
2 Pseudomonas 3 4 ACCEPTED - Journal of...
42
1 1 Organization and PprB-dependent control of the Pseudomonas 2 aeruginosa tad locus involved in Flp pilus biology 3 4 Christophe S. Bernard, Christophe Bordi, Elise Termine, Alain Filloux † and 5 Sophie de Bentzmann * 6 7 Membrane complexes and pathogenicity in Pseudomonas aeruginosa, 8 CNRS LISM (UPR9027)- Aix-Marseille Université, 31 Chemin Joseph Aiguier, 13402 9 Marseille Cedex 20, France. † Present address: Centre for Molecular Microbiology and 10 Infection, Division of Cell and Molecular Biology, Flowers Building, South Kensington 11 Campus, Imperial College London, London SW7 2AZ, UK. 12 13 * To whom correspondence should be addressed; e-mail: [email protected] 14 15 Running title: P. aeruginosa Tad machine 16 17 Key words: Type IVb pili, Flp pilin, Two-component system, PprA-PprB, RcpC, 18 pseudopilin. 19 ACCEPTED Copyright © 2009, American Society for Microbiology and/or the Listed Authors/Institutions. All Rights Reserved. J. Bacteriol. doi:10.1128/JB.01330-08 JB Accepts, published online ahead of print on 16 January 2009 on May 25, 2018 by guest http://jb.asm.org/ Downloaded from
Transcript of 2 Pseudomonas 3 4 ACCEPTED - Journal of...

1
1
Organization and PprB-dependent control of the Pseudomonas 2
aeruginosa tad locus involved in Flp pilus biology 3
4
Christophe S. Bernard, Christophe Bordi, Elise Termine, Alain Filloux† and 5
Sophie de Bentzmann* 6
7
Membrane complexes and pathogenicity in Pseudomonas aeruginosa, 8
CNRS LISM (UPR9027)- Aix-Marseille Université, 31 Chemin Joseph Aiguier, 13402 9
Marseille Cedex 20, France.†
Present address: Centre for Molecular Microbiology and 10
Infection, Division of Cell and Molecular Biology, Flowers Building, South Kensington 11
Campus, Imperial College London, London SW7 2AZ, UK. 12
13
*To whom correspondence should be addressed; e-mail: [email protected] 14
15
Running title: P. aeruginosa Tad machine 16
17
Key words: Type IVb pili, Flp pilin, Two-component system, PprA-PprB, RcpC, 18
pseudopilin. 19
ACCEPTED
Copyright © 2009, American Society for Microbiology and/or the Listed Authors/Institutions. All Rights Reserved.J. Bacteriol. doi:10.1128/JB.01330-08 JB Accepts, published online ahead of print on 16 January 2009
on May 25, 2018 by guest
http://jb.asm.org/
Dow
nloaded from

2
Abstract (189 words) 1
Bacterial attachment to the substratum involves several cell surface organelles, including 2
various types of pili. The Pseudomonas aeruginosa Tad machine assembles type IVb pili, 3
which are required for adhesion to abiotic and to eukaryotic cells. Type IVb pili consist of a 4
major subunit, the Flp pilin, processed by the FppA prepilin peptidase. In this study, we 5
investigated the regulatory mechanism of the tad locus. We showed that the flp gene is 6
expressed late in the stationary growth phase in aerobic conditions. We also showed that the 7
tad locus was composed of five independent transcriptional units. We used transcriptional 8
fusions to show that tad gene expression was positively controlled by the PprB response 9
regulator. We subsequently showed that PprB bound to the promoter regions, directly 10
controlling the expression of these genes. We then evaluated the contribution of two genes, 11
tadF and rcpC, to type IVb pilus assembly. The deletion of these two genes had no effect on 12
Flp production, pilus assembly or Flp-mediated adhesion to abiotic surface in our conditions. 13
However, our results suggest that the putative RcpC protein modifies the Flp pilin, thereby 14
promoting Flp-dependent adhesion to eukaryotic cells. 15
16
17
18
ACCEPTED
on May 25, 2018 by guest
http://jb.asm.org/
Dow
nloaded from

3
Introduction 1
Pseudomonas aeruginosa is an opportunistic pathogen responsible for severe 2
infections in immunocompromised patients and individuals suffering from cystic fibrosis. 3
Postgenomic studies have focused attention on the ability of P. aeruginosa to bind to various 4
surfaces. This binding involves combinations of type IVa (4) and type IVb pili (2), flagella 5
and Cup fimbriae (33, 27) on the surface of the bacterium, the release of adhesive proteins via 6
secretion systems (9) and the production of an exopolysaccharide matrix (10, 23, 16, 35). 7
Most type IVb pilins have similar characteristics (4). The 15- to 30-residue leader 8
peptide of the pre-pilin is processed downstream from a conserved hydrophobic residue, 9
generating a mature pilin of ~ 190 amino acids in length. The resulting exposed D region in 10
assembled type IVb pili is large. The type IVb pilin of Vibrio cholerae, TcpA, assembles as a 11
left-handed three-start helix with six subunits per turn (4). These features result in type IVb 12
pili being thicker than type IVa pili. The type IVb pilin family includes a clearly 13
monophyletic Flp (fimbrial low-molecular weight protein) prepilin subfamily, initially 14
described in Aggregatibacter (Actinobacillus) actinomycetemcomitans. Members of this 15
subfamily have a unique set of features in common: a long leader peptide, a short mature pilin 16
(50 to 80 aa), and an Flp motif consisting of 20 hydrophobic residues at the N terminus of the 17
mature pilin, with adjacent glutamate and tyrosine residues at the centre of this motif (20). 18
The flp gene is linked to the tad (tight adhesion) locus, which encodes a macromolecular 19
machine dedicated to the assembly of adhesive pili of the type IVb Flp subfamily (18, 19, 25). 20
Similar tad loci have been identified in Archaea, actinobacteria and Gram-negative bacteria, 21
usually as a single copy, but in up to four copies in some species (20, 32). Several conserved 22
elements have been identified in the Tad machines to date: the TadA trafficking NTPase (18, 23
24), the RcpA secretin, one or two inner membrane PilC homologues, TadB and TadC, the 24
TadV/FppA prepilin peptidase (2, 31), the Flp pilin and the TadE and TadF pseudopilins (31). 25
ACCEPTED
on May 25, 2018 by guest
http://jb.asm.org/
Dow
nloaded from

4
These components are also shared by type IVa pilus machines (T4P) and type II secretion 1
systems (T2S). However, the presence within the tad locus of several specific genes encoding 2
proteins with no homologue in the T4P and T2S systems, such as RcpC, RcpB, TadZ and 3
TadG, suggests that Tad machines represent a new type of prokaryotic secretion system (32). 4
The tad genes are usually organised linearly in a single direction, suggesting that they may 5
constitute an operon. This is likely, as the tad locus of A. actinomycetemcomitans is 6
transcribed as a polycistronic mRNA (11). Classical tad loci start with the flp gene, followed 7
by the gene encoding the prepilin peptidase (tadV) and the remaining genes encoding the Tad 8
machine elements. The tad locus of P. aeruginosa has a distinctive genetic organisation, with 9
four or five putative transcriptional units operating in different directions: the flp gene, nine 10
genes (rcpC-tadG) constituting the rcp-tad locus, the gene encoding the TadV/FppA prepilin 11
peptidase, the gene encoding the response regulator PprB and the tadF gene (2). The 12
divergent orientations of the flp and rcp-tad genes and the requirement for flp overexpression 13
for efficient Flp pilus assembly (2) suggest that different regulatory processes are involved in 14
Flp production, maturation and assembly. Environmental conditions, such as O2 and CO2 15
levels and nutrient availability have been reported to affect Flp production in Aggregatibacter 16
aphrophilus and A. actinomycetemcomitans (28, 32). In P. aeruginosa, global approaches 17
have shown that some tad genes are either regulated by quorum sensing (29, 39, 40) or under 18
the control of the VqsR master regulator (17), or the MvaT transcriptional regulator (34). The 19
fppA and tadF genes are located between the pprA and pprB genes. These genes encode a 20
classical two-component system (TCS) that was thought to act as a master regulator of 21
quorum sensing (QS) (36, 6) until the recent identification of a secondary mutation in the 22
LasR regulator in the transposon pprB mutant (7). Nevertheless, this TCS has been reported to 23
play a key role in membrane permeability regulation and drug resistance in P. aeruginosa 24
ACCEPTED
on May 25, 2018 by guest
http://jb.asm.org/
Dow
nloaded from

5
(41). The proximity of this TCS to the rcp-tad locus suggests it may be involved in the 1
transcriptional regulation of type IVb pilus gene expression. 2
We determined the conditions under which the flp gene was expressed in P. 3
aeruginosa and showed that flp gene expression occurred late in the stationary growth phase 4
in aerobic conditions. We determined the operon structure of the tad locus and assessed the 5
potential regulatory role of the TCS PprAB. We demonstrated that the tad locus was 6
organised into five transcriptional units and positively regulated by the PprB response 7
regulator. We also studied the products of several genes encoding components of the Tad 8
machine, including TadF, which is probably the only pseudopilin in this system, and RcpC, a 9
putative protein from a family of bacterial proteins containing two β-clip domains and unique 10
to Tad machines. 11
ACCEPTED
on May 25, 2018 by guest
http://jb.asm.org/
Dow
nloaded from

6
Materials and Methods 1
Bacterial strains and growth conditions. The strains and plasmids used in this study are 2
listed in Table 1. Strains were grown at 30°C or 37°C in L broth, in the presence of 100 µM 3
isopropyl-β-D–thiogalactopyranoside (IPTG). The E. coli TG1, MG1655, TOP10F’ and CC118 4
(λpir) strains were used for standard genetic manipulations. Recombinant plasmids were 5
introduced into P. aeruginosa, using the conjugative properties of pRK2013. Transconjugants 6
were selected on Pseudomonas isolation agar (PIA) (Difco Laboratories) supplemented with 7
appropriate antibiotics. The following antibiotic concentrations were used: 1) for E. coli: 8
ampicillin (Ap), 50 µg/ml; kanamycin (Km), 25 µg/ml; tetracycline (Tc), 15 µg/ml;
9
streptomycin (Sm), 50 µg/ml; 2) for P. aeruginosa: carbenicillin (Cb), 250 µg/ml; tetracycline 10
(Tc), 200 µg/ml; streptomycin (Sm), 2 mg/ml. 11
12
Construction of P. aeruginosa deletion mutants. PCR was used to generate a 550 bp DNA 13
fragment upstream (Up) from the pprB, rcpC, tadF genes with the (DelPBUp5, DelPBUp3) 14
(DelRCUp5, DelRCUp3), (DelTFUp5, DelTFUp3) oligonucleotide pairs, respectively (Table 15
2) and a 550 bp DNA fragment downstream (Dn) from the pprB, rcpC, tadF genes, using the 16
(DelPBDn5, DelPBDn3), (DelRCDn5, DelPRCDn3) (DelTFDn5, DelTFDn3) oligonucleotide 17
pairs, respectively, (Table 2). The resulting DNA fragments were inserted separately into the 18
pCR2.1 vector. The Up and Dn DNA fragments for the pprB, rcpC and tadF genes, bearing 19
appropriate sites — SacI/KpnI, KpnI/BamHI — were digested and inserted into the suicide 20
vector pEX18, previously digested with SacI/BamHI, via a three-partner procedure. The 21
recombinant plasmids were introduced into P. aeruginosa and the deletion mutants were 22
obtained by double selection on LB-agar supplemented with irgasan (25 µg/ml) and 23
carbenicillin (250 µg/ml) at 37°C and NaCl-free LB agar containing 6% sucrose at 30°C (21). 24
25
ACCEPTED
on May 25, 2018 by guest
http://jb.asm.org/
Dow
nloaded from

7
Chromosomal transcriptional fusions. DNA fragments containing the putative promoter 1
regions between the flp gene and the rcp-tad locus (501 bp) and the fppA and the tadF genes 2
(150 bp) were amplified by PCR, using the (PromFlRUp PromFlRDn) and (PromFpTUp, 3
PromFpTDn) oligonucleotide pairs, respectively (Table 2). The DNA fragments were inserted 4
into pCR2.1, excised by digestion with KpnI/XhoI or XhoI/HindIII and inserted into the 5
miniCTX-lacZ vector, which is a mini-CTX1 derived vector�(1) — generating the miniCTX-6
pflp-lacZ and miniCTX-prcpC-lacZ constructs — or inserted into the BamHI/XhoI or 7
XhoI/KpnI sites of the miniCTX-lacZ vector, thus generating the miniCTX-pfppA-lacZ and 8
miniCTX-ptadF-lacZ constructs. These plasmids were used to generate chromosomal flp-9
lacZ, rcpC-lacZ, fppA-lacZ and tadF-lacZ fusions in the PAO1 strain, as previously described 10
(12, 13). The expression of the various transcriptional fusions was monitored by assaying 11
galactosidase activity in at least three independent experiments. 12
13
Expression of tad fusions in the E. coli heterologous host. Both plasmids — i.e., miniCTX-14
pflp-lacZ, miniCTX-prcpC-lacZ, miniCTX-pfppA-lacZ or miniCTX-ptadF-lacZ and 15
pMMB67-HE or pMMBpprB vectors— were introduced into the MG1655 strain of E. coli by 16
transformation. After double selection on agar plates containing appropriate antibiotics, cells 17
were cultured and induced by incubation with 0.1 mM IPTG for 1 h. The expression of the 18
various transcriptional fusions was monitored by assaying galactosidase activity in three 19
independent experiments. 20
21
Cloning procedures for the rcpC and pprB genes. A 1411 bp DNA fragment encompassing 22
the putative promoter region of the rcpC gene upstream from the rcp-tad locus and the rcpC 23
gene itself was amplified by PCR with the PromFlRDn and RcpCDn oligonucleotides (Table 24
2). It was inserted into pCR2.1, excised by HindIII/XhoI digestion and inserted into the 25
miniCTX-lacZ vector, thus generating the miniCTX-rcpC construct. This construction was 26
used for cis-complementation experiments in the rcpC mutants, to generate the PAO1∆rcpC 27
ACCEPTED
on May 25, 2018 by guest
http://jb.asm.org/
Dow
nloaded from

8
attB::rcpC and PAO1∆pilA∆fliC∆rcpC attB::rcpC strains. By contrast to the procedure 1
described for lacZ fusions, the FRT cassette-excision step was not performed, resulting in the 2
generation of a strain with tetracycline resistance for the monitoring of complementation. The 3
pprB gene was amplified by PCR, using the PprBUp and PprBDn oligonucleotides (Table 2). 4
It was inserted into pCR2.1 and then excised and inserted between the XbaI and SacI sites of 5
the broad-host range vector pMMB67-HE. 6
7
Electrophoretic mobility shift assays. The His-tagged version of the PprB (PprB-6His) 8
protein was produced in E. coli M15 strain after induction with 0.5 mM IPTG for 4 h at 28°C, 9
and was purified as recommended (41). Electrophoretic mobility shift assays were performed 10
as followed: the intergenic DNA regions encompassing the promoter regions of the five 11
transcriptional units identified in the tad locus were amplified by PCR with the corresponding 12
oligonucleotide pairs (PromFlRUp, PromFlRDn), (PromFpTlUp, PromFpTlDn), (4296-13
95U40. 4296-95R), generating DNA fragments of 501, 561, and 560 bp for the three 14
intergenic regions upstream from the flp-rcp, fppA-tadF and pprB genes, respectively. The 15
360 bp DNA region corresponding to the promoter region of the rsmZ gene was amplified by 16
PCR with the PromrsmZUp and PromrsmZDn oligonucleotides and was used as a negative 17
control. The mixture of PCR products obtained, containing the intergenic DNA region of 18
interest and the negative control (1:2) in a 50 mM Tris-HCl buffer pH 8.2 containing 1 mM 19
EDTA and 0.25 mM saccharose, was incubated for 30 minutes at room temperature with 20
various amounts of purified PprB-6His protein, ranging from 0 to 2.34 µM. The 21
corresponding mixtures were run on native 12 % acrylamide gels and stained with ethidium 22
bromide. 23
24
Tris-glycine gel electrophoresis and western blot analysis. Flp production was analysed by 25
electrophoresis in a 16.5% polyacrylamide Tris-glycine gel, as previously described (2). 26
Bacterial cells were cultured and mixed with loading buffer (the equivalent of 0.025 OD600 27
ACCEPTED
on May 25, 2018 by guest
http://jb.asm.org/
Dow
nloaded from

9
units per µl). The samples were treated with benzonase (1 unit /µl) for 30 minutes at 37°C, 1
boiled for 10 minutes, and the proteins were separated by electrophoresis. Proteins were 2
blotted onto nitrocellulose membranes. Flp protein was immunodetected with the polyclonal 3
antibody against Flp, used at a dilution of 1:150. A peroxidase-conjugated goat anti-rabbit 4
immunoglobulin G was used at a dilution of 1:5,000 in Tris-buffered saline supplemented 5
with 10% milk and 0.1% Tween for the detection of primary antibody binding to the Flp 6
protein. 7
8
Transmission electron microscopy. Aliquots of bacteria were collected for transmission 9
electron microscopy (TEM). The bacterial pellet was obtained by centrifugation and 10
resuspended in 0.15M NaCl in 10 mM Tris pH 7.8 (Tris-NaCl). A drop of the bacterial 11
suspension was placed on Formvar- and carbon-coated copper grids and left for 12
approximately 5 minutes. Grids were then fixed by incubation with 4% paraformaldehyde for 13
5 minutes and rinsed twice with Tris-NaCl, for 5 minutes each. Grids were then incubated 14
with 5% bovine serum albumin (BSA) in Tris-NaCl for 10 minutes and then for 45 minutes 15
with the Flp antiserum at a dilution of 1:150 in 0.5% BSA in Tris-NaCl. Grids were finally 16
incubated for 30 minutes with 10 nm colloidal gold-conjugated protein A in 0.5% BSA in 17
Tris-NaCl. Grids were washed several times in Tris-NaCl and then in water, and were then 18
immersed in a drop of 1% uranyl acetate for 1 minute. Grids were examined in a JEOL 19
1200EX transmission electron microscope operating at 80.0 kV. 20
The length of Flp pili observed in bacteria with different genetic backgrounds was 21
determined by measurements on 30 pictures for each strain, systematically acquired from at 22
least three independent experiments. Statistical analysis was based on unpaired t-tests 23
(GraphPad Prism 4 software). 24
25
Biofilm formation and confocal microscopy. Bacteria were grown in M63 medium 26
supplemented with 0.4% L-arginine and 1 mM MgSO4, at 30°C, without shaking. Coverslips 27
ACCEPTED
on May 25, 2018 by guest
http://jb.asm.org/
Dow
nloaded from

10
were partially immersed in the bacterial cultures in Falcon tubes and were then processed for 1
confocal microscopy. Before observation, coverslips were fixed by incubation with 4% 2
paraformaldehyde and stained by incubation with 4’,6-diamidino-2 phenylindole (DAPI) for 3
15 minutes. Slides were observed in an Olympus FV-1000, using a laser diode for excitation 4
at 405 nm. Images were captured and processed with Fluoview and Amarys software. 5
6
Assay of adhesion to the surface of bronchial epithelial cells. 16HBE14o- human bronchial 7
epithelial cells were incubated in 24-well microplates containing coverslips in minimal 8
essential medium supplemented with 10% foetal calf serum and antibiotics for 24 h at 37°C, 9
under an atmosphere containing 5% CO2. Four hours before infection, the medium was 10
replaced by serum- and antibiotic-free medium. The epithelial cells were infected with 11
bacteria at a multiplicity of infection of 30, for a period of 4 h at 37°C. The samples were 12
rinsed twice with PBS, fixed with 4% formaldehyde, stained with 0.1% crystal violet for 5 13
minutes, washed twice with water, dried, mounted and sealed with Eukitt mounting medium 14
(EMS), and observed with an Axioscop 40 microscope (Zeiss). The bacteria adhering to 15
epithelial cells (30 epithelial cells/assay) were counted in randomly chosen microscopic fields 16
from three independent assays. The data obtained for each genetic background were 17
compared, in a one way-analysis of variance and through unpaired t-tests (GraphPad Prism 4 18
software). 19
20
Isolation of RNA and RT-PCR. The PAO1 strain, into which pMMBpprB was introduced, 21
was cultured with 0.1 mM IPTG for 6 h at 37°C. Total cellular RNA was isolated, using the 22
PureYield RNA Midiprep System (Promega). RT-PCR was carried out with the Access RT-23
PCR System (Promega), according the manufacturer's instructions, but with the addition of 24
6% DMSO to the RT-PCR mixture. We used a total of 1.8 µg of RNA or genomic DNA. RT-25
PCR was carried out with gene-specific primers overlapping junction regions between 26
adjacent genes (listed in Table 2 and in Fig. 1A), with a T1 thermocycler (Biometra), using 27
ACCEPTED
on May 25, 2018 by guest
http://jb.asm.org/
Dow
nloaded from

11
the following protocol: reverse transcription for 45 minutes at 45°C, inactivation of reverse 1
transcriptase by incubation at 94°C for 2 minutes, followed by 45 cycles of PCR amplification 2
with heating at 94°C for 30 s, 60°C for 1 minute and 68°C for 2 minutes. We checked that 3
RNA preparations were not contaminated with DNA, by carrying out the same experiment 4
without adding the reverse transcriptase. The expected sizes of the amplicons for the various 5
pairs of primers were 1: primer 20-primer 21: 460 bp, 2: 22-23: 480 bp, 3: 24-25: 460 bp, 4: 6
26-27: 370 bp, 5: 28-29: 540 bp, 6: 30-31: 460 bp, 7: 32-33: 390 bp, 8: 34-35: 450 bp, 9: 36-7
37: 300 bp (positive control), 10: 36-38: 390 bp (negative control), 11: 39-40: 560 bp and 12: 8
41-42: 590 bp. 9
ACCEPTED
on May 25, 2018 by guest
http://jb.asm.org/
Dow
nloaded from

12
Results 1
Insight into the genetic organisation of the P. aeruginosa tad locus 2
The P. aeruginosa tad locus is characterised by the divergent transcription of the flp gene 3
encoding the prepilin Flp and the rcp-tad locus containing genes encoding the assembly 4
machine, including the secretin RcpA, the ATPase TadA and the two PilC homologues, TadB 5
and TadC (Fig. 1A). We previously identified and characterised the product of the fppA gene: 6
the prepilin peptidase of the Flp assembly machine (2). The PA4294 ORF, for which no 7
putative homologue has been identified, is adjacent to the fppA gene but is transcribed in the 8
opposite direction. However, careful examination revealed that this ORF had another putative 9
start codon, 42 nucleotides downstream from that identified at http://www.pseudomonas.com. 10
The ORF transcribed from this start codon encodes a putative TadF protein homologue, a 11
154-residue pseudopilin containing the conserved GAVXIEF sequence, the putative cleavage 12
site being located after the G residue (31). The fppA and tadF genes are flanked by the 13
PA4293 and PA4296 genes encoding the histidine kinase (HK) PprA and its cognate response 14
regulator (RR) PprB, respectively (Fig. 1A). This two-component system regulates membrane 15
permeability and confers aminoglycoside sensitivity (41). The classical sensor PprA is a 922-16
amino acid protein with no predicted transmembrane domain. However, this protein does 17
have one GAF domain (a domain present in phytochromes and cGMP-specific 18
phosphodiesterases, in Anabaena adenylate cyclases and in E. coli FhlA protein), two PAS 19
domains (domain present in the period circadian protein (Per), the Ah receptor nuclear 20
translocator protein (Arnt) and in the single-minded protein (Sim)) separated by three PAC 21
(PAS-associated C-terminal) domains and one transmitter domain (H) (Fig. 1B) (36). The 22
response regulator PprB, a member of the LuxR family (6), is a 275-amino acid protein with a 23
receiver domain (REC) and a DNA-binding, helix-turn-helix (HTH) domain (Fig. 1B). 24
25
The tad locus is organised into five transcriptional units 26
ACCEPTED
on May 25, 2018 by guest
http://jb.asm.org/
Dow
nloaded from

13
In silico analysis carried out with Operon Finding 2.1 software at 1
http://www.pseudomonas.com predicted co-transcription of the rcpC to tadG genes. We thus 2
investigated whether the rcpC-tadG, fppA-pprB and tadF-pprA loci were transcribed as 3
polycistronic mRNA, using oligonucleotides designed to amplify regions spanning gene 4
junctions (numbered 1 to 12, Fig. 1A and Fig. 1C) on cDNA obtained by the reverse 5
transcription of extracted mRNA (Fig. 1C, lower panel). Control experiments were carried out 6
on genomic DNA (Fig. 1C, upper panel) and RNA (Fig. 1C, middle panel). Co-transcribed 7
genes give a signal of the same size on the cDNA and DNA panels. As shown in Fig. 1C, this 8
was the case for rcpC/rcpA (lane 1), rcpA/tadZ (lane 2), tadZ/tadA (lane 3), tadA/tadB (lane 9
4), tadB/tadC (lane 5), tadC/tadD (lane 6), tadD/PA4298 (lane 7), PA4298/tadG (lane 8) and 10
tadF/pprA (lane 12). A positive internal control within the tadG gene (lane 9) and a negative 11
control between the two divergent genes tadG and pprB (lane 10) were included. Our results 12
demonstrate the transcription of the rcpC-tadG locus as a polycistronic mRNA, constituting 13
transcriptional unit II. The tadF and pprA genes were also co-transcribed, constituting 14
transcriptional unit V, with transcriptional unit I consisting of the flp gene. By contrast, two 15
independently transcribed genes generate a signal on the DNA panel but not on the cDNA 16
panel. This was the case for the fppA and pprB genes (lane 11), which thus form independent 17
transcriptional units, referred to as IV and III, respectively. The lack of an amplification signal 18
(middle panel) in the absence of reverse transcriptase confirmed that RNA samples were not 19
contaminated by genomic DNA. Overall, these results demonstrate that the P. aeruginosa tad 20
locus is organized into five transcriptional units (I-V, Fig. 1A). 21
22
The Flp pilin is produced late in the stationary growth phase 23
In a previous study, we found that Flp was produced late in the growth phase, but this was not 24
strictly reproducible between cultures (2). We inserted the putative promoter region upstream 25
from the flp gene into the mini-CTX-lacZ vector, generating miniCTX-pflp-lacZ. This 26
construct was introduced into the chromosome of the PAO1 strain, to generate the PAO1/flp-27
ACCEPTED
on May 25, 2018 by guest
http://jb.asm.org/
Dow
nloaded from

14
lacZ strain. Expression peaked when the bacteria had been cultured at 37°C in L broth with 1
vigorous shaking, and then subcultured under aerobic conditions, with vigorous shaking, at 2
30°C. In these conditions, expression of the flp-lacZ fusion was maximal in the late stationary 3
growth phase (Fig. 2A). Indeed, immunodetection of the Flp protein with specific anti-Flp 4
antibodies showed that Flp was not produced during the exponential growth phase, was barely 5
detectable at early stationary phase and produced in significant amounts during late stationary 6
growth phase (Fig. 2B). We then investigated whether Flp subunits produced in these 7
conditions assembled into type IVb pili. Using anti-Flp antibodies, we showed by 8
immunogold labelling and TEM that Flp pili were formed at the cell surface (Fig. 2C). The 9
appearance and number (one per cell) of the type IVb pili assembled when Flp was produced 10
from a gene inserted into the chromosome (Fig. 2C) were identical to those observed when 11
Flp was overproduced from a replicative plasmid (reference 2, data not shown). Thus, the Flp 12
pilin was produced under aerobic conditions, late in the growth phase, and was assembled into 13
a single type IVb pilus. 14
15
The PprB response regulator controls tad gene expression 16
Based on the particular location of the pprA and pprB genes within the tad locus and on the 17
co-transcription of the pprA and tadF genes, we further investigated the effect of the PprA-18
PprB TCS on expression of the transcriptional units corresponding to genes encoding the Tad 19
machine (I, II, IV and V). We constructed four different reporter PAO1 strains, corresponding 20
to the various transcriptional units defined above and designated flp-lacZ (I), rcpC-lacZ (II), 21
fppA-lacZ (IV) and tadF-lacZ (V), respectively. We monitored relative expression levels in 22
the presence and absence of inducible pprB overexpression. Overproduction of the PprB 23
regulator led to early expression of the flp-lacZ fusion (I) in the growth phase (Fig. 3A). The 24
overproduction of PprB increased the level of expression of the rcpC-lacZ fusion (II) to levels 25
up to 20 times as high as those in the control strain (Fig. 3B). Moreover, PprB seems to be the 26
main regulator controlling flp expression, as expression of the flp-lacZ fusion was very weak 27
ACCEPTED
on May 25, 2018 by guest
http://jb.asm.org/
Dow
nloaded from

15
throughout the growth phase in a pprB mutant and was restored to parental levels when the 1
pprB gene was introduced in trans (Fig. 3C). Similarly, expression of the rcpC-lacZ fusion 2
was very weak in the pprB deletion mutant, reaching no more than 50 Miller units, 3
corresponding to the basal level of expression for the empty vector (mini-CTX-lacZ 4
integrated into the chromosome at attB sites but without a promoter region, data not shown), 5
but was restored to parental levels by introducing the pprB gene in trans (Fig. 3C). The rcpC-6
lacZ fusion had a level of activity in the wild-type (WT) PAO1 background only one fiftieth 7
that of the flp-lacZ fusion (compare scales in Fig. 3). However, this level of expression was 8
sufficient for the assembly of a single pilus at the cell surface. 9
Expression of the fppA gene was also essentially dependent on the PprB response 10
regulator, as the level of expression of the fppA-lacZ fusion (IV) in the pprB mutant was only 11
one sixth that in the WT strain (Fig. 3C). The expression of the fppA-lacZ fusion was only 12
partially restored by introducing the pprB gene in trans. The tadF-lacZ fusion (V) was much 13
less strongly expressed in the WT strain than the other lacZ fusions and was therefore slightly 14
affected in the pprB mutant (Fig. 3C). However, the expression of this fusion was induced by 15
the overproduction of PprB in the pprB mutant. Thus, the PprB response regulator of the 16
PprA-PprB TCS positively controls the expression of these four transcriptional units of 17
the tad locus, although the PprB-dependent activation of transcriptional unit V was detectable 18
only under conditions of pprB overexpression. To validate this conclusion, the PprB-19
dependent control of the tad locus was investigated in the heterologous host E. coli, in which 20
tad gene-related transcriptional fusions (I, II, IV and V) were introduced and the gene 21
encoding the response regulator was overexpressed. In the E. coli MG1655 strain, PprB 22
overproduction resulted in levels of expression of tad transcriptional fusions 50, 6, 1.7 and 8 23
times higher than those in strains receiving the empty vector pMMB67-HE (Fig. 3D), for 24
transcriptional units I, II, IV and V, respectively. These results are consistent with a model in 25
which the tad locus is directly controlled by the response regulator PprB. 26
27
ACCEPTED
on May 25, 2018 by guest
http://jb.asm.org/
Dow
nloaded from

16
The PprB response regulator binds to the tad intergenic regions 1
To investigate whether PprB-dependent tad locus regulation involves the binding of PprB to 2
the three intergenic regions of the tad locus, we therefore carried out electrophoretic mobility 3
shift assays (EMSA) with a purified His-tagged PprB protein (PprB-6His). These three 4
intergenic regions may contain the flp, rcpC, fppA, tadF and pprB promoters. PprB-6His was 5
purified from E. coli extracts, as previously reported (41) and was essentially present in its 6
unphosphorylated form during EMSA assays. Fig. 4 shows that PprB bound to the three 7
intergenic DNA regions upstream from the flp/rcp, tadF/fppA and pprB genes. The absence of 8
a mobility shift for the 360 bp DNA fragment corresponding to the region upstream from the 9
rsmZ gene demonstrates the specificity of PprB binding to the target tad DNA regions (Fig. 10
4A-C). For the tadF/fppA intergenic region, similar results were obtained when the 150-bp 11
region used in the transcriptional fusion was tested by EMSA instead of the 561-bp DNA 12
fragment (data not shown). For the two intergenic DNA regions carrying two potential 13
promoters in functioning in opposite directions (flp/rcp, Fig. 4A; and tadF/fppA, Fig. 4B), we 14
observed two retarded complexes at high PprB-6His concentrations. By contrast, for the DNA 15
region carrying the pprB promoter, we observed only one retarded complex (Fig. 4C). 16
17
The PprB response regulator controls Flp production and assembly 18
We investigated the impact of PprB overproduction on Flp production. As expected from the 19
effect on flp-lacZ fusion activity, PprB overproduction did not significantly increase Flp 20
production. However, PprB overproduction did affect the electrophoretic mobility (EM) of 21
this protein (Fig. 5A). Flp migrated with a higher apparent molecular weight in the presence 22
of excess PprB than in the control strain. No Flp production was detected in the pprB mutant, 23
whereas the protein was detected, albeit at lower levels than for the WT strain, following 24
trans-complementation with the pprB gene (Fig. 5A). The PprB-dependent Flp production 25
observed is consistent with the lack of expression of the flp-lacZ transcriptional fusion in the 26
PAO1∆pprB strain (See Fig. 3C). No Flp pilus was detected in the pprB mutant, but Flp pili 27
ACCEPTED
on May 25, 2018 by guest
http://jb.asm.org/
Dow
nloaded from

17
were recovered at the cell surface of pprB mutant cells transformed with pMMBpprB (Fig. 1
5B). However, these pili were significantly shorter (0.56 µm ± 0.14) than WT Flp pili (2.25 2
µm ± 0.25) (p<0.0001, Fig. 5C). 3
4
Electrophoretic migration of the Flp pilin is modified in an RcpC-dependent manner 5
As this PprB-dependent EM shift might potentially result from a defect in the FppA-6
dependent processing of the Flp pilin (2), we investigated whether Flp pili assembled at the 7
surface of PprB-overproducing bacteria. The PAO1/pMMBpprB and PAO1/pMMB67-HE 8
strains did have similar numbers and lengths of type IVb pili at their surface, as shown by 9
TEM (Fig. 5B). As assembly of the Flp pilin subunit into a pilus requires prepilin peptidase-10
dependent processing (2), the assembly of Flp pili at the bacterial cell surface demonstrates 11
that Flp pilins are correctly processed in conditions of PprB overproduction and that EM shift 12
observed is not due to an FppA-dependent change in Flp maturation. 13
We therefore investigated the possible involvement of the rcp-tad genes in the 14
observed Flp EM shift. As the functions of RcpA, TadA, TadB, TadC, TadF and FppA have 15
been clearly defined, we investigated the role of the first gene of the rcpC-tadG operon, rcpC, 16
which encodes a putative protein of unknown function. Overproduction of the PprB response 17
regulator in the ∆rcpC background abolished the Flp EM shift (Fig. 6A) with respect to the 18
WT strain (Fig. 6A). This shift was fully restored by introducing the rcpC gene into the 19
chromosome at the attB sites (Fig. 6A). Thus, the RcpC putative protein is involved in the 20
modification of the Flp protein leading to the observed Flp EM shift. Further electron 21
microscopy studies on Flp pilus assembly showed that deletion of the rcpC gene had no effect 22
on Flp assembly at the surface of the bacteria (Fig. 6B). These results suggest that the rcpC 23
gene product may be involved in a modification that does not affect Flp stability and pilus 24
assembly. 25
26
The putative RcpC protein regulates Flp pilus-mediated attachment to eukaryotic cells 27
ACCEPTED
on May 25, 2018 by guest
http://jb.asm.org/
Dow
nloaded from

18
We previously showed that Flp overproduction led to biofilm formation in a P. aeruginosa 1
strain (PAO1∆pilA∆fliC) devoid of both type IVa pili and flagella (2). We investigated the 2
role of the putative RcpC protein in the Flp pilus-dependent biofilm phenotype, by creating an 3
rcpC mutation in the PAO1∆pilA∆fliC strain. Strains collected after culture in conditions of 4
optimal Flp production from the chromosomal gene and for which pili were already 5
assembled at the surface, were used for inoculation in static conditions, as described in the 6
Materials and Methods section. The PAO1∆pilA∆fliC parental strain formed clusters of cells 7
after 10 h (Fig. 6C). These clusters began to grow after 6 h (data not shown). At this time 8
point, the parental PAO1 strain had already formed a continuous layer with a three-9
dimensional structure. The flp mutant adhered only as sparse, isolated bacteria, confirming the 10
strong involvement of Flp pili in bacterium-surface attachment and in the bacterium-11
bacterium contact and aggregation observed in a previous study (2). The rcpC mutant 12
displayed similar bacterial cell clustering to the parental strain (Fig. 6C), suggesting that 13
RcpC-dependent Flp modification had no significant effect in these conditions. 14
We therefore investigated whether the rcpC gene product controlled the Flp-dependent 15
adhesion to respiratory epithelial cells observed in a previous study (2). In the absence of Flp 16
pili, P. aeruginosa cells adhered only weakly to eukaryotic cells. The rcpC mutant displayed a 17
significantly (p<0.001) lower (by a factor of 3) level of adhesion to eukaryotic cells than the 18
parental isogenic strain (Fig. 7 A and B). The parental phenotype was fully restored by 19
introducing the rcpC gene at the attB sites (Fig. 7B, p<0.0001). This finding suggests that the 20
putative RcpC protein influences Flp Type IVb pilus-host receptor interactions. 21
22
The putative TadF pseudopilin is not required for Flp production, type IVb pilus 23
assembly and adhesion 24
The TadF pseudopilin is essential for Flp pilus assembly in A. actinomycetemcomitans. This 25
gene is weakly expressed in P. aeruginosa (see Fig. 3F). The effect of tadF mutation was 26
tested for Flp production, type IVb pilus assembly and adhesion (Fig. S1). The tadF mutant 27
ACCEPTED
on May 25, 2018 by guest
http://jb.asm.org/
Dow
nloaded from

19
produced Flp protein in similar amounts to the PAO1 parental strain and the assembly of Flp 1
type IVb pili was also similar in the tadF and the WT strains. The introduction of a tadF 2
mutation into the PAO1∆pilA∆fliC strain did not affect biofilm formation on glass slides or 3
Flp-dependent adhesion to respiratory epithelial cells. These results demonstrate that, in the 4
conditions used, the P. aeruginosa TadF putative pseudopilin is dispensable for Flp pilus 5
biogenesis and Flp-dependent adhesion phenotypes. 6
ACCEPTED
on May 25, 2018 by guest
http://jb.asm.org/
Dow
nloaded from

20
Discussion 1
P. aeruginosa is the only bacterium known to be able to assemble both type IVa and type IVb 2
pili. Type IVb pili of the Flp subfamily consist of the major pilin, Flp, assembled through the 3
concerted action of a dedicated machine, the Tad machine. We optimised the culture 4
conditions allowing reproducible production of Flp. This made it possible (i) to dissect the 5
mechanism regulating the tad locus and (ii) to obtain insight into the function of putative 6
products encoded by several genes of this locus. By contrast to the results reported here for P. 7
aeruginosa, anaerobic growth conditions favour the production of Flp pili in A. 8
actinomycetemcomitans (28), whereas high CO2 levels ensure reproducible Flp pilus 9
production in A. aphrophilus (32). The P. aeruginosa PprA sensor of the PprAB TCS 10
possesses PAS domains, which are involved in sensing environmental signals, such as oxygen 11
concentration, redox potential or light. The environment-dependent modulation of type IVb 12
pilin production seems to be a common feature, having also been reported in A. aphrophilus, 13
A. actinomycetemcomitans (32) and Caulobacter crescentus (37). In C. crescentus, the 14
histidine kinase PleC must be present at the cell pole for spatial PilA (the pilin subunit) 15
accumulation and assembly (38). In C. crescentus, the global cell cycle regulator, CtrA, and 16
the GcrA master regulator control the concerted expression of different pilus genes over time 17
during the bacterial cell cycle (30, 14), after the initiation of replication and flagellum 18
biogenesis (22). 19
In silico analyses showed that the genetic organisation of the tad locus of P. 20
aeruginosa is more complex than that of other tad loci (20), suggesting major genomic 21
rearrangements or gene shuffling. The P. aeruginosa tad locus is also simpler than most of the 22
tad loci identified to date. It lacks the second flp gene, the rcpB gene and the second tadE 23
pseudopilin gene. However, functional Flp pili are assembled in the absence of these genes in 24
P. aeruginosa (no orthologues were identified in the P. aeruginosa genome, even outside the 25
tad locus). Unlike other tad loci, which are transcribed as a polycistronic mRNA (11), we 26
have shown that the P. aeruginosa tad locus is transcribed as five independent transcriptional 27
ACCEPTED
on May 25, 2018 by guest
http://jb.asm.org/
Dow
nloaded from

21
units (I: flp, II: rcpC-tadG, III: pprB, IV: fppA, V: tadF-pprA). The presence of the pprAB 1
genes, encoding a classical two component system (TCS), within the locus suggests that this 2
system is involved in regulating the tad cluster. Indeed, our results demonstrate that 3
transcriptional units I, II and IV are regulated by the PprB response regulator and that PprB 4
binds to the putative promoters of the five transcriptional units of the tad locus. The tadF-5
pprA operon (V) is weakly expressed, but its expression is stimulated by PprB 6
overproduction. As tadF is co-transcribed with pprA, the positive effect of the PprB response 7
regulator on the gene encoding its cognate sensor, PprA, creates a positive loop that may 8
contribute to the sequential and hierarchical regulation of a subset of genes, depending on the 9
affinity of the response regulator for different DNA-binding regions, as shown for the BvgS-10
BvgA TCS of Bordetella pertussis (5). The regulation of the transcriptional units IV and V 11
seems to be more complicated than that of transcriptional units I and II. The presence of 12
regulatory elements (two promoters, each with at least one PprB-binding site) on a short DNA 13
region between transcriptional units IV and V suggested that the PprB-dependent transcription 14
of one gene might influence the transcription of the other. In the absence of PprB, both these 15
genes are turned off. At low (physiological) PprB concentration, the fppA gene was expressed, 16
whereas the tadF gene was not. At high PprB concentration (pprB overexpression), the 17
activity of the fppA promoter was repressed, whereas the tadF promoter was switched on. 18
This hypothesis requires the existence of two independent PprB binding sites. Alternatively, 19
given the incomplete complementation of the pprB mutant for fppA-lacZ expression, it should 20
also be borne in mind that the pprB deletion generated may have had an unexpected polar 21
effect on the neighbouring genes, including fppA in particular, despite the removal of the full 22
nucleotide sequence of the gene, with the retention of only the start and stop codons, in our 23
deletion strategy. The partial complementation of the pprB mutant for the phenotypes tested 24
suggests that additional regulatory elements are lacking. Lastly, the binding of PprB to its 25
own promoter region is consistent with a model involving auto-regulation. 26
ACCEPTED
on May 25, 2018 by guest
http://jb.asm.org/
Dow
nloaded from

22
The PprA-PprB TCS was first identified as controlling cell permeability and 1
aminoglycoside sensitivity in the P. aeruginosa PAK strain (41). In this previous study, the 2
pprA mutant grew more slowly, had an altered membrane protein profile and produced more 3
OprF porin than the WT strain. However, our pprB deletion mutant grew as well as the WT 4
strain (data not shown), suggesting that the growth defect of the PAK pprA mutant may be 5
linked to a regulatory network involving PprA but not PprB, despite the demonstration of a 6
functional link between PprA and PprB (41). The overproduction of full-length PprA did not 7
affect flp expression, production or assembly (data not shown). This suggests that either PprB-8
dependent activation of the tad locus is independent of PprA or the overproduced full-length 9
PprA is not active. Alternatively, genetic organisation and regulation may differ between the 10
PAO1 and PAK strains. Differences in genetic organisation were ruled out by our observation 11
that the organisation of the tad locus was identical in PAK and PAO1 (data not shown). 12
Transcriptomic analyses of a PAO1 pprB mutant showed that this gene positively regulated 13
several operons, with levels of virulence factor secretion and of swimming or swarming 14
motility much lower in the mutant than in the WT strain (6). This result was recently 15
attributed to a second mutation in the LasR regulator in the pprB mutant used, rather to PprB 16
itself (7). In our hands, in-frame deletion of the pprB gene had no effect on proteolytic 17
activity, the secretion of extracellular compounds, twitching, swarming or swimming motility 18
(see supplementary Fig. S2). Our fusions were not affected in a pprA mutant (data not 19
shown), indicating that the functional relationships between PprA and PprB in control over 20
the tad locus may not be simple. 21
We also investigated the involvement of several other genes from this genetic locus in 22
type IVb pilus assembly or function. A gene encoding a pilin-like protein similar to the TadF 23
pseudopilin of A. actinomycetemcomitans was identified in the P. aeruginosa tad locus. By 24
contrast, the tad locus of A. actinomycetemcomitans contains two genes, tadE and tadF, 25
encoding putative pseudopilins. These two proteins are processed in a TadV-dependent manner 26
(31). By contrast to our results, a tadF mutant of A. actinomycetemcomitans was shown to 27
ACCEPTED
on May 25, 2018 by guest
http://jb.asm.org/
Dow
nloaded from

23
produce less Flp pilin (31), to be unable to assemble Flp pili (42, 18) and not to adhere to inert 1
surfaces (18). In our laboratory conditions, the putative TadF protein had no effect on P. 2
aeruginosa Flp biology. However, we cannot rule out the possibility of another gene in the P. 3
aeruginosa genome complementing TadF function or that P. aeruginosa tadF gene product plays 4
some role, as suggested by observations that the overproduction of PprB in a pprB mutant leads 5
to short Flp pili and induces the tadF-lacZ fusion. The putative TadF protein may control Flp 6
pilus length in P. aeruginosa, as previously reported for the pseudopilin XcpX controlling the 7
length of the pseudopilus assembled by the type II secretion machinery (8). 8
We then targeted the rcpC gene, because (i) this gene is present only in type IVb loci dedicated 9
to Flp pilus assembly and (ii) this gene has no known function (32). The rcpC gene product of P. 10
aeruginosa has a signal peptide (predicted by SignalP 3.0 software, data not shown), suggesting 11
that it may be located in the outer membrane or periplasm. The putative RcpC protein has been 12
localised to the A. actinomycetemcomitans outer membrane, although it is also detected in the 13
inner membrane (3). The additional protein of the machine, TadD, a putative lipoprotein (signal 14
peptidase (II) cleavage site between amino acids 16 and 17, as predicted by LipoP software, data 15
not shown), is probably located in the outer membrane. The putative RcpC protein, together with 16
the putative lipoprotein TadD and the secretin RcpA may therefore constitute an outer 17
membrane-complex unique to the Flp type IVb pilus machine in P. aeruginosa. Flp protein 18
stability and Flp pilus assembly were not altered in the rcpC mutant. However, we observed an 19
RcpC-dependent EM shift of the P. aeruginosa Flp pilin upon PprB overproduction. We 20
interpret this result as indicating only a small proportion of the thousands of copies of Flp 21
subunits assembled in a WT background is modified in an RcpC-dependent manner and that this 22
is not sufficient for detection of the change in electrophoretic mobility. Upon PprB 23
overproduction, levels of the putative RcpC protein — but not of Flp — increased, and the 24
modified Flp protein was thus the major form observed. This RcpC-dependent EM shift of Flp 25
pilin has previously been reported in A. actinomycetemcomitans (26). In P. aeruginosa, the rcpC 26
mutation had no effect on biofilm formation, but decreased adhesion to epithelial cells. These 27
ACCEPTED
on May 25, 2018 by guest
http://jb.asm.org/
Dow
nloaded from

24
findings suggest that the RcpC putative protein may be multifunctional, with independent roles 1
in Flp modification and adhesion. Furthermore, we cannot rule out the possible role of another 2
factor from epithelial cells during contact, rendering the Flp pili of rpcC bacteria more sensitive 3
to shearing. Alternatively, the putative RcpC protein may influence Flp pilin quality by possible 4
post-translational modification of the pilin subunit before its assembly��It has been suggested that 5
this phenomenon is due to incomplete or partial Flp pilin glycosylation in an A. 6
actinomycetemcomitans rcpC mutant (31). The RcpC-like proteins FlgA and CpaB belong to a 7
family of bacterial proteins containing two β-clip domains, probably located in the periplasm, 8
which are thought to bind sugar moieties (15). Interactions between the RcpC β-clip domains 9
and modified Flp pilin to facilitate extrusion through the secretin ring (32) are not consistent with 10
the assembly of the Flp pilus in an rcpC mutant, at least in P. aeruginosa. In this attractive 11
hypothesis, the “unmodified” Flp pilus in the rcpC mutant fails to bind to eukaryotic receptors, 12
whereas the modified version of this protein in the WT promotes adhesion to such receptors. 13
However, it remains unknown whether the adhesion phenotype of rcpC cells is linked to Flp 14
pilin modification. 15
Our results demonstrate that P. aeruginosa is able to produce type IVb Flp pili with a 16
simplified version of the Tad machine, the expression of which depends on the PprB response 17
regulator of the PprAB two-component system. 18 ACCEPTED
on May 25, 2018 by guest
http://jb.asm.org/
Dow
nloaded from

25
Acknowledgements 1
We would like to thank A. Bernadac and D. Byrne for technical assistance with the 2
microscopy platform and the protein production platform of the Institut de Microbiologie de 3
la Méditerrannée (Marseille, France), and M. Ansaldi’s team (G. Panis and L. Jacquet) from 4
UPR9043 (Marseille, France) for their advice on RT-PCR experiments. We thank Prof. 5
Shougouang Sin (University of Florida, Gainesville, USA) who kindly provided the E. coli 6
strain containing the plasmid with the His-tagged version of the PprB response regulator. We 7
are particularly grateful to Eric Cascales (UPR9027), for his invaluable scientific insight. The 8
work of SdB and AF is supported by the French Cystic Fibrosis Foundation (VLM), the 9
Bettencourt-Schueller Foundation and CNRS institutional grants. CSB is supported by the 10
French Cystic Fibrosis Foundation (VLM). AF is supported by the British Royal Society. 11
12
ACCEPTED
on May 25, 2018 by guest
http://jb.asm.org/
Dow
nloaded from

26
References 1
1. Becher, A., Schweizer, H.P. 2000. Integration-proficient Pseudomonas aeruginosa vectors 2
for isolation of single-copy chromosomal lacZ and lux gene fusions. Biotechniques. 29:948-3
50, 952. 4
5 2. de Bentzmann, S., Aurouze, M., Ball, G., and Filloux, A. 2006. FppA, a novel 6
Pseudomonas aeruginosa prepilin peptidase involved in assembly of type IVb pili. 7
J Bacteriol. 188: 4851-60. 8
9
3. Clock, S.A., Planet, P.J., Perez, B.A., and Figurski, D.H. 2008. Outer membrane 10
components of the Tad (tight adherence) secreton of Aggregatibacter actinomycetemcomitans 11
J Bacteriol. 190: 980-90. 12
13
4. Craig, L., Pique, M.E., and Tainer, J.A. 2004. Type IV pilus structure and bacterial 14
pathogenicity. Nat Rev Microbiol. 2: 363–378. 15
16
5. Deora, R., Bootsma, H.J., Miller, J.F., and Cotter, P.A. 2001. Diversity in the Bordetella 17
virulence regulon: transcriptional control of a Bvg-intermediate phase gene. Mol Microbiol. 18
40: 669-83. 19
20 6. Dong, Y.H., Zhang, X.F., Soo, H.M., Greenberg, E.P., and Zhang, L.H. 2005. The two-21
component response regulator PprB modulates quorum-sensing signal production and global 22
gene expression in Pseudomonas aeruginosa. Mol Microbiol. 56: 1287-301. 23
24
7. Dong, Y.H., Zhang, X.F., Soo, H.M., Greenberg, E.P., and Zhang, L.H. 2008. The two-25
component response regulator PprB modulates quorum-sensing signal production and global 26
gene expression in Pseudomonas aeruginosa. Mol Microbiol. 69: 780. 27
28
8. Durand, E., Michel, G., Voulhoux, R., Kürner, J., Bernadac, A., and Filloux, A. 2005. 29
XcpX controls biogenesis of the Pseudomonas aeruginosa XcpT-containing pseudopilus. J 30
Biol Chem. 280:31378-89. 31
32 9. Filloux, A., Bleves, S., van Ulsen, P., and Tommassen, J. 2004 Protein secretion 33
mechanisms in Pseudomonas. In Pseudomonas, Genomics, Life Style and Molecular 34
Architecture. 1: 749-91, edited by Juan-Luis Ramos. 35
36
10. Friedman, L., and Kolter, R. 2004. Two genetic loci produce distinct carbohydrate-rich 37
structural components of the Pseudomonas aeruginosa biofilm matrix. 38
J Bacteriol. 186: 4457-65. 39
40
11. Haase, E.M., Stream, J.O., and Scannapieco, F.A. 2003. Transcriptional analysis of the 41
5' terminus of the flp fimbrial gene cluster from Actinobacillus actinomycetemcomitans. 42
Microbiology. 149: 205-15. 43
44
12. Hoang, T.T., Karkhoff-Schweizer, R.R., Kutchma, A.J., and Schweizer, H.P. 1998. A 45
broad-host-range Flp-FRT recombination system for site-specific excision of chromosomally 46
located DNA sequences: application for isolation of unmarked Pseudomonas aeruginosa 47
mutants. Gene. 212: 77-86. 48
49
13. Hoang, T.T., Karkhoff-Schweizer, R.R., Kutchma, A.J., and Schweizer, H.P. 2000. 50
Integration proficient plasmids for Pseudomonas aeruginosa : site-specific integration and use 51
for engineering of reporter and expression strains. Plasmid. 43: 59-72. 52
ACCEPTED
on May 25, 2018 by guest
http://jb.asm.org/
Dow
nloaded from

27
14. Holtzendorff, J., Hung, D., Brende, P., Reisenauer, A., Viollier, P.H., McAdams, 1 H.H., and Shapiro, L. 2004. Oscillating global regulators control the genetic circuit driving a 2
bacterial cell cycle. Science. 304 : 983-7. 3
4
15. Iyer, L.M., and Aravind, L. 2004 The emergence of catalytic and structural diversity 5
within the beta-clip fold. Proteins. 55: 977-91. 6
7
16. Jackson, K.D., Starkey, M., Kremer, S., Parsek, M.R., and Wozniak, D.J. 2004 8
Identification of psl, a locus encoding a potential exopolysaccharide that is essential for 9
Pseudomonas aeruginosa PAO1 biofilm formation. J Bacteriol. 186: 4466-4475. 10
11
17. Juhas, M., Wiehlmann, L., Salunkhe, P., Lauber, J., Buer, J., and Tümmler, B. 2005. 12
GeneChip expression analysis of the VqsR regulon of Pseudomonas aeruginosa TB. FEMS 13
Microbiol Lett. 242: 287-95. 14
15
18. Kachlany, S.C., Planet, P.J., Bhattacharjee, M.K., Kollia, E., Desalle, R., Fine, D.H., 16 and Figurski, D.H. 2000. Nonspecific adherence by Actinobacillus actinomycetemcomitans 17
requires genes widespread in bacteria and archaea. J Bacteriol. 182: 6169-76. 18
19 19. Kachlany, S.C., Planet, P.J., Desalle, R., Fine, D.H., and Figurski, D.H. 2001. Genes 20
for tight adherence of Actinobacillus actinomycetemcomitans: from plaque to plague to pond 21
scum. Trends Microbiol. 9: 429-37. 22
23
20. Kachlany, S.C., Planet, P.J., Desalle, R., Fine, D.H., Figurski, D.H., and Kaplan, J.B. 24 2001. flp-1, the first representative of a new pilin gene subfamily, is required for non-specific 25
adherence of Actinobacillus actinomycetemcomitans. Mol Microbiol. 40: 542-54. 26
27
21. Kaniga, K., and Davison, J. 1991. Transposon vectors for stable chromosomal 28
integration of cloned genes in rhizosphere bacteria. Gene. 100:201-5. 29
30
22. Laub, M.T., McAdams, H.H., Feldblyum, T., Fraser C.M., and Shapiro, L. 2000. 31
Global analysis of the genetic network controlling a bacterial cell cycle Science. 290: 2144-8. 32
33
23. Matsukawa, M., and Greenberg, E.P. 2004. Putative exopolysaccharide synthesis genes 34
influence Pseudomonas aeruginosa biofilm development. J Bacteriol. 186: 4449-56. 35
36
24. Planet, P.J., Kachlany, S.C., Desalle, R., and Figurski, D.H. 2001. Phylogeny of genes 37
for secretion NTPases: identification of the widespread tadA subfamily and development of a 38
diagnostic key for gene classification. Proc Natl Acad Sci USA. 98: 2503-8. 39
40
25. Planet, P.J., Kachlany, S.C., Fine, D.H., Desalle, R., and Figurski, D.H. 2003 The 41
widespread colonization island of Actinobacillus actinomycetemcomitans. Nat Gene. 34: 193-42
8. 43
44
26. Perez, B.A., Planet, P.J., Kachlany, S.C., Tomich, M., Fine, D.H., and Figurski, D.H. 45 2006. Genetic analysis of the requirement for flp-2, tadV, and rcpB in Actinobacillus 46
actinomycetemcomitans biofilm formation. J Bacteriol. 188: 6361-75. 47
48
27. Ruer, S., Stender, S., Filloux, A., and de Bentzmann, S. 2007. Assembly of fimbrial 49
structures in Pseudomonas aeruginosa: functionality and specificity of chaperone-usher 50
machineries. J Bacteriol. 189: 3547-55. 51
52
ACCEPTED
on May 25, 2018 by guest
http://jb.asm.org/
Dow
nloaded from

28
28. Scannapieco, F.A., Millar, S.J., Reynolds, H.S., Zambon, J.J., and Levine, M.J. 1987. 1
Effect of anaerobiosis on the surface ultrastructure and surface proteins of Actinobacillus 2
actinomycetemcomitans (Haemophilus actinomycetemcomitans). Infect Immun. 55: 2320-3. 3
4
29. Schuster, M., Lostroh, C.P., Ogi, T., and Greenberg, E.P. 2003. Identification, timing, 5
and signal specificity of Pseudomonas aeruginosa quorum-controlled genes: a transcriptome 6
analysis. J Bacteriol. 185: 2066-79. 7
8
30. Skerker, J.M., and Shapiro, L. 2000. Identification and cell cycle control of a novel 9
pilus system in Caulobacter crescentus. EMBO J. 19: 3223-34. 10
11
31. Tomich, M., Fine, D.H., and Figurski, D.H. 2006. The TadV protein of Actinobacillus 12
actinomycetemcomitans is a novel aspartic acid prepilin peptidase required for maturation of 13
the Flp1 pilin and TadE and TadF pseudopilins. J Bacteriol. 188: 6899-914. 14
15
32. Tomich, M., Planet, P.J., and Figurski, D.H. 2007. The tad locus: postcards from the 16
widespread colonization island. Nat Rev Microbiol. 5: 363-75. 17
33. Vallet, I., Olson, J.W., Lory, S., Lazdunski, A., and Filloux, A. 2001. The 18
chaperone/usher pathway of Pseudomonas aeruginosa : identification of fimbrial gene clusters 19
(cup) and their involvement in biofilm formation. Proc Natl Acad Sci USA. 98: 6911-6. 20
21
34. Vallet, I., Diggle, S.P., Stacey, R.E., Cámara, M., Ventre, I., Lory, S., Lazdunski, A., 22 Williams, P., and Filloux, A. 2004. Biofilm formation in Pseudomonas aeruginosa: fimbrial 23
cup gene clusters are controlled by the transcriptional regulator MvaT. J Bacteriol. 186: 2880-24
90. 25
26
35. Vasseur, P., Vallet-Gely, I., Soscia, C., Genin, S., and Filloux, A. 2005. The pel genes 27
of the Pseudomonas aeruginosa PAK strain are involved at early and late stages of biofilm 28
formation. Microbiology. 151: 985-97. 29
30
36. Ventre, I., Filloux, A., and Lazdunski, A. 2004. Two-component signal transduction 31
systems: a key to the adaptative potential of Pseudomonas aeruginosa. In Pseudomonas, 32
Virulence and Gene Regulation 2: 257-88, edited by Juan-Luis Ramos. 33
34
37. Viollier, P.H., Sternheim, N., and Shapiro, L. 2002. A dynamically localized histidine 35
kinase controls the asymmetric distribution of polar pili proteins. EMBO J. 21: 4420-8. 36
37
38. Viollier, P.H., Sternheim, N., and Shapiro, L. 2002. Identification of a localization 38
factor for the polar positioning of bacterial structural and regulatory proteins. Proc Natl Acad 39
Sci USA. 99: 13831-6. 40
41
39. Wagner, V.E., Bushnell, D., Passador, L., Brooks, A.I., and Iglewski, B.H. 2003. 42
Microarray analysis of Pseudomonas aeruginosa quorum-sensing regulons: effects of growth 43
phase and environment. J Bacteriol. 185: 2080-95. 44
45
40. Wagner, V.E., Gillis, R.J., and Iglewski, B.H. 2004. Transcriptome analysis of quorum-46
sensing regulation and virulence factor expression in Pseudomonas aeruginosa. Vaccine. 22: 47
S15-20. 48
49
41. Wang, Y., Ha, U., Zeng, L., and Jin, S. 2003. Regulation of membrane permeability by 50
a two-component regulatory system in Pseudomonas aeruginosa. Antimicrob Agents 51
Chemother. 47: 95-101. 52
ACCEPTED
on May 25, 2018 by guest
http://jb.asm.org/
Dow
nloaded from

29
42. Wang, Y., and Chen, C. 2005. Mutation analysis of the flp operon in Actinobacillus 1
actinomycetemcomitans. Gene. 351: 61-71 2
3
ACCEPTED
on May 25, 2018 by guest
http://jb.asm.org/
Dow
nloaded from

30
Figure Legends 1
Fig. 1. A. Genetic organisation of the P. aeruginosa tad locus and locations of the primers 2
(see Table 2) used for genetic manipulations (indicated above the locus, in bold, for RT-PCR 3
manipulations, leading to amplification of the fragments (dotted arrows) circled from 1 to 12, 4
indicated under the locus for all other manipulations). Arrows indicate the five transcriptional 5
units identified (I-V). Known functions of tad genes are specified below. RR for response 6
regulator, HK for histidine kinase and TCS for two-component system. B. Schematic 7
representation of the domains identified by SMART prediction in the sensor PprA and in the 8
response regulator PprB. Lengths of peptide sequences are indicated on the left (aa: amino 9
acids). C. The polycistronic transcription of tad genes within the locus was studied using 10
primers (indicated below numbers) designed to amplify regions (numbered 1 to 12) spanning 11
gene junctions with genomic DNA (upper panel), RNA (middle panel) or cDNA (lower 12
panel) obtained by reverse transcription of extracted mRNA. The molecular weights of the 13
marker bands (bp for base pair) in each gel are indicated on the left. 14
15
Fig. 2. A. Expression of the chromosomal flp-lacZ fusion, monitored in the PAO1 strain. Data 16
are expressed in Miller units and correspond to the mean values (with error bars) obtained 17
from three independent experiments. The corresponding growth curve (dotted curve) for the 18
strain carrying the flp-lacZ chromosomal fusion is presented. B. Detection of Flp pilin 19
production in whole-cell extracts from the PAO1 strain at various time points (2, 4, 7, 12 and 20
24 hours referred to on the growth curve as points 1, 2, 3, 4 and 5, respectively) during growth 21
in L broth at 30oC, with vigorous shaking. Signal specificity was checked by Flp detection in 22
the PAO1∆flp strain collected at points 1 and 5 of growth. The proteins were separated by 23
electrophoresis in a 16.5% polyacrylamide Tris-glycine gel, transferred onto nitrocellulose 24
and detected with an antibody against Flp. The numbers on the right indicate the sizes of the 25
molecular weight standards (kDa). C. Flp type IVb pilus (arrow) on the surface of P. 26
aeruginosa cells following Flp production from a chromosomal gene (left panel), 27
ACCEPTED
on May 25, 2018 by guest
http://jb.asm.org/
Dow
nloaded from

31
magnification x 50,000, and absence of a labelled structure at the surface of the PAO1∆flp 1
strain (right panel), magnification x 50,000. Size markers correspond to 0.7µm. 2
3
Fig. 3. A-B. The expression of flp-lacZ (A) and rcpC-lacZ (B) chromosomal fusions was 4
monitored in the PAO1 strain containing pMMB67-HE (black circles) and pMMBpprB (open 5
circles). The corresponding growth curves (dotted curves) are presented. Numbers in brackets 6
refer to the corresponding transcriptional units. C. The level of expression of the flp-lacZ (I), 7
rcpC-lacZ (II), fppA-lacZ (IV) and tadF-lacZ (V) fusions was evaluated when cells reached 8
the early stationary phase, in the PAO1/pMMB67-HE, PAO1∆pprB/pMMB67-HE and 9
PAO1∆pprB/pMMBpprB strains. D. The levels of expression of the flp-lacZ (I), rcpC-lacZ 10
(II), fppA-lacZ (IV) and tadF-lacZ (V) fusions were monitored in MG1655 E. coli strain 11
transformed with pMMB67-HE or pMMBpprB and induced with IPTG. Data are expressed in 12
Miller units and correspond to the mean values (with error bars) obtained from three 13
independent experiments. 14
15
Fig. 4. A-C. Electrophoretic mobility shift assays performed with the purified PprB-6His 16
protein, at concentrations of 0 to 2.34 µM, and the intergenic DNA regions identified in the P. 17
aeruginosa tad locus that were amplified by PCR to generate DNA fragments of 501, 561 and 18
560 bp for the regions upstream from the flp/rcp (A), tadF/fppA (B), pprB (C) genes. Two 19
retarded complexes (**) and (*) were clearly identified at high PprB-6His concentrations for 20
the flp/rcp (A), tadF/fppA (B) intergenic regions, whereas only one retarded complex was 21
observed for the pprB intergenic region. No shift was observed for the 360 bp DNA fragment 22
corresponding to the DNA region upstream from the rsmZ gene. Molecular size markers were 23
run on each gel and are indicated on the right (bp for base pair). 24
25
Fig. 5. A. Flp production in the PAO1/pMMB67-HE (lane 1), PAO1/pMMBpprB (lane 2), 26
PAO1∆pprB/pMMB67-HE (lane 3) and PAO1∆pprB/pMMBpprB (lane 4) strains. Cells at an 27
ACCEPTED
on May 25, 2018 by guest
http://jb.asm.org/
Dow
nloaded from

32
OD600 of 0.5 were induced by incubation with 0.1 mM IPTG. The numbers on the right 1
indicate the sizes of the molecular weight standards (kDa). B. The assembly of Flp type IVb 2
pili (arrow) at the surface of the bacteria was checked by TEM coupled to immunogold 3
labelling, using the anti-Flp antibody in the PAO1/pMMB67-HE, PAO1/pMMBpprB, 4
PAO1∆pprB/pMMB67-HE and PAO1∆pprB/pMMBpprB strains, magnifications x 50,000. 5
The size bar indicates 0.7 µm. C. Lengths of Flp pili were quantified for 30 images for each 6
strain acquired systematically from at least three independent experiments in the wild-type 7
PAO1 and the pprB mutant trans-complemented with the pprB gene. Unpaired t-tests were 8
used for the comparison of results, ***: p<0.0001. 9
10
Fig. 6. A. Flp production in the PAO1/pMMBpprB (lane 1), PAO1∆rcpC/pMMBpprB (lane 11
2), PAO1∆rcpC attB::rcpC/pMMBpprB (lane 3), PAO1/pMMB67-HE (lane 4), and 12
PAO1∆rcpC/pMMB67-HE (lane 5) strains. Cells at an OD600 of 0.5 were induced by 13
incubation with 0.1 mM IPTG. The numbers on the right are the sizes of the molecular weight 14
standards (kDa). B. Assembly of Flp type IVb pili (arrow) at the bacterial cell surface in the 15
PAO1/pMMBpprB, PAO1∆rcpC/pMMB67-HE, PAO1∆rcpC/pMMBpprB and PAO1∆rcpC 16
attB::rcpC/pMMBpprB strains, magnification x 30,000 and x 40,000. The size bars 17
correspond to 1 µm and 0.5 µm, respectively. C. Biofilm formation at the air-liquid interface 18
of glass slides immersed in culture medium was analysed by confocal laser scanning 19
microscopy observation after DAPI staining. Stacked images and corresponding extracted z 20
images (z-slices of 200 nm) and their respective xy and xz planes at x 180 magnification were 21
obtained for the PAO1, PAO1∆pilA∆fliC, PAO1∆pilA∆fliC∆flp, and PAO1∆pilA∆fliC∆rcpC 22
strains. 23
24
Fig. 7. A. Representative views of adhesion to bronchial epithelial cells incubated with the 25
PAO1∆pilA∆fliC, PAO1∆pilA∆fliC∆flp, PAO1∆pilA∆fliC∆rcpC and PAO1∆pilA∆fliC∆rcpC 26
ACCEPTED
on May 25, 2018 by guest
http://jb.asm.org/
Dow
nloaded from

33
attB::rcpC strains. B. The number of bacteria adhering to bronchial epithelial cells was 1
evaluated in three independent assays Adherent bacteria were quantified for the various 2
genetic backgrounds (PAO1∆pilA∆fliC, PAO1∆pilA∆fliC∆flp, PAO1∆pilA∆fliC∆rcpC and 3
PAO1∆pilA∆fliC∆rcpC attB::rcpC strains), on epithelial cells (30 epithelial cells/assay), in 4
randomly chosen microscopic fields. Comparisons were made by one way-analysis of 5
variance and unpaired t-tests, ***: p<0.0001, **: p<0.001. 6
7
Supplementary Figures 8
Fig. S1. A. Flp production in the PAO1 (lane 1) and PAO1∆tadF (lane 2) strains. B. 9
Assembly of Flp type IVb pili (arrow) at the bacterial cell surface in the PAO1 and 10
PAO1∆tadF strains, magnifications x 50,000 and x 30,000, respectively. Size bars correspond 11
to 0.7 µm and 1.2 µm, respectively. C. Biofilm formation, assessed by confocal laser 12
scanning microscopy observation after DAPI staining. Extracted z images (z-slices of 200 13
nm) at x 180 magnification were obtained for the PAO1∆pilA∆fliC and 14
PAO1∆pilA∆fliC∆tadF strains. D. Adhesion to bronchial epithelial cells was assessed in the 15
PAO1∆pilA∆fliC and the PAO1∆pilA∆fliC∆tadF strains. 16
17
Fig. S2. Characterisation of the in-frame pprB deletion mutant constructed in this study. A. 18
Proteolytic activity of the PAO1 and ∆pprB mutant strains was analysed on TSA-milk agar 19
plates. The presence of a halo indicates proteolytic activity of the strains after overnight 20
incubation at 30°C. Supernatants of the PAO1 and PAO1∆pprB mutant strains were analyzed 21
by SDS-PAGE electrophoresis. B. Twitching motility was evaluated by picking a single 22
colony and stabbing it to the bottom of a 1.5% LB agar plate. After 48 h of incubation at 37°C, 23
the agar was removed, and the twitching zones were stained by incubation with crystal violet 24
for 5 minutes. No difference was observed between the PAO1 and PAO1∆pprB mutant 25
strains. Control: PAO1∆pilA mutant strain. C. Swarming motility was evaluated by depositing 26
ACCEPTED
on May 25, 2018 by guest
http://jb.asm.org/
Dow
nloaded from

35
Table 1: Strains and plasmids used in this study 1 2
*Sm
R, streptomycin resistance, Ap
R, ampicillin resistance, Km
R, kanamycin resistance, Tc
R 3
tetracycline resistance 4
Strains/Plasmids Relevant characteristics* Source
Strains
E. coli
TG1 supE ∆(lac-proAB) thi hsdR∆5 (F’ traD36 rpoA+ B
+ lacI
q Z∆M15) Lab collection
MG1655 K ∆lacZ pcnB::KnR Lab collection
TOP10F’ F’ (lacIq Tn10 (Tet
R)) mrcA ∆(mrr-hsdRMS-mcrBC) Φ80 lacZ∆M15
∆lacX74 recA1
INVITROGEN
CC118(λpir)
M15
Host strain for pKNG101 replication, ∆(ara-leu) araD ∆lacX74 galE
galK phoA20 thi-1 rpsE rpoB argE(Am) recA1 RfR (λpir)
lac ara gal mtl NalS Str
S Rif
S
Lab collection
(41)
P. aeruginosa
PAO1 Wild-type Lab collection
PAO1∆flp PAO1 deletion mutant for the flp gene (2)
PAO1∆pprB PAO1 deletion mutant for the pprB gene This study
PAO1∆tadF PAO1 deletion mutant for the tadF gene This study
PAO1∆rcpC
PAO1∆pilA∆fliC
PAO1∆pilA∆fliC∆flp
PAO1∆pilA∆fliC∆rcpC
PAO1∆pilA∆fliC∆tadF
PAO1∆rcpC attB::rcpC
PAO1∆pilA∆fliC∆rcpC
attB::rcpC
PAO1/flp-lacZ
PAO1/rcpC-lacZ
PAO1/fppA-lacZ
PAO1/tadF-lacZ
PAO1∆pprB/flp-lacZ
PAO1∆pprB/rcptad-lacZ
PAO1∆pprB/fppA-lacZ
PAO1∆pprB/tadF-lacZ
PAO1 deletion mutant for the rcpC gene
PAO1 deletion mutant for the pilA, fliC genes
PAO1 deletion mutant for the pilA, fliC, flp genes
PAO1 deletion mutant for the pilA, fliC, rcpC genes
PAO1 deletion mutant for the pilA, fliC, tadF genes
PAO1∆rcpC strain cis-complemented with wild-type rcpC gene inserted
at attB sites
PAO1∆pilA∆fliC∆rcpC strain cis-complemented with wild-type rcpC
gene inserted at attB sites
PAO1 strain with flp-lacZ fusion inserted at attB sites
PAO1 strain with rcpC-lacZ fusion inserted at attB sites
PAO1 strain with fppA-lacZ fusion inserted at attB sites
PAO1 strain with tadF-lacZ fusion inserted at attB sites
PAO1∆pprB strain with flp-lacZ fusion inserted at attB sites
PAO1∆pprB strain with rcpC-lacZ fusion inserted at attB sites
PAO1∆pprB strain with fppA-lacZ fusion inserted at attB sites
PAO1∆pprB strain with tadF-lacZ fusion inserted at attB sites
This study
(2)
This study
This study
This study
This study
This study
This study
This study
This study
This study
This study
This study
This study
This study
This study
Plasmids
pCR2.1 TA cloning vector for PCR products, lacZα ColE1 f1 ori, ApR ; Km
R INVITROGEN
pMMB67-HE Broad host range vector, IncQ; ptac; lacZα; ApR Lab collection
pEX18 ApR; oriT
+; sacB
+, gene replacement vector with MCS from pUC18 Lab collection
miniCTX-lacZ TcR; lacZ
+ ; self-proficient integration vector with tet, V-FRT-attPMCS,
ori; int; and oriT (13)
pMMBpprB
pYW024
pprB gene cloned in pMMB67-HE, ApR
pprB gene cloned in pQE31 vector with a His-tag, ApR
This study
(41)
miniCTX-pflp-lacZ Promoter region of flp gene cloned in miniCTX-lacZ at KpnI/XhoI sites This study
miniCTX-prcpC-lacZ
miniCTX-pfppA-lacZ
miniCTX-ptadF-lacZ
miniCTX-rcpC
Promoter region of rcpC locus cloned in miniCTX-lacZ at XhoI/HindIII
sites
Promoter region of fppA gene cloned into miniCTX-lacZ at XhoI/BamHI
sites
Promoter region of tadF locus cloned into miniCTX-lacZ at XhoI/KpnI
sites
rcpC gene and its promoter region cloned into miniCTX-lacZ at
XhoI/HindIII sites
This study
This study
This study
This study
pEX18-DelpprB pprB gene mutator cloned into pEX18, ApR; sacB
+ This study
pEX18-DelrcpC rcpC gene mutator cloned into pEX18, ApR; sacB
+ This study
pEX18-DeltadF tadF gene mutator cloned into pEX18, ApR; sacB
+ This study
pRK2013 ColE1 ori; tra+; mob
+; Km
R Lab collection
ACCEPTED
on May 25, 2018 by guest
http://jb.asm.org/
Dow
nloaded from

36
Table 2 . Oligonucleotides used for mutation engineering and gene cloning 1
Oligonucleotides (5’→3’)
Mutations
pprB
1. DelPBUp5 TATATAGAGCTCCCGTCTGGCTGAAAAGCATGGCC
2. DelPBUp3 TATATAGGTACCTGACAGGCGCGATGGCCCGG
3. DelPBDn5 TATATAGGTACCCATGGTCATACGCTCCATTTG
4. DelPBDn3 TATATAGGATCCAACGTTTGCGCGTCTCCAAC
rcpC 5. DelRCUp5 TATATAGAGCTCTTCTTCGTCCGCCAGGAATGC
6. DelRCUp3 TATATAGGTACCCATGGGCAGACCCCTGCTATG
7. DelRCDn5 TATATAGGTACCGGCAGCGCCATGACCCATGA
8. DelRCDn3
tadF
9. DelTFUp5
10. DelTFUp3
11. DelTFDn5
12. DelTFDn3
TATATAGGATCCACGAACGAGGTGCTGGCCTG
TATATAGAGCTCCCATGGCGAATATAAAAGGGAATG
TATATAGGTACCCATGAGCTGACATCCCCGATC
TATATAGGTACCCAGACGCTGAAGGCGAGTGCC
TATATAGGATCCATCTCCTGGACCGCCTCCGC
lacZ-fusions
13. PromFlRUp CACCGCACCGAGCAGCAGCAG
14. PromFlRDn
15. PromFpTUp
16. PromFpTDn
TTCTTCGTCCGCCAGGAATGC
ACAAGGCCAGTGCTAATTGC
CGGAAAGCTTTTCCGAGCAT
Complementations
pprB 17. PprBUp TAAGCAAATGGAGCGTATGACC
18. PprBDn GCGCCTGTCAGTGCACCAC
rcpC
RcpCUp or 14. PromFlRDn TTCTTCGTCCGCCAGGAATGC
19. RcpCDn
Operon structure 1: rcpC-rcpA genes
2: rcpA-tadZ genes
3: tadZ-tadA genes
4: tadA-tadB genes
5: tadB-tadC genes
6: tadC-tadD genes
7: tadD-PA4298 genes
8: PA4298-tadG genes
9: tadG gene
10: tadG-pprB genes
11: pprB-fppA genes
12: tadF-pprA genes
PromFpTlUp
PromFpTlDn
PromrsmZUp
PromrsmZDn
20. 4305-04U
21. 4305-04R
22. 4304-03U
23. 4304-03R
24. 4303-02U
25. 4303-02R
26. 4302-01U
27. 4302-01R
28. 4301-00U
29. 4301-00R
30. 4300-4299U
31. 4300-4299R
32. 4299-98U
33. 4299-98R
34. 4298-97U
35. 4298-97R
36. 4297-96U
37. 4297-96R1
38. 4297-96R2
39. 4296-95U
40. 4296-95R
41. 4294-93U
42. 4294-93R
CCGGTGCTCCGATGCATATG
AGCAAGGACGAAGAACTCTACC
AAGCCGCGACGATCCAGCA
TCGACAAGTTTCCCTGGCTG
CCGATACCAATCCCTCGAC
GAGCGCTACCTGCCCAATGT
AGTTGATGGCGCGAGCCGTA
CTGATGATTTGCGCCGCGCT
CCAGTGACTGCTGCGCGG
GCTCATCCACGAGCGCGA
ACCGAGATCAGGTACAGGGT
CTGCGGCGTGCTACGGCA
GATCAGCGCCCTTTCCTCG
AGCTGGATGCCCGGGACAT
TAGCGCTGGTAGGCACGCT
AGCGTGCCTACCAGCGCTA
CGCGCTGGCTATGAAACTGC
GAAAGCCGACGGTTCCGATC
CTAGATCAGCAGTTCCGCCC
CACCCAGTTGGCGCTGGC
GGTTTCTGGTAGTAGTCGGC
GCCTTCGAAGGGCACCGTCA
CCAGGGACGAGTTGCTCGCA
CTGCATCGGCTGGAAACGAC
GGCTGAGGCAGACGCCTATAA
GCCGGCATCCAGTCCAGG
CCGGAATTCCCTTAGACCCACTGAAGACC
GGGGTACCATCCTTCGGGGTTGCGTGTTCC
Nucleotides corresponding to restriction sites are underlined. 2
ACCEPTED
on May 25, 2018 by guest
http://jb.asm.org/
Dow
nloaded from

flp
rcpC
rcpA ta
dZ
tadA
tadB
tadC
tadD
tadG
pprB
pprA
tadFfp
pA
PA
4298
Figure 1: Bernard et al.
A
B
C1
20-21
2
22-23
3
24-25
4
26-27
5
28-29
6
30-31
7
32-33
8
34-35
9
36-37
10
36-38
11
39-40
12
41-42M
DNA
RNA
cDNA
GAF PAS PAS H
PAC PAC PAC
HTH
LuxRRECPprB (RR)
275 aa
PprA (HK)
922 aa
Oligonucleotide
pairs used1,000
500
300
bp
1,000
500
300
1,000
500
300
I II IV VIII
4306
4305
4304
4303
4302
4301
4300
4299
4297
4296
4295
4294
4293
pilin secretin ATPase PilC-like
proteins
prepilin
peptidase
pseudo-
pilin
TCS
RR
TCS
HK
ACCEPTED
on May 25, 2018 by guest
http://jb.asm.org/
Dow
nloaded from
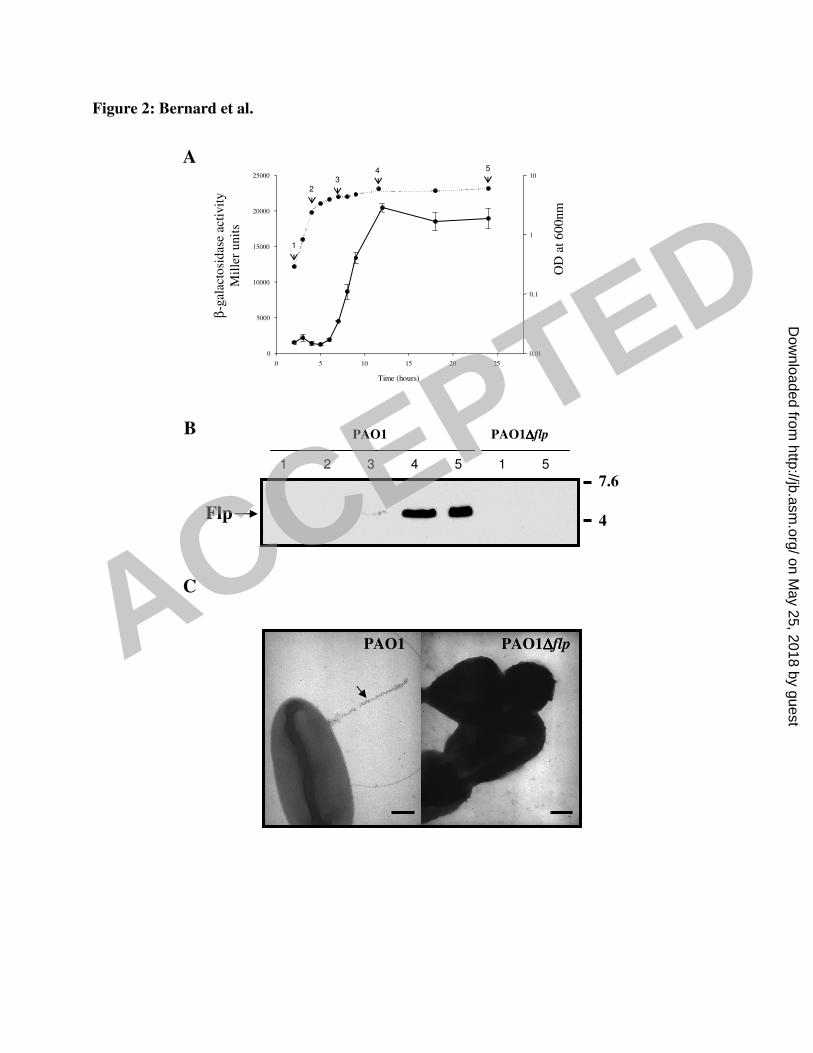
Flp
B
C
PAO1 PAO1∆∆∆∆flp
A
Time (hours)
0 5 10 15 20 25
0
5000
10000
15000
20000
25000
OD
at
60
0nm
0,01
0,1
1
10
β-g
alac
tosi
das
e ac
tivit
y
Mil
ler
unit
s1
32
5
Figure 2: Bernard et al.
4
7.6
PAO1 PAO1∆∆∆∆flp
1 2 3 4 5 51
4
ACCEPTED
on May 25, 2018 by guest
http://jb.asm.org/
Dow
nloaded from

A B
flp-lacZ (I) rcpC-lacZ (II)
Time (hours)
0 5 10 15 20 25
0
5000
10000
15000
20000
25000
30000
OD
at
600nm
0,01
0,1
1
10
Time (hours)
0 5 10 15 20 25
0
2000
4000
6000
8000
0,01
0,1
1
10
OD
at
600nm
β-g
alac
tosi
das
e ac
tivit
y
Mil
ler
unit
s
Figure 3: Bernard et al.
C
I II IV
D
ß-g
alac
tosi
das
eac
tivit
y
Mille
r unit
s
0
500
1000
1500
5000
10000
15000
20000
25000 PAO1 pMMB67-HE
PAO1∆pprB pMMB67-HE
PAO1∆pprB pMMBpprB
V0
5000
15000
20000
25000
ß-g
alac
tosi
das
eac
tivit
y
Mille
r unit
s
pMMB67-HE
pMMBpprB
I II IV V
ACCEPTED
on May 25, 2018 by guest
http://jb.asm.org/
Dow
nloaded from

Figure 4: Bernard et al.
PprB-6His (µM)
rsmZ
rsmZ
rsmZ
flp/rcp
tadF/fppA
pprB
500 bp
500 bp
500 bp
A
***
***
B
C
0 2.34
0.07
0.15
0.29
0.58
1.17
1000 bp
1000 bp
1000 bp
3000 bp
3000 bp
3000 bp
ACCEPTED
on May 25, 2018 by guest
http://jb.asm.org/
Dow
nloaded from
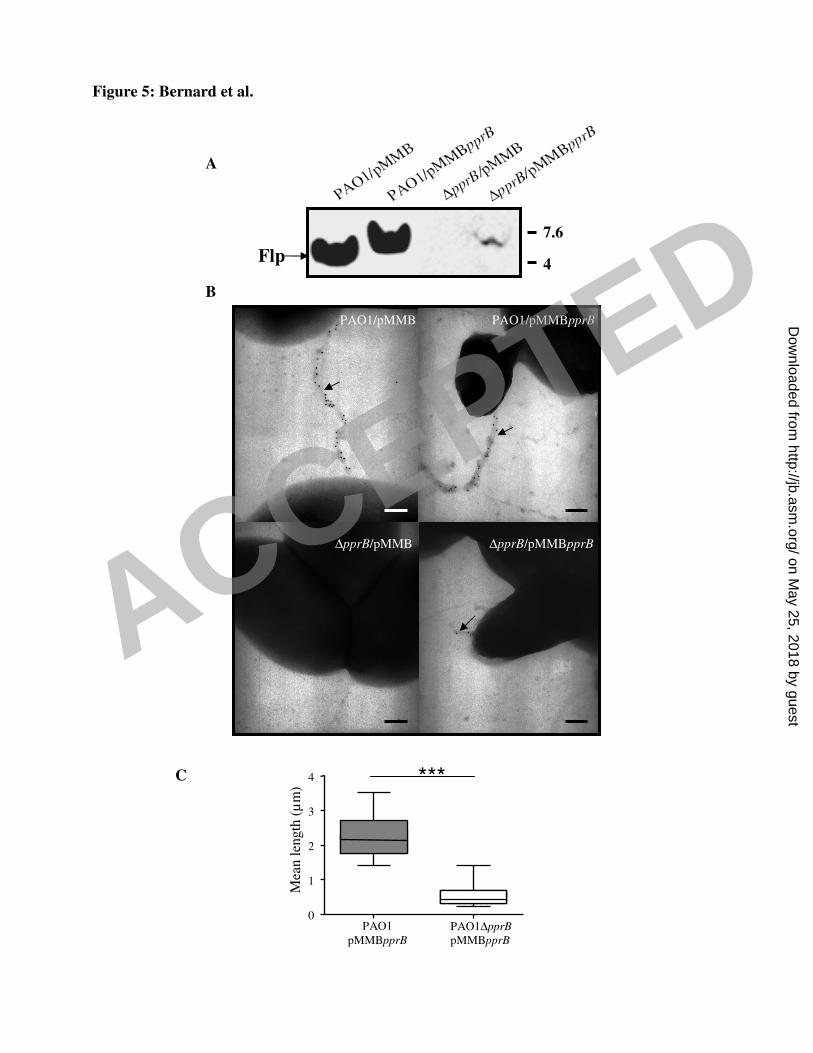
Figure 5: Bernard et al.
PAO1
pMMBpprB
PAO1∆pprB
pMMBpprB
0
1
2
3
4
Mea
nle
ngth
(µm
)
***
A
B
PAO1/pMMB PAO1/pMMBpprB
∆pprB/pMMB ∆pprB/pMMBpprB
Flp
C
4
7.6
ACCEPTED
on May 25, 2018 by guest
http://jb.asm.org/
Dow
nloaded from

A
B
C
PAO1 ∆pilA∆fliC ∆pilA∆fliC∆flp ∆pilA∆fliC∆rcpC
4
7.6
Flp
Figure 6: Bernard et al.
∆rcpC/pMMBpprB
∆rcpC attB::rcpC/pMMBpprB
PAO1/pMMBpprB
∆rcpC/pMMB67-HE
ACCEPTED
on May 25, 2018 by guest
http://jb.asm.org/
Dow
nloaded from

∆pilA∆fliC ∆pilA∆fliC∆flp ∆pilA∆fliC∆rcpC ∆pilA∆fliC∆rcpC
attB::rcpC
Figure 7: Bernard et al.
A
B
∆pilA∆fliC ∆pilA∆fliC
∆flp
0
25
50
75
100
Num
ber
of
bac
teri
a/
epit
hel
ial
cell
*****
***
∆pilA∆fliC
∆rcpC
∆pilA∆fliC
∆rcpC attB::rcpC
ACCEPTED
on May 25, 2018 by guest
http://jb.asm.org/
Dow
nloaded from


















