
2013 Eagle Creek Effect of White Tail Deer Report Jenkins, Purdue University
18
Effects of herbivory by white-tailed deer on forest vegetation in Eagle’s Crest and Spring Pond Nature Preserves, Eagle Creek Park, Indianapolis, Indiana Prepared by: Michael A. Jenkins Associate Professor of Forest Ecology Department of Forestry and Natural Resources Purdue University West Lafayette, Indiana October 7, 2013
-
Upload
indy-parks -
Category
Documents
-
view
217 -
download
0
description
Â
Transcript of 2013 Eagle Creek Effect of White Tail Deer Report Jenkins, Purdue University

Effects of herbivory by white-tailed deer on forest vegetation in Eagle’s Crest
and Spring Pond Nature Preserves, Eagle Creek Park, Indianapolis, Indiana
Prepared by:
Michael A. Jenkins
Associate Professor of Forest Ecology
Department of Forestry and Natural Resources
Purdue University
West Lafayette, Indiana
October 7, 2013

2
Introduction
Chronic herbivory by overabundant white-tailed deer (Odocoileus virginianus) populations
threaten native plant communities in forests throughout eastern North America (Waller and
Alverson 1997, Rooney 2001, Russell et al. 2001, Côté et al. 2004, Webster et al. 2005, Heckel et
al. 2010). Chronic herbivory often drives the local extinctions of woody and herbaceous
browse-sensitive species (Rooney et al. 2004, Thiemann et al. 2009), facilitates the invasion and
recalcitrance of non-native invasive plants (Webster et al. 2008, Knight et al. 2009), shifts
overstory composition towards non-preferred species (Tilghman 1989, Waller and Alverson
1997, Rossell et al. 2005, Jenkins 2011, Tanentzap et al. 2011), and alters below ground
communities and processes (Bressette et al. 2012, Lessard et al. 2012).
In natural areas managed to exclude human activities, hunting is commonly prohibited in favor
of natural regulation. Under such a system, deer populations are allowed to fluctuate naturally
and typically become overabundant without hunting or predation; a response that is
augmented in fragmented landscapes with abundant edge habitat and/or mild winters (Leopold
et al. 1947, Alverson et al. 1988, Cadenasso and Pickett 2000, Rooney 2001). Within the
Midwest region of the United States, overabundant deer populations in natural areas are
particularly problematic where woodlots and forests are surrounded by a matrix dominated by
agriculture and exurban development (Hurley et al. 2012).
In June and July of 2013, I assessed impacts of white-tailed deer herbivory in Eagle’s Crest and
Spring Pond Nature Preserves located within Eagle Creek Park, Indianapolis, Indiana. Eagle’s
Crest NP is a 296 (120 ha) acre forest reserve comprised of ridges and ravines. Spring Pond is a
44 acre (18 ha), poorly-drained forest located east of the Eagle Creek Reservoir. Both preserves
have been closed to hunting since their establishment and concern has grown over the last two
decades about negative impacts of herbivory by white-tailed deer.
Methods
I sampled vegetation in both nature preserves to determine: (1) severity of browse damage
based upon indicator species, (2) woody species composition and stem density in forest
understories, and (3) severity of browse on woody species in forest understories.
Indicator species:
To evaluate browse severity based upon herbaceous indicator species, I followed the methods
of Webster and Parker (2000) and Webster et al. (2001). Specifically, I measured the heights of
three species, jack-in-the-pulpit (Arisaema triphyllum), white baneberry (Actaea pachypoda),
and sweet cicely (Osmorhiza claytonia). I measure all mature plants within variable radius plots

3
(ranging from 10 to 15 m depending upon the abundance focal species) that used the same
center points as permanent plots established by Dolan and Moore (2007a, 2007b; Figure 1).
Jack-in-the-pulpit was abundant at Eagle’s Crest NP and fairly common at Spring Pond NP.
However, I only observed the other two species at Eagle’s Crest, where they were uncommon.
Therefore, I measured all individuals of these two species that I encountered both on and off
the plots. I also performed systematic searches at Spring Pond NP to increase the number of
jack-in-the-pulpit plants measured. I measured a total of 1276 jack-in-the-pulpit plants at
Eagle’s Crest NP and 109 plants at Spring Pond NP. I measured four baneberry and seven sweet
cicely plants at Eagle’s Crest NP. I also noted whether plants were flowering and/or had been
browsed. The gender of jack-in-the-pulpit plants was also identified.
Mean heights were calculated for each species, and adjusted percent cover (APC) was
calculated using regression equations developed by Webster and Parker (2000).
Jack in the pulpit:
APC = 1.234(plant height) - 12.07
Sweet cicely:
APC = 0.704(plant height) – 4.782
White baneberry:
APC = 0.806(plant height) + 8.737
APC was used to estimate the browse damage class of each preserve based upon ranges
provided by Parker and Webster (2000) and Webster et al. (2001).
Understory woody species composition and stem density
A nested plot design was used to sample understory woody vegetation. Stems > 1m in height
(saplings) were tallied by species into two height classes (stems ≥ 1 m tall, but < 2.54 cm dbh
and stems 2.1-10 cm dbh) within a 300 m2 plot (9.77 m radius). Four 4 m2 quadrats positioned
four meters from plot center in cardinal directions were used to tally seedlings (stems < 1 m
height) into two height classes; 0-20 cm and 21-100 cm. Mean density of saplings per hectare
was calculated by height class for each sapling species. Mean density per 100 m2 was
calculated by height class for each seedling species.
Browse severity
Browsing of woody understory stems was assessed in four 2 x 2 m quadrats placed in cardinal
directions four meters from plot center. The techniques of Williamson and Hirth (1985) were

4
Figure 1. (A) Eagle’s Crest and (B) Spring Pond Nature Preserves. Sample grid locations from
Dolan and Moore (2007a, 2007b) are shown for both Preserves. Points A3, A4, B2, G3, G5, G6,
H7, H8, and H9 were sampled at Eagle’s Crest NP. Points A1, A3, A9, C7, C8, C9, F2, and F5 were
sampled at Spring Pond.

5
used to quantify browse of individual twigs. Available (non-browsed) and browsed twigs (0.15
to 1.8 m above ground) were counted by species from up to eight randomly selected individuals
per species. All woody species were quantitatively assessed except spicebush (Lindera
benzoin), a non-preferred species and paw paw (Asimina triloba), a highly non-preferred
species. Based upon field observations, I observed light and intermittent browsing of spicebush
and no browsing of paw paw. Percent of twigs browsed for each species was calculated as a
per plot average.
Results and discussion:
Indicator species
Jack-in-the-pulpit
The mean height of jack-in-the-pulpit plants at Eagle’s Crest NP was 16.1 ± 0.2 cm. When this
value was inserted as the independent variable into the regression equation developed by
Webster and Parker (2000), APC was estimated as 7.8 %, which corresponds to a browse
damage rating of severe (the lowest rating; Webster et al. 2001). The mean height of jack-in-
the-pulpit plants at Spring Pond NP was 13.7 ± 0.5 cm. According to the regression equations
developed by Webster and Parker (2000), this value equates to an APC of 4.8%, which also
corresponds to a damage class rating of severe. This APC rivals the most severely degraded
state parks examined by Webster and Parker (2000) and Webster et al. (2001). Reduced
heights of forest perennials in forests with overabundant deer populations have been widely
observed for palatable species (e.g. Augustine and Frelich 1998, Fletcher et al. 2001, Jenkins et
al. 2007, Koh et al. 2010). However, jack-in-the-pulpit is generally viewed as unpalatable
(Augustine and Jordan 1998, Heckel et al. 2010) and its reduced stature is not likely due to
herbivory. Rather, deer-mediated indirect effects, including increased resistance of surface
soils to water penetration and decreased leaf litter depth, have likely driven reduced height
growth (Heckel et al. 2010). These indirect effects have also likely affected the demography of
jack-in-the-pulpit populations in the two preserves; at Eagle’s Crest, only 1.8% of plants were
flowering, while 2.8% were flowering at Spring Pond. Published rates of flowering for jack-in-
the-pulpit populations in areas without overabundant deer populations range from 35-65%
(Bierzychudek 1982b, Ewing and Klein 1982). While jack-in-the pulpit is not a preferred browse
species, 12.5 % (3 out of 24) of flowering plants were browsed at Eagle’s Crest. In addition, two
out of four female flowers were browsed at Eagle’s Crest NP (Figure 2), as was the only
flowering female plant observed at Spring Pond. In jack-in-the-pulpit populations, flowering
individuals are larger than nonflowering plants, and female flowering plants are larger than
male flowering plants (Bierzychudek 1982a). While not preferred, deer appear to have

6
selectively browsed the largest individuals within the populations of the two nature preserves.
Overall, I observed few large individuals at either preserve (Figure 3).
Of all flowering jack-in-the-pulpit plants, only 16.7% at Eagle’s Crest NP were female. Similarly,
only one out three flowering plants at Spring Pond NP was female. Jack-in-the-pulpit plants are
able to switch gender in response to environmental stress, including defoliation (Bierzychudek
1984). Plants typically switch from female to male or sterile plants because flowering as a male
or not flowering requires less expenditure of stored energy reserves than flowering as a female
plant. The relative scarcity of flowering female plants we observed suggests that herbivory has
caused most individuals from the populations of both nature preserves to revert to either
sterile or male forms. Following the methods of Webster et al. (2001), I measured no plants
less than 8 cm in height. Typically, such small individuals could be new seedlings and not
mature plants. However, considering the very low fecundity of jack-in-the-pulpit across both
reserves (only two out of 1385 plants produced fruit; 0.14%), it is unlikely that these small
individuals were new seedlings.
Figure 2. Female jack-in-the-pulpit
plant browsed by deer in the Eagle’s
Crest Nature Preserve at Eagle Creek
State Park.

7
Other indicator species
Based on the seven sweet cicely plants I measured at Eagle’s Crest NP, I calculated a mean
height of 33.3 ± 4.8 cm. According to the regression equation developed by Webster and
Parker (2000) this value equates to an APC of 18.7%, which corresponds to a damage rating of
heavy-severe. The mean height of white baneberry was 8.8 ± 1.4 (based upon four plants),
which corresponds to an APC of 15.8% and a damage class rating of severe. Because of the low
sample size that resulted from the relative rarity of these species, results should be viewed with
greater caution than those for jack-in-the-pulpit. However, these results do support the
damage class ratings calculated for jack-in-the-pulpit. The low rate of flowering I observed for
Figure 3. Height distributions of jack-in-the-pulpit plants sampled at Eagle’s
Crest and Spring Pond Nature Preserves. Please note the different scales of
the Y-axes; far fewer plants were found at Spring Pond NP.

8
these two species (one out of seven sweet cicely plants and none out of four baneberry plants)
also supports the results observed for jack-in-the-pulpit.
Understory woody species composition and stem density
Eagle’s Crest Nature Preserve
The lower stratum (0-20 cm height) of the seedling layer (all woody stems < 1 m height) at
Eagle’s Crest NP was heavily dominated by white ash (Fraxinus americana; 49 ± 21
stems/100m2) with a lower density of sugar maple (Acer saccharum; 9 ± 6 stems/100m2). The
other five species surveyed consisted of individual or scattered seedlings (Figure 4). In the
upper stratum (21-100 cm), paw paw was the dominant species, followed by white ash and
spicebush.
Figure 4. Density (mean ± 1 SE) by species of woody seedlings (stems < 1 m
height) per 100 m2 at
Eagle’s Crest Nature Preserve.

9
Paw paw and spicebush dominated the small sapling class (stems ≥ 1 m height, but ≤ 2.5 cm
dbh) at Eagle’s Crest NP. Paw paw is highly unpalatable to deer and this species has been
observed dominating understories of other forests under chronic heavy browse pressure
(Webster and Parker 1997, Asnani et al. 2006, Jenkins 2011). While spicebush may be
negatively affected by browsing (Asnani et al. 2006), other studies have observed minimal
effects (Rossell et al. 2007). The large saplings (2.6-10 cm dbh) class was dominated by sugar
maple and American elm (Ulmus americana). The much lower density of these two species in
the seedling and small sapling classes suggest that additional stems are not recruiting into the
large sapling class (Figures 5 and 6). The mean density of stems in the large sapling class was
282 stems/ha in Eagle’s Crest NP. In an old-growth forest in Ohio without a documented history
of deer overabundance, Cho and Boerner (1991) observed 456 large saplings/ha in non-gap
areas. When all size classes of seedlings and saplings were combined, the total understory
woody stem density at Eagle’s Crest NP was 13,992 stems/ha. For comparison, Goebel and Hix
(1996) observed 81,887 stems/ha in the understory of 110-129 year-old forests in southern
Ohio.
Figure 5. Facing west on plot G5, Eagle’s Crest Nature Preserve, Eagle Creek State Park. The
herbaceous layer is largely dominated by unpalatable species such as sedges (Carex spp.), wild ginger
(Asarum canadense), wood nettle (Laportea canadensis), and wild leek (Allium tricoccum). The woody
understory is dominated by unpalatable species such as paw paw and browse resistant species, such as
white and green ash.

10
Figure 6. Density (mean ± 1 SE) by species of woody saplings (≥ 1 m height, > 10 cm dbh) per
hectare at Eagle’s Crest Nature Preserve.

11
Spring Pond Nature Preserve
Green ash was, by far, the dominant seedling species at Spring Pond NP in both height strata
Figure 7). With the exception of spicebush, other species occurred as single or scattered
individuals. The small sapling class was dominated by green ash, paw paw, and spicebush. Paw
paw, green ash, and sugar maple dominated the large sapling class (Figure 8). However, the
lack of sugar maple in the small sapling class suggests that continued recruitment into the large
sapling class is unlikely (Figure 8).
Figure 7. Density (mean ± 1 SE) by species of woody seedlings (stems < 1 m height) per 100 m2 at
Spring Pond Nature Preserve.

12
Browsing of woody plants
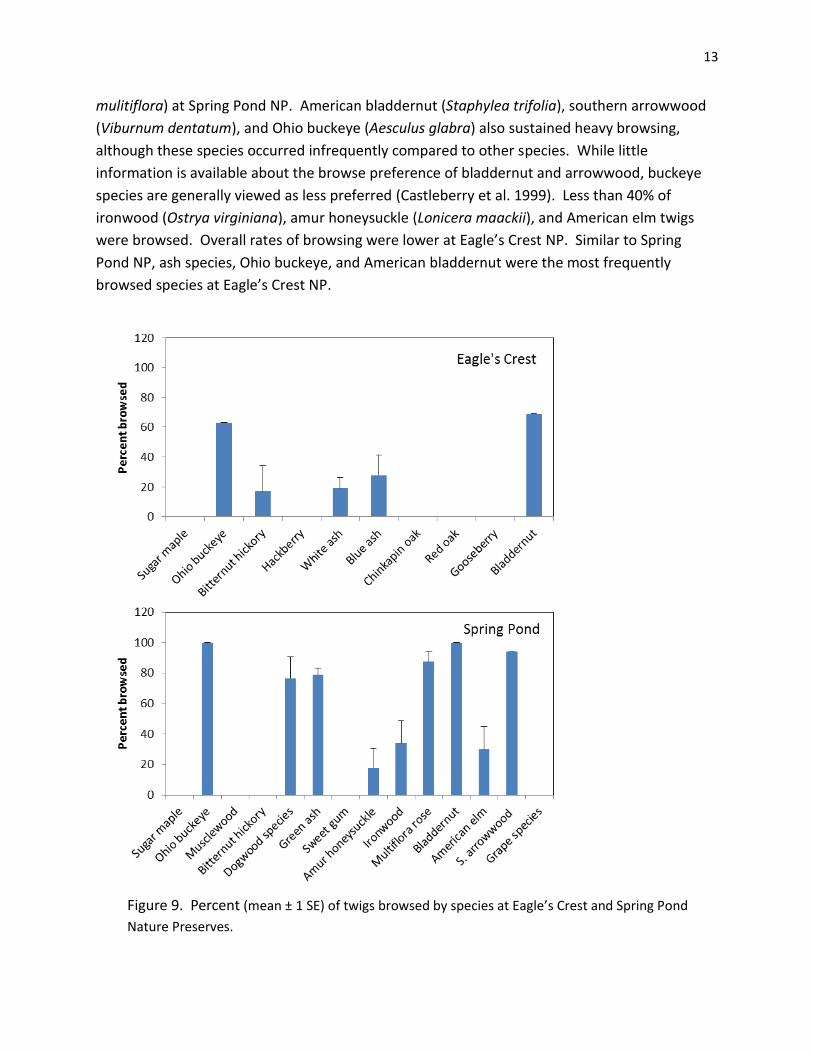
While twigs were browsed at both preserves, the frequency and severity of browsing was much
greater at Spring Pond NP (Figures 9 and 10). Green ash, the dominant seedling and small
sapling species at Spring Pond NP, sustained heavy browsing; 79 ± 4% of all twigs were
browsed. Although green ash was heavily browsed, ash species are tolerant of browsing and
may increase in importance as less-browse tolerant species begin to disappear (Rossell et al
2005). I also observed heavy browsing of dogwood (Cornus) species and multiflora rose (Rosa
Figure 8. Density (mean ± 1 SE) by species of woody saplings (≥ 1 m height, < 10 cm dbh)
per hectare at Spring Pond Nature Preserve.
13
mulitiflora) at Spring Pond NP. American bladdernut (Staphylea trifolia), southern arrowwood
(Viburnum dentatum), and Ohio buckeye (Aesculus glabra) also sustained heavy browsing,
although these species occurred infrequently compared to other species. While little
information is available about the browse preference of bladdernut and arrowwood, buckeye
species are generally viewed as less preferred (Castleberry et al. 1999). Less than 40% of
ironwood (Ostrya virginiana), amur honeysuckle (Lonicera maackii), and American elm twigs
were browsed. Overall rates of browsing were lower at Eagle’s Crest NP. Similar to Spring
Pond NP, ash species, Ohio buckeye, and American bladdernut were the most frequently
browsed species at Eagle’s Crest NP.
Figure 9. Percent (mean ± 1 SE) of twigs browsed by species at Eagle’s Crest and Spring Pond
Nature Preserves.

14
Conclusions
Based upon the results of this study, I conclude that both Eagle’s Crest and Spring Pond Nature
Preserves exhibit degraded plant communities resulting from long-term chronic herbivory by
white-tailed deer. Indicator species analysis based upon earlier work by Webster and Parker
(2000) and Webster et al. (2001) placed both preserves in the severe impact class.
A
C
B
Figure 10. Browse damage on twigs of (A) green ash, (b) multiflora rose, and (C)
Ohio buckeye. All photos were taken in Spring Pond Nature Preserve.

15
Demographic observations of jack-in-the-pulpit suggest a shift in gender and reproductive
status consistent with populations under severe environmental stress. The understories of
both preserves are dominated by unpalatable and browse-resistant species. Overall, the
understory stem densities of both preserves are quite low and reflect the impacts of heavy
long-term browse pressure. This browse pressure is particularly pronounced at Spring Pond NP
where most species exhibited browsing rates greater than 75%.
The low density of canopy species regeneration in the understory, combined with the rarity of
preferred browse species in the herbaceous layer, suggest that the vegetation communities of
both preserves are shifting in both composition and structure towards an alternate state
defined by reduced plant species diversity and reduced recruitment of canopy trees. While
reducing deer populations through controlled hunts is controversial, recent research in Indiana
State Parks suggests that such hunts can successfully reduce deer abundance and allow the
recovery of native plant communities (Jenkins 2011).
Literature Cited
Alverson, W.S., D.M. Waller and S.L. Solheim. 1988. Forests too deer: edge effects in northern Wisconsin. Conservation Biology 2:348–358.
Asnani, K.M., R.A. Klips, and P.S. Curtis. 2006. Regeneration of woodland vegetation after deer
browsing in Sharon Woods Metro Park, Franklin County, Ohio. Ohio Journal of Science 106: 86-92.
Augustine, D.J. and L.E. Frelich. 1998. Effects of white-tailed deer on populations on an
understory forb in fragmented deciduous forests. Conservation Biology 12: 995–1004. Augustine D.J. and P.A. Jordan. 1998. Predictors of white-tailed deer grazing intensity in
fragmented deciduous forests. Journal of Wildlife Management 62:1076-1085. Bierzychudek, P. 1982a. The demography of jack-in-the-pulpit, a forest perennial that changes
sex. Ecological Monographs 52:335–351. Bierzychudek, P. 1982b. Life histories and demography of shade-tolerant temperate forest
herbs: a review. New Phytologist 90: 757-776. Bierzychudek, P. 1984. Determinants of gender in jack-in-the-pulpit: the influence of plant size
and reproductive history. Oecologia 65: 14-18.

16
Bressette, J.W., H. Beck and V.B. Beauchamp. 2012. Beyond the browse line: complex cascade effects mediated by white-tailed deer. Oikos 121: 1749–1760.
Cadenasso, M. L. and S.T.A. Pickett. 2000. Linking forest edge structure to edge function:
mediation of herbivore damage. Journal of Ecology 88:31–44. Castleberry, S.B., M.W. Ford, K.V. Miller, and W.P. Smith. 1999. White-tailed deer browse
preferences in a southern Bottomland Hardwood Forest. Southern Journal of Applied Forestry 78-82.
Cho, D. and R.E.J. Boerner. 1991. Canopy disturbance patterns and regeneration of Quercus
species in two old-growth forests. Vegetatio 93:9–18. Côte’ S.D., T.P. Rooney, J-P Tremblay, C. Dussault, and D.M. Waller. 2004. Ecological impacts of
deer overabundance. Annual Review of Ecology, Evolution, and Systematics 35: 113–147.
Dolan, R. and M. Moore. 2007a. Eleven year resurvey of permanent plots at Spring Pond Nature
Preserve, Eagle Creek Park, Indianapolis, IN. Friesner Herbarium, Butler University, Indianapolis, IN.
Dolan, R. and M. Moore. 2007b. Ten year resurvey of permanent plots at Eagle’s Crest Nature
Preserve, Eagle Creek Park, Indianapolis, IN. Friesner Herbarium, Butler University, Indianapolis, IN.
Ewing, J.W., and R.M. Klein. 1982. Sex expression in Jack-in-the-Pulpit. Bulletin of the Torrey
Botanical Club 109: 47-50. Fletcher, J.D., L.A. Shipley, W.J. McShea, and D.L. Shumway. 2001. Wildlife herbivory and rare
plants: the effects of white-tailed deer, rodents, and insects on growth and survival of Turk’s cap lily. Biological Conservation, 101: 229-238.
Goebel, P.C. and D.M. Hix. 1996. Development of mixed-oak forests in southeastern Ohio: a
comparison of second-growth and old-growth forests. Forest Ecology and Management 84:1-21.
Heckel, C.D., N.A. Bourg, W.J. McShea, and S. Kalisz. 2010. Nonconsumptive effects of a
generalist ungulate herbivore drive decline of unpalatable forest herbs. Ecology 91:319-326.
Hurley, P.M., C.R. Webster, D.J. Flaspohler, and G.R. Parker 2012 Untangling the landscape of
deer overabundance: reserve size vs. landscape context in the agricultural Midwest. Biological Conservation 146: 62–71.

17
Jenkins, L.H. 2011. Evaluating the recovery of vegetation communities in Indiana State Parks after more than a decade of deer population reduction. MS thesis, Purdue University, West Lafayette, Indiana. 173 p.
Jenkins, M.A., C.R. Webster, and J.R. Rock. 2007. Effects of chronic herbivory and historic land
use on population structure of a forest perennial, Trillium catesbaei. Applied Vegetation Science 10: 441-450.
Knight T.M., J.L. Dunn, L.A. Smith, J. Davis, and S. Kalisz. 2009. Deer facilitate invasive plant
success in a Pennsylvania forest understory. Natural Areas Journal 29: 110-116. Koh, S., D.R. Bazely, A.J. Tanentzap, D.R. Voigt, and E. Da Silva. 2007. Trillium grandiflorum
height is an indicator of white-tailed deer density at local and regional scales. Forest Ecology and Management 259: 1472–1479.
Lessard, J.P., W.N. Reynolds, W.A. Bunn, M.A Genung, M.A. Cregger, E. Felker-Quinn, M.N.
Barrios-Garcia, M.L. Stevenson; M. Lawton, C.B. Brown, M. Patrick, J.H. Rock, M.A. Jenkins, J.K. Bailey, and J.A. Schweitzer. 2012. Equivalence in the strength of deer herbivory on above and below ground communities. Basic and Applied Ecology 13: 59-66.
Leopold, A., L.K. Sowls, and D.L. Spencer. 1947. A survey of over-populated deer ranges in the
United States. The Journal of Wildlife Management 11: 162-177. Liang, S.Y. and S.W. Seagle. 2002. Browsing and microhabitat effects on riparian forest woody
seedling demography. Ecology 83: 227. Rooney, T.P. 2001. Deer impacts on forest ecosystems: a North American perspective. Forestry
74: 201-208 Rooney, T.P., S.M. Weigmann, D.A. Rogers, and D.M. Waller. 2004. Biotic impoverishment and
homogenization in unfragmented forest understory communities. Conservation Biology 18: 787–798.
Rossell, C.R., B. Gorsira, and S. Patch. 2005. Effects of White-tailed Deer on vegetation structure
and woody seedling composition in three forest types on the Piedmont Plateau. Forest Ecology and Management 210: 415–424.
Rossell, C.R., S. Patch, S. Salmons. 2007. Effects of deer browsing on native and non-native
vegetation in a mixed oak-beech forest on the Atlantic coastal plain. Northeastern Naturalist 14: 61–72.

18
Russell, F.L., D.B. Zippin, and N.L. Fowler. 2001. Effects of white-tailed deer (Odocoileus virginianus) on plants, plant populations and communities: a review. American Midland Naturalist 146: 1-26.
Tanentzap, A.J., D.R. Bazely, S. Koh, M. Timciska, E.G. Haggith, T.J. Carleton, and D.A. Coomes.
2011. Seeing the forest for the deer: Do reductions in deer-disturbance lead to forest recovery? Biological Conservation 144: 376-382.
Thiemann, J.A., C.R. Webster, M.A. Jenkins, P.M. Hurley, J.H. Rock, and P.S. White. 2009.
Herbaceous-layer impoverishment in a post-agricultural southern Appalachian landscape. American Midland Naturalist 162: 148-168.
Tilghman, N. G. 1989. Impacts of white-tailed deer on forest regeneration in northwestern
Pennsylvania. Journal of Wildlife Management 53: 524-532. Waller, D.M. and W.S. Alverson. 1997. The white-tailed deer: a keystone herbivore. Wildlife
Society Bulletin 25: 217-226. Webster, C. R., M.A. Jenkins, and G.R. Parker. 2001. A field test of herbaceous plant indicators
of deer browsing intensity in mesic hardwood forests of Indiana, USA. Natural Areas Journal 21: 149-158.
Webster, C.R., M.A. Jenkins, and J.H. Rock. 2005. Long-term response of spring flora to chronic
herbivory and deer exclusion in Great Smoky Mountains National Park, USA. Biological Conservation 125: 297-307.
Webster, C.R. and G.R. Parker. 1997. The effects of white-tailed deer on plant communities
within Indiana state parks. Proceedings of the Indiana Academy of Science 106: 213-231. Webster, C.R. and G.R. Parker. 2000. Evaluation of Osmorhiza claytonii (Michx.) C.B. Clarke,
Arisaema triphyllum (L.) Schott, and Actaea pachypoda Ell. as potential indicators of white-tailed deer overabundance. Natural Areas Journal 20: 176-188.
Webster, C.R., J.H. Rock, R.E. Froese, and M.A. Jenkins. 2008. Drought-herbivory interaction
disrupts competitive displacement of native plants by Microstegium vimineum, 10 year results. Oecologia 157: 497–50.
Williamson, S. and D.H. Hirth. 1985. An evaluation of edge use by white tailed deer. Wildlife
Society Bulletin 13:252-257.


















