in passive tension (upper trace) and an apparent depolarization (lower trace)both reversible and of the same duration as the stretch. Right side: The apparent depolarization (lower trace) was accompanied by a stretch-induced action potential which activated the muscle (upper trace). The stretch, to LmaI, invariably disturbed the impalement even if themuscle was not released during the induced contraction. Stimulation frequency, 30/min; temperature, I9°C; muscle slack length, 2.5 mm;weight, 3.2 mg. (These records were obtained in Dr. B.R. Jewell's laboratory at University College London, with the assistance of Dr. M.Boyett) Inset: Effect of transient stretches at different times on recorded potentials. Monophasic action potentials, obtained with an in-sulated gap technique (upper traces calibration bar, 20 mV), tension (middle traces calibration bar, 10 mNlmm2), and length (lower tracescalibration bar, Lo to Lmax) recorded from an isolated strip of frog ventricle. The recordings are taken over five consecutive beats. Stretches1 and 2 during the early and middle of the plateau phase of the action potential produced a transient repolarization tendency, whereas stretch3 did not disturb the action potential at all. Stretch 4, however, during the resting phase, produced an apparent depolarization that was ac-companied by a new action potential.
could be accompanied by new action potentials as if thedepolarizations reached threshold (inset, Fig. 7). Therewas a period during activity in which a stretch producedno electrical effect.
Changes in the Electrogram of the Intact Ventricle
The Q-T interval and T wave of the electrocardiogramrepresent the extracellular voltage manifestations of therepolarization wave through the ventricle as a whole, and,as has been seen, mechanical changes are associated withalterations in cellular repolarization.
If, therefore, different segments of ventricular wall havedifferent mechanical behavior during isovolumic com-pared with isotonic contraction, then contraction underthe two conditions should produce different repolarizingrates and vectors. That is, the time course of the repolari-zation wave through the intact heart (Q-T interval) maybe altered. The localized changes in action potentialshould therefore be reflected in the electrogram, andFigure 8 shows an example of this. The superimposedtracings of the action potentials show both the steepeningof plateau (accelerated initial repolarization) and depolar-izing afterpotential previously described with isovolumiccontraction. The associated electrocardiogram has ashorter Q-T interval and reduction in T wave amplitudecompared with auxotonic contraction.
Simulated Mechanical and Electrical Changes in the IntactVentricle (Intraventricular Injections)
The changes in segment length seen in isovolumiccontraction do appear to be associated with alterations inthe shape of the action potential. Further, imposedchanges in length of an isolated strip of myocardium affectthe recorded potentials. It is possible, therefore, thatsimilar electrical effects might be generated if some of themechanical changes in the epicardium were producedartificially. An experiment was therefore performed in theintact ventricle analogous to that in the isolated strip (seeinset, Fig. 7). This was done by injecting small volumes ofRinger's solution into a ventricle contracting isovolumi-cally at a small diastolic volume. The ventricle was notdistended beyond its normal diastolic volume and theinjections, followed by withdrawals, were given at differ-ent times during the cardiac cycle. The results are seen inFigure 9. In all the sections of this figure, the firstcontraction took place at a small, unchanged intraventric-ular volume during which time the epicardial segmentcontracted, the intraventricular pressure rose, and theaction potential had a normal configuration. When aninjection and withdrawal were made early in the cycle, V2
in Figure 9, stretching the segment to L2 and increasingthe intraventricular pressure to P2, there was a transientdecrease in the amplitude of the plateau of the action
by guest on June 28, 2018http://circres.ahajournals.org/
Dow
nloaded from
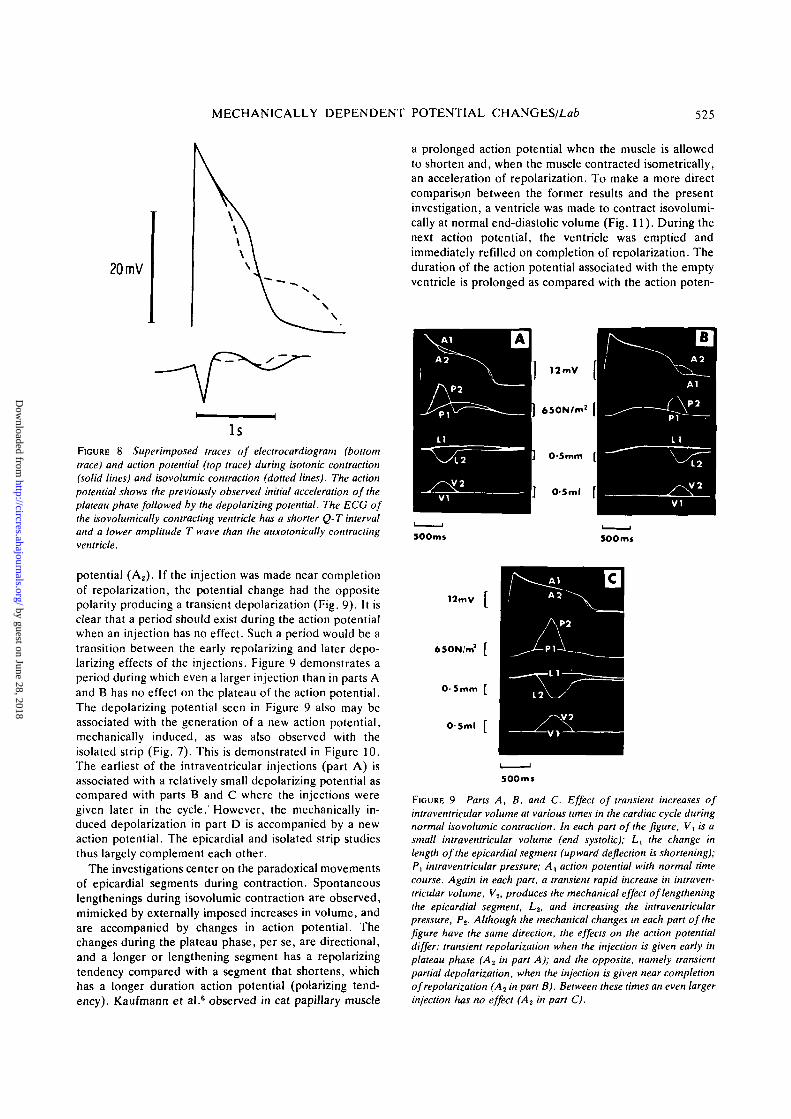
MECHANICALLY DEPENDENT POTENTIAL CHANGES/Lafc 525
20 mV
FIGURE 8 Superimposed traces of electrocardiogram (bottomtrace) and action potential (top trace) during isotonic contraction(solid lines) and isovolumic contraction (dotted lines). The actionpotential shows the previously observed initial acceleration of theplateau phase followed by the depolarizing potential. The ECG ofthe isovolumically contracting ventricle has a shorter Q-T intervaland a lower amplitude T wave than the auxotonically contractingventricle.
potential (A2). If the injection was made near completionof repolarization, the potential change had the oppositepolarity producing a transient depolarization (Fig. 9). It isclear that a period should exist during the action potentialwhen an injection has no effect. Such a period would be atransition between the early repolarizing and later depo-larizing effects of the injections. Figure 9 demonstrates aperiod during which even a larger injection than in parts Aand B has no effect on the plateau of the action potential.The depolarizing potential seen in Figure 9 also may beassociated with the generation of a new action potential,mechanically induced, as was also observed with theisolated strip (Fig. 7). This is demonstrated in Figure 10.The earliest of the intraventricular injections (part A) isassociated with a relatively small depolarizing potential ascompared with parts B and C where the injections weregiven later in the cycle. However, the mechanically in-duced depolarization in part D is accompanied by a newaction potential. The epicardial and isolated strip studiesthus largely complement each other.
The investigations center on the paradoxical movementsof epicardial segments during contraction. Spontaneouslengthenings during isovolumic contraction are observed,mimicked by externally imposed increases in volume, andare accompanied by changes in action potential. Thechanges during the plateau phase, per se, are directional,and a longer or lengthening segment has a repolarizingtendency compared with a segment that shortens, whichhas a longer duration action potential (polarizing tend-ency). Kaufmann et al.6 observed in cat papillary muscle
a prolonged action potential when the muscle is allowedto shorten and, when the muscle contracted isometrically,an acceleration of repolarization. To make a more directcomparison between the former results and the presentinvestigation, a ventricle was made to contract isovolumi-cally at normal end-diastolic volume (Fig. 11). During thenext action potential, the ventricle was emptied andimmediately refilled on completion of repolarization. Theduration of the action potential associated with the emptyventricle is prolonged as compared with the action poten-
12mV
650N/m2
O-Stnm
OSml
300ms 500 ms
12mV [
SOOms
FIGURE 9 Parts A, B, and C. Effect of transient increases ofintraventricular volume at various times in the cardiac cycle duringnormal isovolumic contraction. In each part of the figure, Vt is asmall intraventricular volume (end systolic); Lx the change inlength of the epicardial segment (upward deflection is shortening);P, intraventricular pressure; A, action potential with normal timecourse. Again in each part, a transient rapid increase in intraven-tricular volume, V2, produces the mechanical effect of lengtheningthe epicardial segment, L2, and increasing the intraventricularpressure, Pt. Although the mechanical changes in each part of thefigure have the same direction, the effects on the action potentialdiffer: transient repolarization when the injection is given early inplateau phase (A2 in part A); and the opposite, namely transientpartial depolarization, when the injection is given near completionof repolarization (A2 in part B). Between these times an even largerinjection has no effect (A? in part C).
by guest on June 28, 2018http://circres.ahajournals.org/
Dow
nloaded from
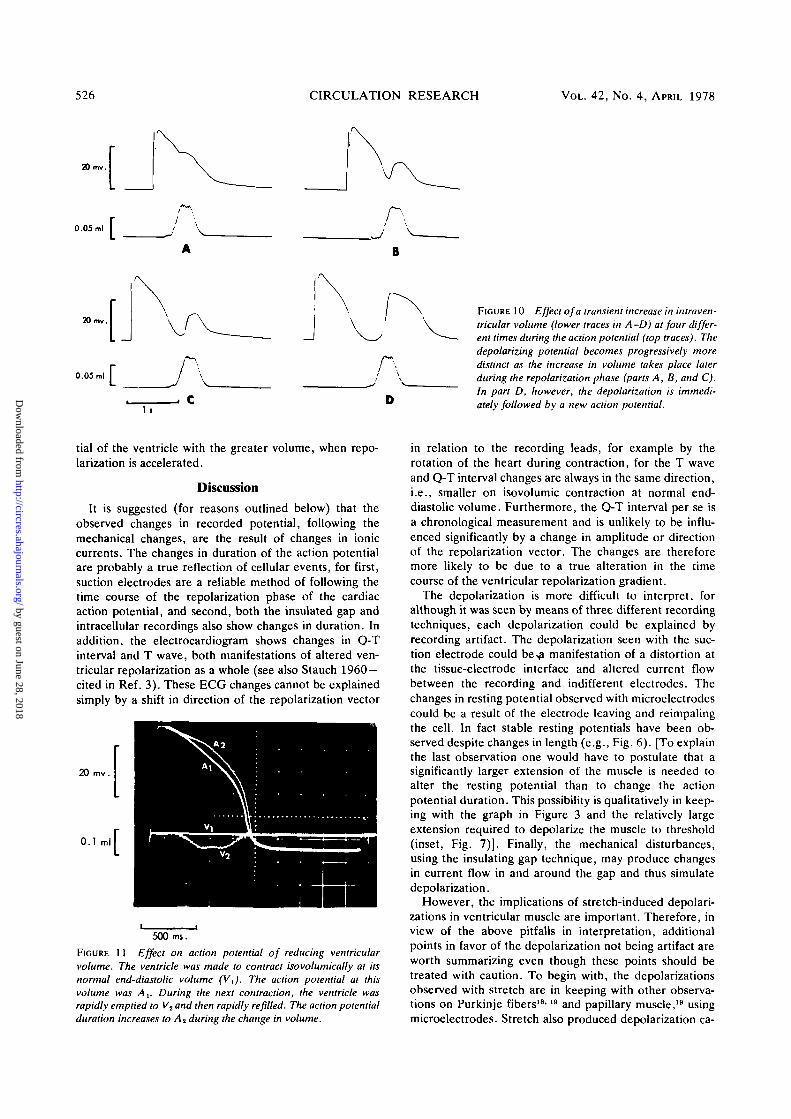
526 CIRCULATION RESEARCH VOL. 42, No. 4, APRIL 1978
20 mv.
0.05 ml
20 mv.
0.05 ml I
A
B
JA
1 s
AD
FIGURE 10 Effect of a transient increase in intraven-tricular volume (lower traces in A -D) at four differ-ent times during the action potential (top traces). Thedepolarizing potential becomes progressively moredistinct as the increase in volume takes place laterduring the repolarization phase (parts A, B, and C).In part D, however, the depolarization is immedi-ately followed by a new action potential.
tial of the ventricle with the greater volume, when repo-larization is accelerated.
Discussion
It is suggested (for reasons outlined below) that theobserved changes in recorded potential, following themechanical changes, are the result of changes in ioniccurrents. The changes in duration of the action potentialare probably a true reflection of cellular events, for first,suction electrodes are a reliable method of following thetime course of the repolarization phase of the cardiacaction potential, and second, both the insulated gap andintracellular recordings also show changes in duration. Inaddition, the electrocardiogram shows changes in Q-Tinterval and T wave, both manifestations of altered ven-tricular repolarization as a whole (see also Stauch 1960 —cited in Ref. 3). These ECG changes cannot be explainedsimply by a shift in direction of the repolarization vector
20
0.1 ml
500 ms.
FIGURE 11 Effect on action potential of reducing ventricularvolume. The ventricle was made to contract isovolumically at itsnormal end-diastolic volume (V,). The action potential at thisvolume was A,. During the next contraction, the ventricle wasrapidly emptied to V2 and then rapidly refilled. The action potentialduration increases to A2 during the change in volume.
in relation to the recording leads, for example by therotation of the heart during contraction, for the T waveand Q-T interval changes are always in the same direction,i.e., smaller on isovolumic contraction at normal end-diastolic volume. Furthermore, the Q-T interval per se isa chronological measurement and is unlikely to be influ-enced significantly by a change in amplitude or directionof the repolarization vector. The changes are thereforemore likely to be due to a true alteration in the timecourse of the ventricular repolarization gradient.
The depolarization is more difficult to interpret, foralthough it was seen by means of three different recordingtechniques, each depolarization could be explained byrecording artifact. The depolarization seen with the suc-tion electrode could be>a manifestation of a distortion atthe tissue-electrode interface and altered current flowbetween the recording and indifferent electrodes. Thechanges in resting potential observed with microelectrodescould be a result of the electrode leaving and reimpalingthe cell. In fact stable resting potentials have been ob-served despite changes in length (e.g., Fig. 6). [To explainthe last observation one would have to postulate that asignificantly larger extension of the muscle is needed toalter the resting potential than to change the actionpotential duration. This possibility is qualitatively in keep-ing with the graph in Figure 3 and the relatively largeextension required to depolarize the muscle to threshold(inset, Fig. 7)]. Finally, the mechanical disturbances,using the insulating gap technique, may produce changesin current flow in and around the gap and thus simulatedepolarization.
However, the implications of stretch-induced depolari-zations in ventricular muscle are important. Therefore, inview of the above pitfalls in interpretation, additionalpoints in favor of the depolarization not being artifact areworth summarizing even though these points should betreated with caution. To begin with, the depolarizationsobserved with stretch are in keeping with other observa-tions on Purkinje fibers18'19 and papillary muscle,19 usingmicroelectrodes. Stretch also produced depolarization ca-
by guest on June 28, 2018http://circres.ahajournals.org/
Dow
nloaded from

MECHANICALLY DEPENDENT POTENTIAL CHANGES/Lab 527
pable of reaching threshold in ventricular muscle.19 (Thelast two publications were not concerned with actionpotential durations per se.)
Furthermore, predictions on the expected behavior ofthe myocardium, based on the results being a true reflec-tion of changes in resting membrane potential, are verifiedby two observations in these present experiments. First, itis significant that the depolarizing effect, observed byusing all three recording techniques when the muscleelongates during relaxation, can apparently reach thresh-old and be followed by an "extrasystole." Second, thisnew action potential often has a reduced amplitude, as isexpected when the membrane is partially depolarized.One cannot exclude the possibility however that "stretch-sensitive" cells of the pacemaker type may conceivably befound in frog ventricle. If one allows that the length of thesegment of muscle fibers was adjusted to be near thephysiological optimum (U,ax), then the final lengthreached when the segment was stretched, for thresholddepolarization, agrees with that found in the isolatedventricular strip.
The above observations and arguments taken togetherwith results of previous experiments strongly suggest thatthe recorded electrical changes between isovolumic andfree-loaded contractions are a true reflection of alteredcurrent flow across membranes and, considering the elec-trocardiographic changes, between groups of cells, in theintact frog ventricle. Furthermore, the epicardial move-ments under these two conditions show that the mechani-cal behavior between groups of cells also differs. This issimply because epicardial fiber lengthening in one sectionof the myocardium, when taken together with a rise inintraventricular pressure, necessitates shortening and ten-sion development in other sections of the myocardium.These experiments are not as far removed from thephysiological situation as they first appear. In fact, com-prehensive studies of epicardial strains have been carriedout by Fischer et al.16 and Dieudonne and Jean,17 whoalso found positive strains (lengthening) during the normalisovolumic phase of contraction of intact dog ventricle.The segments thereafter showed some shortening duringisovolumic relaxation.
Having demonstrated that both mechanical and electri-cal behavior in a particular segment alters at the sametime, the question arises as to whether they are causallyrelated and, if so, what mechanisms are involved. It isclear from the effects of stretches on isolated strips (Figs.6 and 7), as well as the intraventricular injections inFigures 9 and 10 and other experimental situations,6 thatthe mechanical change precedes and thus is somehowrelated to the electrical change. This observed time rela-tionship, together with the evidence described above, iscompatible with the existence of a "mechano-electriccoupling" in ventricular cardiac muscle. Some "contrac-tion-exitation-contraction" coupling has been describedby Kaufmann et al.6 Keeping cat papillary muscle at alonger (isometric) length results in an abbreviated actionpotential duration as compared with a muscle that isallowed to shorten (isotonic contraction). The direction ofelectrical change is identical to that seen in the presentstudies where isovolumic contraction is accompanied by a
shorter action potential than an auxotonic contraction.The switch from isometric to isotonic contraction in thestudies on papillary muscle resulted in a small positivemechanical staircase. This mechanical transient may beattributable in part to the same mechanism that causes thetransients when the action potential is electrically pro-longed.20 Small pressure or segment length staircases wereoccasionally observed in the intact frog when altering themechanical conditions of contraction. To determinewhether these small mechanical changes are related toionic fluxes across the membrane or to the viscoelasticproperties of cardiac muscle needs further investigation.In this context, it is worth noting that a prolongation ofthe action potential in frogs affects only the immediatelyaccompanying contraction and not the subsequent beats;-"i.e., no tension transients follow. It therefore may bedifficult to ascribe the transients that were occasionallyseen in these experiments to changes in the action poten-tial per se.
It is appropriate at this stage to begin speculation as towhat architectural alteration, cellular or molecular, couldtake place during the imposed mechanical disturbance toinfluence the membrane potential. It is significant in thiscontext that both the active state (tension development)and membrane conductance are altered by changes inmembrane potential. The calcium ion, implicated in bothactive state and membrane phenomena,12 could play arole and the time course of repolarization be influencedby a change in membrane properties or internal calciumion kinetics as a result of the mechanical changes. Thischange in active state and internal calcium would in turnaffect ionic flux across the membrane either directly orindirectly. There is, in fact, considerable evidence thatcalcium release in muscle may be mechanically dependentor length dependent21"27 and that changes in internalcalcium can alter the outward potassium current.28 Onecould speculate further that the calcium ion could berelated to the depolarization observed when stretching themuscle at rest again, either by direct or indirect action.Gordon and Ridgeway27 have found in the single barnaclemuscle fiber a length-dependent electromechanical cou-pling in which a depolarization produced by stretch iscalcium mediated. It is thus conceivable that mechanicalstress or strain in the membranes of the ventricular musclecould alter the ionic permeability of the membranes thuschanging membrane potentials.
Hypotheses alternative to the preceding one are morespeculative and will be only briefly recounted, as they arediscussed in more detail elsewhere.14 First, bearing inmind the possible involvement of the active state men-tioned above, a link between a mechanically inducedchange in the muscle and membrane phenomena could be"energetically" mediated, either by cyclic AMP, orATP.29"31 Second, the immediate extracellular spaces and/or sarcoplasmic reticulum may be distorted such that analtered diffusion or accumulation of some ion thereincould influence the membrane potential to produce poten-tials analagous to those described under certain conditionswith skeletal muscle.32"34
Further discussion concerning the mechanism of "ine-chano-electric coupling" must await more direct experi-
by guest on June 28, 2018http://circres.ahajournals.org/
Dow
nloaded from

528 CIRCULATION RESEARCH VOL. 42, No. 4, APRIL 1978
mental evidence. However, if there is sufficient evidenceat this stage to support the hypothesis that some forms of"mechano-electric coupling" or mechanically dependentpotential changes are to be found in cardiac ventricularmuscle, then it is necessary for the hypothesis to fit orexplain existing observations. It is thus important tooutline the possible role and context of this phenomenonin cardiac ventricular muscle. First, it has suggested thatthere is a localized control system, operating on a beat-to-beat basis, in which the length-tension relations of cardiacventricular muscle may influence the duration of theimmediate and ensuing action potentials and active state.6
This mechanism probably is closely related and in factworks in the appropriate direction to be in keeping withthe mechanical changes described by Jewell and Rovell.35
Second, the observation that different degrees of shorten-ing or tension development of ventricular muscle areassociated with varying rates of repolarization, as indi-cated by the duration of action potentials, may be one ofthe determinants of the ventricular repolarization gra-dient. This repolarization vector (T wave) is normally inthe same direction as the QRS vector and thus deflects inthe same directions in the electrocardiogram. The T wavedirection indicates, in part, that the epicardial fibersrepolarize faster than the endocardial fibers, and it hasbeen suggested36 that this observation is related to the factthat the epicardial fibers shorten less than the endocardialfibers.37 This point cannot be more than a suggestion atthis stage, for no systematic study has been done on thetime course of these action potential changes followingmechanical changes, over a prolonged period of time.
Finally, mechano-electric coupling may have importantclinical relevance. It is now well established that there canbe exaggerated inhomogeneity of ventricular wall contrac-tion after a myocardial infarct.38-39 It is feasible that theventricular dysynergy which can follow such an episoderesults in an appropriately timed deforming stretch of thesurrounding viable myocardial tissue to produce ectopicbeats or extrasystoles and thus facilitate ventricular ar-rhythmias.'10
References1. Langer GA: Heart: excitation-contraction coupling. Ann Rev Physiol
35:55-86, 19732. Morad M, Goldman YE: Excitation-contraction coupling in heart
muscle membrane control of development of tension. Prog BiophysMolBiol 27: 259-316, 1973
3. Stauch M: Electromechanische Beziehungen am isolierten Frosch-Herzen (Das monophasische Aktionspotential bein isotonischer undisometrischer Kontraktion). Krieslaufforsch 49: 1-14, 1966
4. Bozler E: Tonus changes in cardiac muscle and their significance forthe initiation of impulses. Am J Physiol 139: 477-480, 1943
5. Lab MJ: The effect on the left ventricular action potential of clampingthe aorta. J Physiol (Lond) 202: 73-74P, 1969
6. Kaufmann RL, Lab MJ, Hennekes R, Krause H: Feedback interactionof mechanical and electrical events in the isolated mammalian ventric-ular myocardium (cat papillary muscle). Pfleugers Arch 324: 100-123, 1971
7. Speare JF, Moore EN: A comparison of alternation in myocardialaction potentials and contractility. Am J Physiol 220: 1708-1716,1971
8. Bozler E, Delahayes: Mechanical and electrical oscillations in cardiacmuscle of the turtle. J Gen Physiol 62: 523-534, 1971
9. Hoffman BF, Cranefield PF, Lepeschkin E, Surawicz B, Herrlich HC:Comparison of cardiac monophasic action potentials recorded byintracellular and suction electrodes. Am J Physiol 196: 1297-1301,1959
10. Lab MJ, Morris GA: A caliper for sampling relative movement oftissue surfaces. J Physiol (Lond) 224: 55-56P, 1972
11. Lewis AF: An improved isotonic transducer. J Physiol (Lond) 203:15-17P, 1969
12. Allen DG, Jewell BR, Wood EH: Studies of the contractility ofmammalian myocardium at low rates of stimulation. J Physiol (Lond)254:1-17, 1976
13. Niedergerke R: The staircase phenomenon and the action of calciumon the heart. J Physiol (Lond) 134: 569-583, 1956
14. Lab MJ: Mechano-electric interactions in cardiac muscle. Ph.D.Thesis, University of London, 1974
15. Lewis AF: An isometric strain gauge transducer unit (abstr). J Physiol(Lond)203: 17P,1969
16. Fisher VJ, Lee RJ, Gourin A, Bolocki H, Stuckey JH, Kavaler F:Muscle fiber length: A determinant of left ventricular contractionpattern. Am J Physiol 211: 301-306, 1966
17. Dieudonne JM, Jean CF: Epicardial strains during left ventricularcontraction cycle. Am J Physiol 216: 1188-1193, 1969
18. Deck KA: Alterations in the resting potential and the cable propertiesof Purkinje fibres during stretch. Pfleugers Arch 280: 131-140, 1964
19. Kaufmann R, Theophile U: Automatic-fordernded Dehnungseffecteam Purkinje-Faden, Papillarmuskeln und Vorhoftabekein van Rhesus-Affen. Pfleugers Arch 291: 174-189, 1967
20. Antoni H, Jacob R, Kaufmann R: Merchanische reaction des Frosch-und-Saugtiermyokard bei veranderungen der Aktions-potentialdaurdurch konstante gleichstromimpulse. Pfleugers Arch 306: 33-57.1969
21. Allen DG, Jewell BR, Murray JW: The contribution of activationprocess to the length-tension relation of cardiac muscle. Nature 248:606-607, 1974
22. Lakatta, EG, and Jewell BR: Length-dependent activation. Its effecton the length-tension relation in cat ventricular muscle. Circ Res 40:251-257,1977
23. Fabiato A, Fabiato F: Dependence of the contractile activation ofskinned cardiac cells on the sarcomere length. Nature 256: 54-56,1975
24. Taylor SR: Vetebrate striated muscle: Length dependence of calciumrelease during contraction. Eur J Cardiol 4: 31-40, 1976
25. Winegrad S, McClellan G, Robinson T, Nan Ping L: Variable diastoliccompliance and variable Ca sensitivity of the contractile system incardiac muscle. Eur J Cardiol 4: 41-46, 1976
26. Henderson AH, Cattel MR: Length as a factor in excitation-contrac-tion coupling in heart muscle: An investigation of mechanical tran-sients in strontium mediated contractions. Eur J Cardiol 4: 47-52,1976
27. Gordon AM, Ridgeway EB: Length-dependent electromechanicalcoupling in single muscle fibres. J Gen Physiol 68: 653-670, 1976
28. Bassingthwaighte JB, Fry CH, McGuigan JAS: Relationship betweeninternal calcium and outward current in mammalian ventricular mus-cle; a mechanism for the control of the action potential duration. JPhysiol (Lond) 262: 15-38, 1976
29. Sutherland EW: On the biological role of cyclic AMP. J Am MedAssoc214: 1281-1288, 1970
30. McDonald RF, MacLoed DP: Anoxia-recovery cycle in ventricularmuscle: Action potential duration, contractility and ATP content.Pfluegers Arch 325: 305-322, 1971
31. McDonald RF, MacLeod DP: The effect of 2,4-dinitrophenol onelectrical and mechanical activity, metabolism and ion movements inguinea-pig ventricular muscle. Br J Pharmacol 44: 1-12, 1972
32. Freygang WH, Rapoport SI, Peachy LD: Some relations between thelinear electrical properties of striated muscle fibres and changes inultrastructure. J Gen Physiol 50: 2437-2458, 1967
33. Rapoport S: Swelling of the transverse tubular system in frog sartorius.J Gen Physiol 54: 166-177, 1969
34. Gage PW, Eisenberg RS: Action potentials without contraction infrog skeletal muscle fibres with disrupted transverse tubules. Science158:1702-1703, 1967
35. Jewell BR, Rovell JM: Influence of previous mechanical events on thecontractility of isolated cat papillary muscle. J Physiol (Lond) 235:715-740, 1973
36. Lab MJ: Comparison action potentials from endocardial and epicar-dial surfaces of frog ventricle. J Physiol (Lond) 216: 44-45P. 1971
37. Rushmer RF: Cardiovascular Dynamics, ed 3. Philadelphia, W. B.Saunders, 1970.
38. Tennant R, Wiggers DJ: The effect of coronary occlusion on myocar-dial contraction. Am J Physiol 112: 351-361, 1935
39. Tyberg JV, Forester JS, Parmley W: Altered segmental function andcompliance in acute myocardial ischemia. Eur J Cardiol 1: 307-317.1974
40. Scherf D, Schott A: Extrasystoles and Allied Arrhythmias. NewYork, Heineman, 1973
by guest on June 28, 2018http://circres.ahajournals.org/
Dow
nloaded from

M J LabMechanically dependent changes in action potentials recorded from the intact frog ventricle.
Print ISSN: 0009-7330. Online ISSN: 1524-4571 Copyright © 1978 American Heart Association, Inc. All rights reserved.is published by the American Heart Association, 7272 Greenville Avenue, Dallas, TX 75231Circulation Research
doi: 10.1161/01.RES.42.4.5191978;42:519-528Circ Res.
http://circres.ahajournals.org/content/42/4/519World Wide Web at:
The online version of this article, along with updated information and services, is located on the
http://circres.ahajournals.org//subscriptions/
is online at: Circulation Research Information about subscribing to Subscriptions:
http://www.lww.com/reprints Information about reprints can be found online at: Reprints:
document. Permissions and Rights Question and Answer about this process is available in the
located, click Request Permissions in the middle column of the Web page under Services. Further informationEditorial Office. Once the online version of the published article for which permission is being requested is
can be obtained via RightsLink, a service of the Copyright Clearance Center, not theCirculation Research Requests for permissions to reproduce figures, tables, or portions of articles originally published inPermissions:
by guest on June 28, 2018http://circres.ahajournals.org/
Dow
nloaded from


















