Chapter 30 Disorders of Hepatobiliary and Exocrine Pancreas Function

4-HYDROXYAMINOQUINOLINE-1-OXIDE-INDUCED ULTRA- STRUCTURAL CHANGES IN THE GUINEA-PIG EXOCRINE PANCREAS
M. S. RAO AND J. K. REDDY* Department of Pathology and Oncology, University of Kansas Medical Center,
College of Health Sciences and Hospital, Kansas City, Kansas 66103
PLATES XXXVIII-XLIII
~ - H Y D R ~ X Y A M I N O Q ~ I N O L I N E - ~ - O X I D E (4-HAQO), a catalytic reduction product of 4-nitroquinoline-1-oxide (Ochiai, Ohta and Nomura, 1957) possesses mutagenic (Okabayashi, 1962) and carcinogenic (Shirasu and Ohta, 1963) properties. This compound has been used in various experimental animals for the induction of cancer at different sites such as stomach (Mori et a/., 1969a and b), lung (Mori et al., 1961), and subcutaneous tissue (Endo and Kume, 1963). Recently Hayashi and Hasewaga (1971) have shown that 4-HAQO induced a high incidence of adenoma or adenoma-like lesions in the exocrine pancreas of rats. Although, the compound 4-HAQO has been studied mainly in terms of its carcinogenicity, its acute effects on various organs including pancreas have received very little attention (Konishi, Popp and Shinozuka, 1974; Reddy et a/., 1975). To our knowledge the subcellular effects of 4-HAQO on pancreatic acinar cells have not been reported. The present investigation, therefore, deals with the acute ultrastructural changes in the exocrine pancreas of guinea-pigs given a single intravenous injection of 4-HAQO.
MATERIALS AND METHODS
Both male and female inbred guinea-pigs (NIH strain 13, obtained from the Frederick Cancer Research Center, Frederick, Md.) weighing 250-300 gm were used in these studies. The animals were fed standard diet (Purina guinea-pig chow) ad libitum and had free access to water. 4-hydroxyaminoquinoline-I-oxide (4-HAQO) was dissolved in 0.005N-HCl and injected intravenously into the antecubital vein at a dose of 22.5 mg per kilogram body weight. Control animals were given a corresponding volume of 0.005N-HCI by the same route. Four animals (two male and two female) were sacrificed under light ether anesthesia, at 18, 24, 43, 50, 55 and 60 hr after the injection of 4-HAQO. Portions of the pancreas were prepared for light microscopy after fixing in 10 per cent. neutral buffered formalin, embedded in paraffin wax and sections were stained with haematoxylin and eosin. For electron microscopy, random samples of pancreas were fixed in 2 per cent. osmium tetroxide buffered to pH 7.4 with S-collidine. After fixation for 2 hr at 0"-5"C, tissues were dehydrated in graded series of alcohol and embedded in epon 812 (Luft, 1961). One micron thick sections were cut with glass knives, stained with Toluidine blue, and were examined with a Zeiss Ultraphot I1 microscope. Ultrathin sections were cut with an LKB ultramicrotome using glass knives. Sections were stained with lead hydroxide and examined in an RCA EMU-3G or Phillips 300 electron microscope.
Received 3 Nov. 1975; accepted 1 Dec. 1975. *Present address: Department of Pathology, Northwestern University School of Medicine,
Chicago, Illinois 6061 1. J. PATH.-VOL. 120 (1976) 109

110 M. S. RAO AND J. K. REDDY
RESULTS The morphology of the acinar cells of the exocrine pancreas in control
guinea-pigs conformed to previous descriptions (Palade, 1956; Palade and Siekevitz, 1956). In guinea-pigs given 4-HAQO, the ultrastructural changes varied considerably at a given interval from acinus to acinus as well as from cell to cell within a particular acinus.
Changes at 18 and 24 hr. Between 18 and 24 hr after the single dose of 4- HAQO, the nucleoli of acinar cells appeared somewhat larger than normal and displayed segregation of the granular and fibrillar components into distinct areas (fig. 1). The interchromatin granules in the nucleoplasm appeared prominent. No changes in the nuclear membranes were evident at this interval. The cytoplasmic alterations were characterised by minimal dilatation of rough endoplasmic reticulum (RER) throughout the cell. The membrane attached and free ribosomes appeared unaltered. The intracisternal granules, which are normally seen in guinea-pig pancreatic acinar cells, were markedly decreased. The mitochondria, in general, were not markedly altered, but an occasional one was swollen with disruption, or focal dilatation of the outer membrane. The Golgi complex, in most cells, remained intact. The mature zymogen granules were markedly decreased in number, and the apical cytoplasm was occupied by increased number of immature (prozymogen) granules. There was a moderate increase in the number of cytoplasmic dense bodies in acinar cells of guinea-pigs given 4-HAQO, which were only occasionally encountered in control animals. During this period no changes were observed in centroacinar and ductal cells.
Changes at 43 and 60 hr. The acinar architecture of the pancreas appeared completely distorted in most areas. The peak degenerative changes in the exocrine pancreas were noted at 55 hr by light microscopy, although few scattered acini or several cells in a particular acinus appeared unaffected (fig. 2). By electron microscopy the RER showed vesicular to cisternal dilatation contain- ing either electron lucent, or finely fibrillar, electron-dense material (figs. 3 and 4). The focal mitochondria1 changes observed at earlier intervals showed further progression, with disruption of limiting membranes, and cristae. Some of the disrupted mitochondria were seen in autophagic vacuoles.
The zymogen granules were markedly decreased or completely absent in most cells. Even the prozymogen granules were rarely seen. In some cells the membrane bound vacuoles were increased. An occasional atypical large zymogen granule displayed striations in the matrix (fig. 5). A conspicuous reduction in the size of the Golgi complex was evident. A marked increase in the number of lysosomes was noted in the apical cytoplasm of some cells.
In addition to the above described changes in various organelles, the hyalo- plasm contained pleomorphic cytoplasmic degenerative bodies composed of membrane bound vacuoles (fig. 6), myelin figures (fig. 7), or a combination of both. Jn addition, numerous small to large autophagic vacuoles containing recognisable cytoplasmic organelles such as RER, ribosomes and mitochondria (fig. S), were also observed. At 43 lir some acinar cells displayed lipid droplets

PANCREATIC EFFECTS OF 4-HAQO 1 1 1
in the cytoplasm as well as in the nucleus. By 55 hr numerous acinar cells were necrotic.
The nucleolar segregation seen at earlier intervals was no longer persistent in the acinar cells of animals killed between 43 and 60 hr after 4-HAQO injec- tion. Instead, the nucleoli were small, appeared fragmented or condensed. Depending on the severity of cell injury, the nuclear changes consisted of margination of chromatin, or fragmentation of nuclear envelope leading to com- plete dissolution of nucleus.
Centroacinar and ductal cell changes. The centroacinar cells are normally situated with acinar cells and participate in the formation of acinar lumen. The centroacinar cells in guinea-pigs resembled those described by Ekholm, Zealander and Edlund (1962) in rats. However, in guinea-pigs the mitochondria are not scanty as in rats. The ductal cell morphology was similar to that described in rats (Ekholm et al.). Both centroacinar and ductal cells showed degenerative changes to a variable degree including the organelle changes, cytoplasmic degenerative bodies, increased lysosomes and fat vacuoles (fig. 9). Necrotic cells were seen in both centroacinar and ductal cell types but were infrequent (fig. 10).
DISCUSSION
It is evident from these ultrastructural studies that, after an intravenous injection of 4-HAQO, marked cytoplasmic and nuclear alterations in acinar, cen troacinar and ductal epithelium of exocrine pancreas of guinea-pigs are produced. These studies extend our earlier observations, by light microscopy, of pancreatic damage (Reddy et al. 1975) and deal with detailed analysis of the subcellular events leading to cell necrosis.
The earliest morphological changes observed in the development of 4-HAQO- induced exocrine pancreatic injury are nucleolar segregation and progressive dilatation of endoplasmic reticulum channels followed by detachment and depletion of ribosomes. The segregation of granular and fibrillar components of the nucleolus into distinct zones was observed in acinar cells at 18 and 24 hr after 4-HAQO administration. This type of nucleolar alteration can be easily induced in liver cells by a variety of chemical carcinogens (Svoboda and Higginson, 1968) and antimetabolites (Schoefl, 1964) and is associated with inhibition of DNA-dependent RNA polymerase activity (Reddy, et al., 1968; Reddy and Svoboda, 1968). In contrast, nucleolar segregation in pancreatic acinar cells is not a frequent alteration and has been observed only with anthra- inycin (Harris et al., 1968b), cycloheximide (Harris et al., 1968a), and actinomycin D (Rodriguez, 1967; Horky and Toman, 1971). The induction of nucleolar segregation by 4-HAQO in pancreatic acinar cells may be related to its capacity to complex with DNA (Kawazoe et nl., 1972) resulting in an interference with chromatin-nucleolar axis. Whatever might be the underlying mechanism of nucleolar segregation, this alteration is associated with inhibition of RNA synthesis (Simard and Bernhard, 1966; Reddy and Svoboda) and may eventually result in decreased protein synthesis. The changes in RER as well as progressive

112 M. S. RAO AND J. K . REDDY
depletion of mature zymogen granules are indicative of altered protein synthesis, and are consistent with the decreased pancreatic amylase activity noted in our earlier studies (Reddy et al. 1975). These changes, however, are considered rather non-specific and have been reported in the pancreas of rats and guinea pigs treated with compounds such as ethionine (Herman and Fitzgerald, 1962; Boquist, 1969), aflatoxin B1 (Rao et al., 1974), anthramycin (Harris et al. 1968b), Puromycin (Longnecker et al., 1968) and azaserine (Rao and Reddy, 1976), which inhibit either RNA or protein synthesis. The mitochondrial lesions as well as appearance of lipid droplets in these cells at later intervals, are considered as further evidence for impaired protein synthesis (Weisblum et al., 1962). However, the possibility that mitochondrial damage may be responsible for lipid accumulatioii cannot be excluded (Scott and Vermillion, 1966; Darle et al., 1970).
The occurrence of pleomorphic cytoplasmic degenerative bodies in acinar, centroacinar and ductal cells reflects the degree of cell damage. The presence of these lesions signifies a common but rather limited pattern of cellular reaction to various injurious effects (Herman and Fitzgerald; Scott and Vermillion; Longnecker et al; Boquist; Rao et al. 1974).
It is apparent from these studies, that the acute effects of 4-HAQO on pancreas, after a single intravenous injection, are mainly necrotic. Although interference with DNA directed RNA synthesis was implicated (Kawazoe, et al.) the exact mechanism of the pancreaticotoxic effect of 4-HAQO is unknown. Probably the selective toxic effects on pancreas may be due to an increased uptake of 4-HAQO by the pancreas and the inability of this organ to metabolise this compound to non-toxic metabolites.
SUMMARY 4-hydroxyaminoquinoline, a carcinogen was injected into guinea-pigs
intravenously in a single dose of 22.5 mg/kg. The sequential morphological changes in the exocrine cells of the pancreas were studied by electron micro- scopy at selected intervals between 18 and 60 hr. The earliest alterations in the acinar cells comprised nucleolar segregation into granular and fibrillar components and minimal dilatation of endoplasmic reticulum. At later intervals all the cytoplasmic organelles appeared altered. The endoplasmic reticular channels were markedly dilated with depletion of membrane bound and free ribosomes. The mitochondria were swollen and irregular with disruption of limiting membranes and cristae. The zymogen granules were markedly reduced or completely absent. The hyaloplasm contained morphologically variable cytoplasmic degenerative bodies. In addition to the acinar cells, centriacinar and ductal epithelial cells showed similar changes.
This work was supported by Public Health Service Contract CP-3271 from the division of Cancer Cause and Prevention, National Cancer Institute. We thank Saeed Ahrnad Qureshi for the excellent technical assistance and Miss Juanita Stika for help in preparing this manuscript.

RAO AND REDDY
PANCREATIC EFFECTS OF 4-HAQO
P L A r E XXXVIII
FIG. 1.-Acinar cell of the pancreas, 18 hr after administration of 4-HAQO, showing segregation of nucleolus into granular and fibrillar components. Note also increase in the number of inter- chromatin granules in the nucleoplasm. x 25,600.
FIG. 2.-Marked necrosis of acinar cells of the pancreas 55 hr after administration of 4HAQO. Haematoxylin and eosin. x 140.

RAO AND REDDY
PANCREATIC EFFtCr.5 OF 4-HAQO
PLATE XXXIX
Fici. 3.-Portion of an acinar cell of the pancreas, 50 hr after administration of 4-HAQO, showing cisternal dilatation of endoplasmic reticulum containing fine granular and fibrillar material.
~ 13.500.

PANCREATIC EFFECTS OF 4-HAqO
PLATE XL
FIG. 4.-Portion of another acinar cell, 50 hr after administration of 4-HAQO, showing irregularly dilated endoplasmic reticulum. The mitochondria are pleomorphic with disruption of limiting membranes and disorganisation of cristae. x 24,000.

I’ANCKTATIC t t t t C IS Or 4-HAQV
PLATI: XLI
FIG. 5.-Abnormal zymogen granules containing electron-dense homogenous and fibrillar material. :< 44.500.
FIG. 6.-The pancreatic acinar cell, 43 hr after administration of 4-HAQO, containing smooth membrane-bounded vacuoles. x 12,000. smooth membranes (myelin figures). x 16,800.
FG. 7.-Another pancreatic acinar cell, 50 hr after admin- istration of 4-HAQO, containing whorls of closely packed
RAO AND RFI>I )Y
PANCREATIC EFFECTS OF 4-HAQO
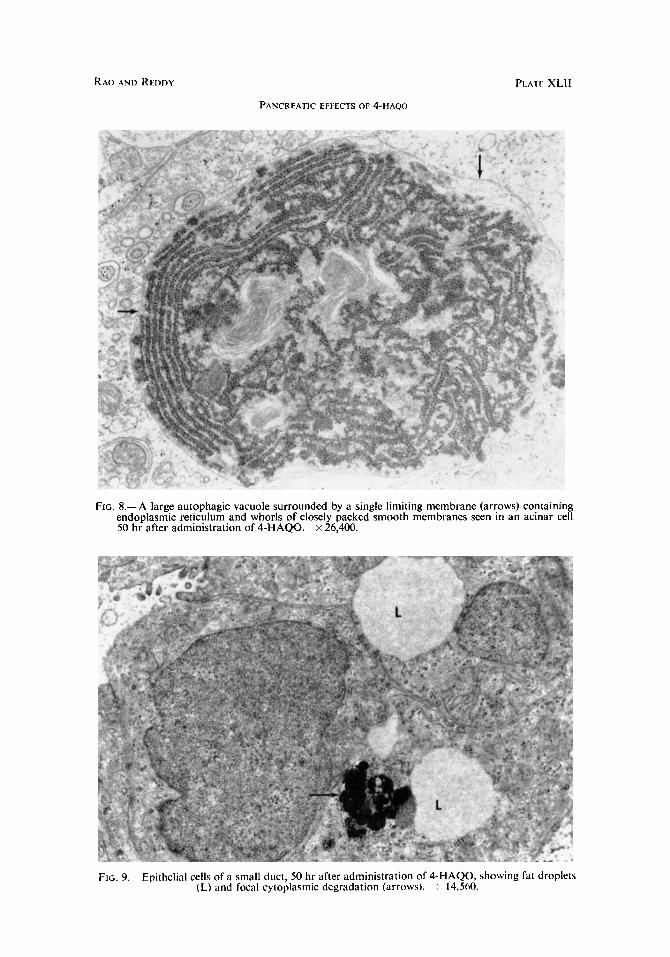
FIG. 8.-A large autophagic vacuole surrounded by a single limiting membrane (arrows) containing endoplasmic reticulum and whorls of closely packed smooth membranes seen in an acinar cell 50 hr after administration of 4-HAQO. x 26,400.
FIG. 9.- Epithelial cells of a small duct, 50 hr after administration of 4-HAQO, showing fat droplets (L) and focal cytoplasmic dcgradation (arrows). 14,560.
RAO AND REDDY
PANCREATIC EFFECTS OF 4-HAQO
PLATE XLlll
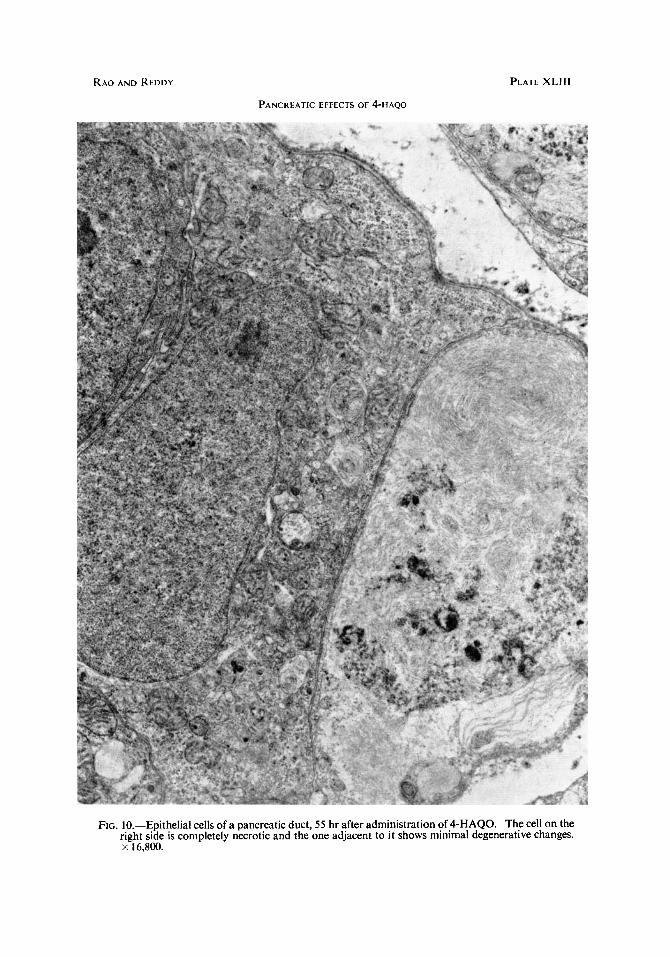
FIG. 10.-Epithelial cells of a pancreatic duct, 55 hr after administration of 4-HAQO. The cell on the right side is completely necrotic and the one adjacent to it shows minimal degenerative changes. x 16.800.

PANCREATIC EFFECTS OF 4-HAQO 113
REFERENCES BOQUIST, L. 1969. Morphologic effects of ethionine on the pancreas of the Chinese hamster.
Acta Path. Microbiol. Scand., 76, 91. DARLE, N., EKHOLM, R., AND EDLUND, Y . 1970. Ultrastructure of the rat exocrine pancreas
after long-term intake of ethanol. Gastroenterol., 58, 62. EKHOLM, R., ZEALANDER, T., AND EDLUND, Y . 1962. The ultrastructural organization of the
rat exocrine pancreas. J. Ultrastruct. Res., 7, 73. ENDO, H., AND KUME, F. 1963. Comparative studies on the biological actions of 4-nitro-
quinoline-1-oxide and its reduced compound 4-hydroxyaminoquinoline-1-oxide. Gann, 54, 443.
HARRIS, C., GRADY, H., AND SVOBODA. D. 1968a. Alterations in Dancreatic and heDatic ultrastructure following acute cycloheximide intoxication. J.-Ultrastruct. Res.; 22, 240.
HARRIS, c. , GRADY, H., AND SVOBODA, D. 1968b. Segregation of the nucleolus produced by anthramycin. Cancer Res., 28, 81.
HAYASHI, Y., FURUKAWA, H., AND HASEGAVA. T. 1972. Pancreatic tumors in rats induced by 4-nitroquinoline-1-oxide derivatives. In Topics in chemical carcinogenesis, edited by W. Nakara, S. Takayama, T. Sugimurs, and S. Odashima, University Park Press, Baltimore, pp. 53-65.
HAYASHI, Y . , AND HASEGAVA, T. 1971. Experimental pancreatic tumor in rats after intra- venous injection of 4-hydroxyaminoquinoline-1-oxide. Gann, 62, 329.
HERMAN, L., FITZGERALD, P. 1962. The degenerative changes in pancreatic acinar cells caused by DL-Ethionine. J. Cell. Biol., 12, 277.
HORKY, D., AND TOMAN, J. 1971. Effect of actinomycin D on the submicroscopic structure of the exocrine cells of the adult rat pancreas. Folia Morphologica, 19, 51.
KAWAZOE, Y., ARAKI, M., AND HUANG, G. 1972. Chemical aspects of carcinogensis by 4-nitroquinoline-1-oxide. In Topics in chemical carcinogensis. edited by W. Nakahara, S. Takayama, T. Sugimura, and S. Odashima, Baltimore, pp. 1-13.
KONISHI, Y. , POPP, J., AND SHINOZUKA, H. 1974. Pancreatic acinar cell damage induced by 4-nitroquinoline-1-oxide and 4-hydroxyaminoquinoline-1-oxide. J. Nail. Cancer Inst., 52, 917.
LONGNECKER, D. S., SHINOZUKA, H., AND FARBER, E. 1968. Electron microscopy of rat pancreatic acinar cells in puromycin induced mitosis. Amer. J. Pathol., 52, 891.
LUFT, J. H. 1961. Improvements in epoxy resin embedding methods. J. Biophys. & Biochem. Cyrol., 9, 409.
MORI, K., KONDO, M., KOIBUCHIE, E., AND HASHIMOTO, A. 1961. Induction of lung cancer in mice by injection of 4-hydroxyaminoquinoline-1-oxide. Gann, 58, 105.
MORI, K., OHTA, A., MURAKAMI, T., TAMURA, M., AND KONDO, H. 1969a. Carcinoma of the glandular stomach of mice induced by 4-hydroxyaminoquinoline-1-oxide hydro- chloride. Gunn, 50, 15 1.
MORI, K., OHTA, A., MURAKAMI, T., TAMURA, M., KONDO, M., AND ISHIMURA, H. 19696. Carcinomaof theglandular stomach and other organs of rats by4-hydroxyarninoquinoline- 1-oxide hydrochloride. Gann, 60, 627.
OCHIAI, E., OHATA, A., AND NOMURA, H. 1957. Polarisation der heterozyklischen Ringe mit aromatischem charakter. Uber das 4-hydroxyaminochinolin-N-oxyd. Chenz. Pharm. Bull (Tokyo), 5, 310.
OKABAYASHI, T. 1962. Mutagenic activity of 4-hydroxyaminoquinoline-1-oxide. Chem. Pharm. Bull (Tokyo), 10, 1127.
PALADE, G. E. 1956. Intracisternal granules in the exocrine cells of the pancreas. J. Biophys. Biochem. Cytol., 2, 417.
PALADE, G. E., AND SIEKEVITZ, P. 1956. Pancreatic microsomes. An integrated morpholo- gical and biochemical study. J. Biophys. Biochem. Cytol., 2, 671.
RAO, M. S., AND REDDY, J. K. 1976. Subcellular changes in exocrine pancreas of the guinea pig induced by azaserine. Biochem. & Exp. Biol., in press.
J. PATH.-VOL. 120 (1976) H

114 M. S. RAO AND J. K. REDDY
RAO, M. S., SVOBODA, D., AND REDDY, J. K. 1974. The ultrastructural effects of aflatoxin B1 in the rat pancreas. Virchows Arch. B. Cell. Path., 17, 149.
REDDY, J., HARRIS, C., AND SVOBODA, D. 1968. Inhibition by lasiocarpine of RNA synthesis, RNA polymerase and induction of tryptophan pyrrolase activity. Nature, 217, 659.
REDDY, J. K., AND SVOBODA, D. J. 1968. The relationship of nucleolar segregation to ribonucleic acid synthesis following the administration of selected hepatocarcinogens. Lab. Invest., 19, 132.
REDDY, J. K., RAo, M. S., SVOBODA, D. J., PRASAD, J. D. 1975. Pancreatic necrosis and regeneration induced by 4-hydroxyaminoquinoline-I-oxide in the guinea pig. Lab. Invest., 32, 98.
RODRIQUEZ, T. G. 1966. Ultrastructural changes in the exocrine pancreas induced by prolonged treatment with actinomycin D. J. Ultrastruct. Res., 19, 116.
SCHOEFL, G. 1964. The effect of actinomycin D on the fine structure of the nucleolus. J. Ultrastruct. Res., 10, 224.
SCOTT, E. B., AND VERMILLION, S. D. 1966. Histopathology of amino acid deficiencies. Arch. Path., 82, 119.
SHIRASU, Y., AND OHTA, A. 1963. A preliminary note on the carcinogenicity of 4-hydroxy- aminoquinoline-1-oxide. Gann, 54, 221.
SIMARD, R. D., AND BERNHARD, W. 1966. Le Phenomtne de la Segregation nucleolaire: SpecificitC d’action de certains antimetabolites. Intern. J . Cancer. 1, 463.
SVOBODA, D., AND HIGGINSON, J. 1968. A comparison of ultrastructural changes in rat liver due to chemical carcinogens. Cancer Res., 28, 1703.
WEISBLUM, B., HERMAN, L., AND FITZGERALD, P. 1962. Changes in pancreatic acinar cells during protein deprivation. J . Cell Biol., 12, 313.