#2 Lecture2
26
Chapter 9 Chromosomes
-
Upload
kyle-broflovski -
Category
Documents
-
view
42 -
download
1
Transcript of #2 Lecture2

Chapter 9
Chromosomes

9.1. Introduction
Cellular genetic material (genome): DNA or RNA • They must be condensed to be packaged into phages, viruses,
bacteria, or eukaryotic nuclei. • Genome condensation results from its binding to basic proteins. • Bacterial genome is present as nucleoid. • Eukaryotic genome is present as chromatin at interphase or
chromosome during mitosis.
Figure 9.1
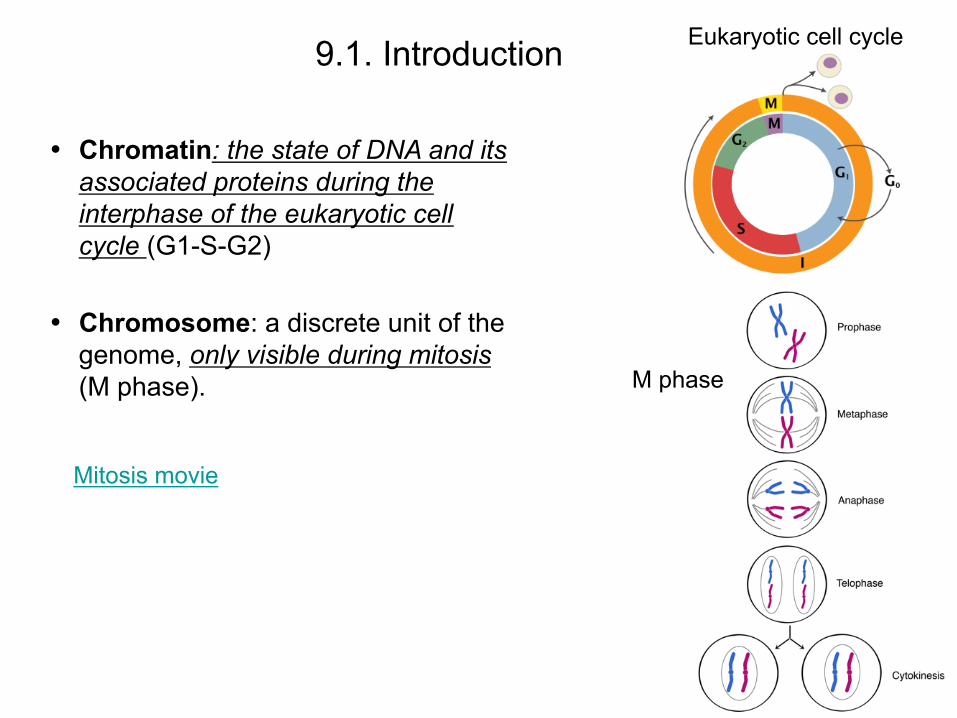
• Chromatin: the state of DNA and its associated proteins during the interphase of the eukaryotic cell cycle (G1-S-G2)
• Chromosome: a discrete unit of the genome, only visible during mitosis (M phase).
Eukaryotic cell cycle 9.1. Introduction
Mitosis movie
M phase
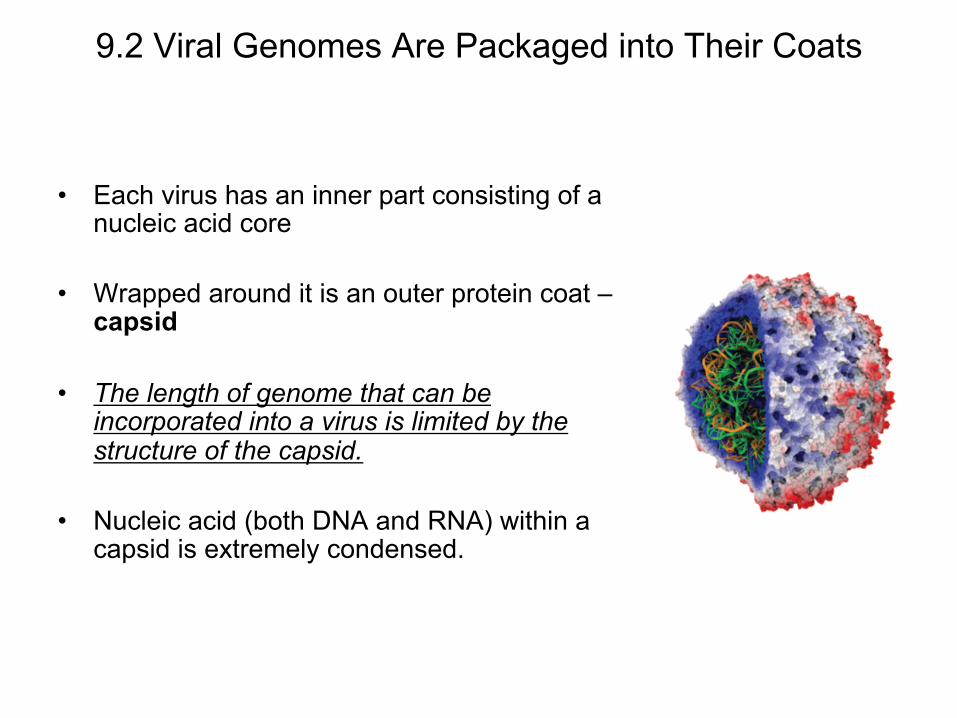
9.2 Viral Genomes Are Packaged into Their Coats
• Each virus has an inner part consisting of a nucleic acid core
• Wrapped around it is an outer protein coat – capsid
• The length of genome that can be incorporated into a virus is limited by the structure of the capsid.
• Nucleic acid (both DNA and RNA) within a capsid is extremely condensed.
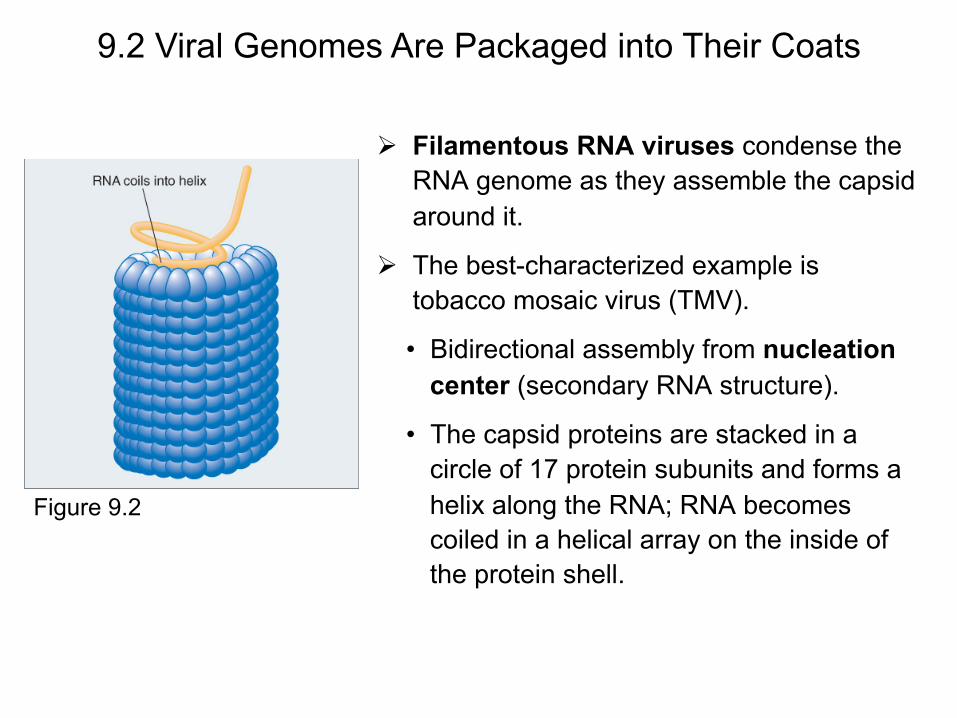
Filamentous RNA viruses condense the RNA genome as they assemble the capsid around it.
The best-characterized example is tobacco mosaic virus (TMV).
• Bidirectional assembly from nucleation center (secondary RNA structure).
• The capsid proteins are stacked in a circle of 17 protein subunits and forms a helix along the RNA; RNA becomes coiled in a helical array on the inside of the protein shell.
9.2 Viral Genomes Are Packaged into Their Coats
Figure 9.2

Spherical DNA viruses (e.g., Phage lambda and T4)
Phage lambda assembly • Empty protein core (prohead I) is made. • Core changes its shape to prohead II. • DNA packaging begins and head
expands while maintaining its shape. • The headshell reaches full size and
sealed by the addition of the tail.
9.2 Viral Genomes Are Packaged into Their Coats
Figure 9.3

Lambda DNA packaging involves two types of reaction: translocation and condensation.
• Translocation – terminase enzyme cleaves multimeric λ DNA into a monomer and translocates the DNA into an empty capsid using ATP hydrolysis (energy).
• Condensation - DNA is condensed as it enters the head. Its mechanism is unknown but it is not sequence-specific.
9.2 Viral Genomes Are Packaged into Their Coats
Figure 9.4
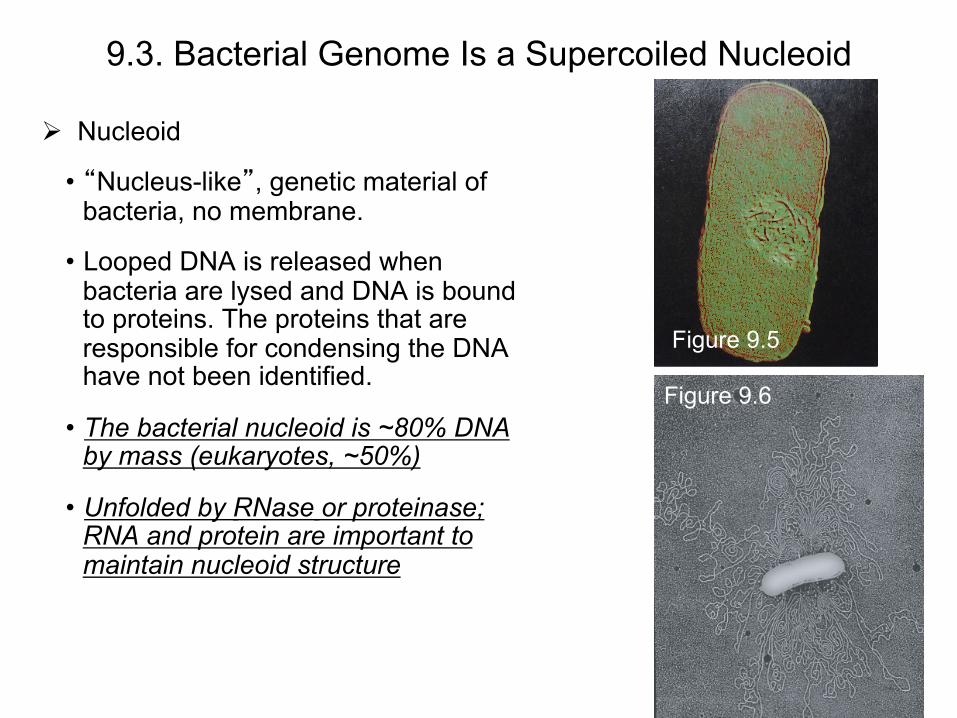
9.3. Bacterial Genome Is a Supercoiled Nucleoid
Nucleoid
• “Nucleus-like”, genetic material of bacteria, no membrane.
• Looped DNA is released when bacteria are lysed and DNA is bound to proteins. The proteins that are responsible for condensing the DNA have not been identified.
• The bacterial nucleoid is ~80% DNA by mass (eukaryotes, ~50%)
• Unfolded by RNase or proteinase; RNA and protein are important to maintain nucleoid structure
Figure 9.5
Figure 9.6
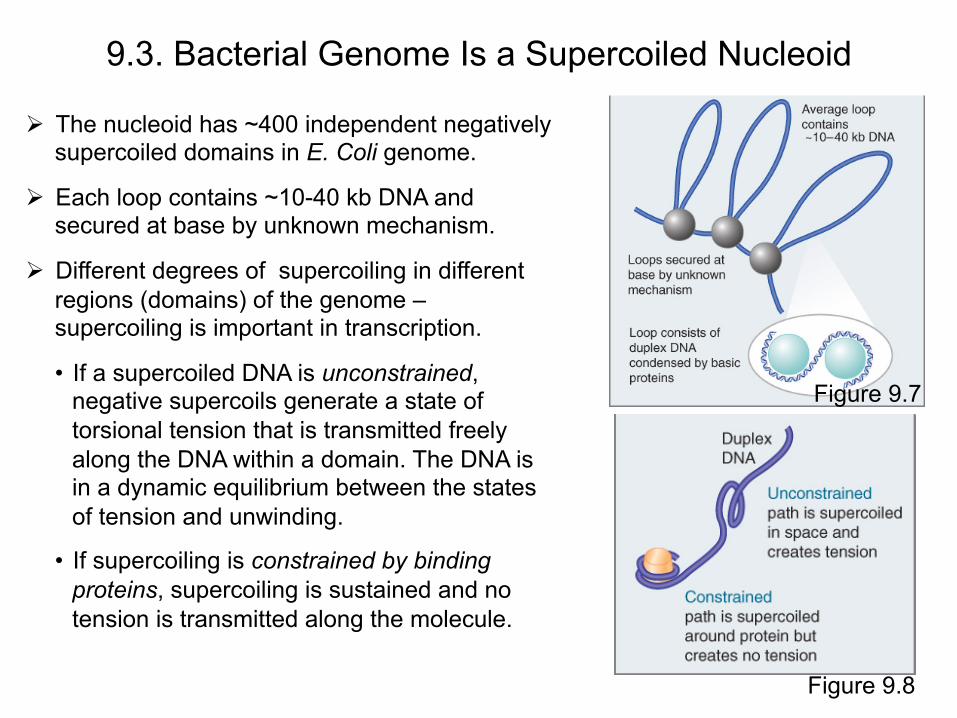
The nucleoid has ~400 independent negatively supercoiled domains in E. Coli genome.
Each loop contains ~10-40 kb DNA and secured at base by unknown mechanism.
Different degrees of supercoiling in different regions (domains) of the genome – supercoiling is important in transcription.
• If a supercoiled DNA is unconstrained, negative supercoils generate a state of torsional tension that is transmitted freely along the DNA within a domain. The DNA is in a dynamic equilibrium between the states of tension and unwinding.
• If supercoiling is constrained by binding proteins, supercoiling is sustained and no tension is transmitted along the molecule.
9.3. Bacterial Genome Is a Supercoiled Nucleoid
Figure 9.7
Figure 9.8

9.4. Eukaryotic DNA Has Loops and Domains Attached to a Scaffold
• Eukaryotic DNA has domains (~85 kb/domain)
• Loops can be seen when the majority of proteins (i.e., histones) are extracted from metaphase chromosomes.
• Metaphase scaffold contains ~8% of the original protein content and resembles the general form of sister chromatids.
• In interphase nuclei, this proteinaceous structure changes its organization to occupy the entire nucleus - matrix rather than scaffold
• Specific DNA regions are attached to the nuclear matrix or scaffold: matrix attachment regions (MARs) or scaffold attachment regions (SARs).
• The MARs are A-T-rich (~70%) but do not have any specific consensus sequence.
Figure 9.9
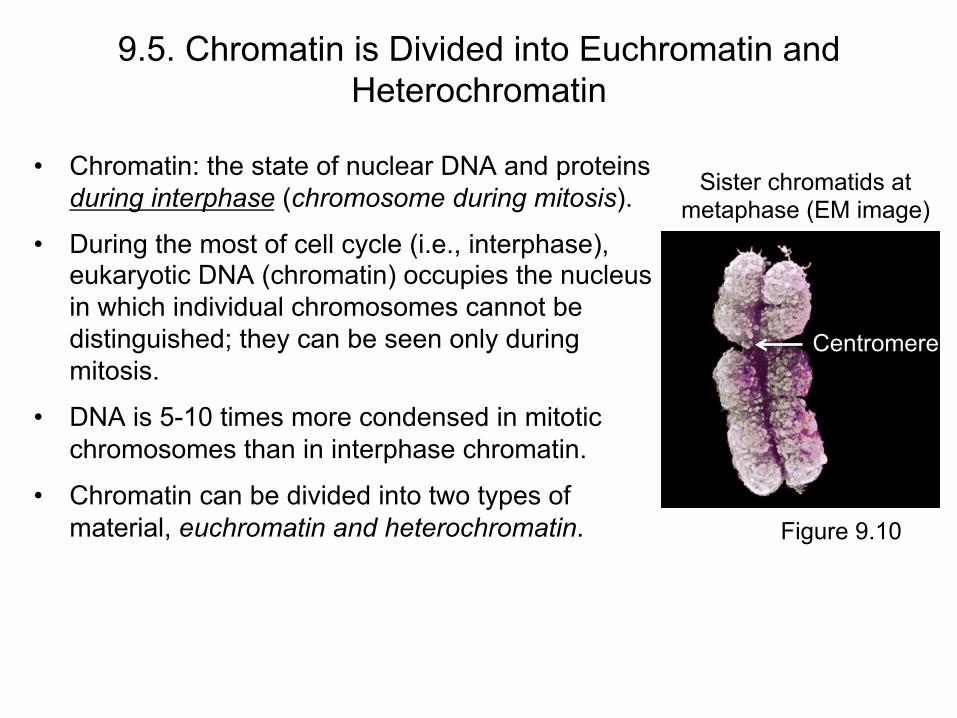
9.5. Chromatin is Divided into Euchromatin and Heterochromatin
• Chromatin: the state of nuclear DNA and proteins during interphase (chromosome during mitosis).
• During the most of cell cycle (i.e., interphase), eukaryotic DNA (chromatin) occupies the nucleus in which individual chromosomes cannot be distinguished; they can be seen only during mitosis.
• DNA is 5-10 times more condensed in mitotic chromosomes than in interphase chromatin.
• Chromatin can be divided into two types of material, euchromatin and heterochromatin.
Sister chromatids at metaphase (EM image)
Figure 9.10
Centromere

Euchromatin and Heterochromatin
• During interphase, most of the nuclear region is occupied by euchromatin, which is (1) less tightly packed than mitotic chromosomes, (2) stains lightly with DNA-specific dyes, and (3) contains transcriptionally active genes.
• Regions of heterochromatin remain densely packed throughout interphase. They strongly stains with DNA dyes and are as dense as chromosome.
9.5. Chromatin is Divided into Euchromatin and Heterochromatin
Figure 9.11
− Found at several locations within the chromatin, including the centromeres. − Chromocenter: aggregate of various heterochromatic regions − Constitutive chromatin: regions that are always heterochromatic. − Facultative chromatin: euchromatins converted to a heterochromatic state

Features of Constitutive Heterochromatin
• Permanently condensed • Multiple repeats of a few sequences of DNA that are not
transcribed or transcribed at very low levels • Replicates late in S phase and reduced frequency of
recombination • Reduced density of genes compared with euchromatin • Genes translocate into or near it are often inactivated (ribosomal DNA is exception; heterochromatic but
transcriptionally active)
9.5. Chromatin is Divided into Euchromatin and Heterochromatin

9.6. Chromosomes Have Banding Patterns • Certain staining techniques (e.g., Giemsa)
cause the chromosomes to have the appearance of a series of striations, which are called G-bands.
• Mechanism – unknown
• Each band is ~107 bp.
• The bands are lower in G-C content than the interbands.
• Genes are concentrated in interbands.
• Each chromosome has a unique pattern. - Identify chromosomes, translocation
between chromosomes.
Figure 9.12
Figure 9.13
• A gene location is indicated by its position on the long (q) or short (p) arm, region of the arm, band, and subband. − CFTR is located at 7q31.2.
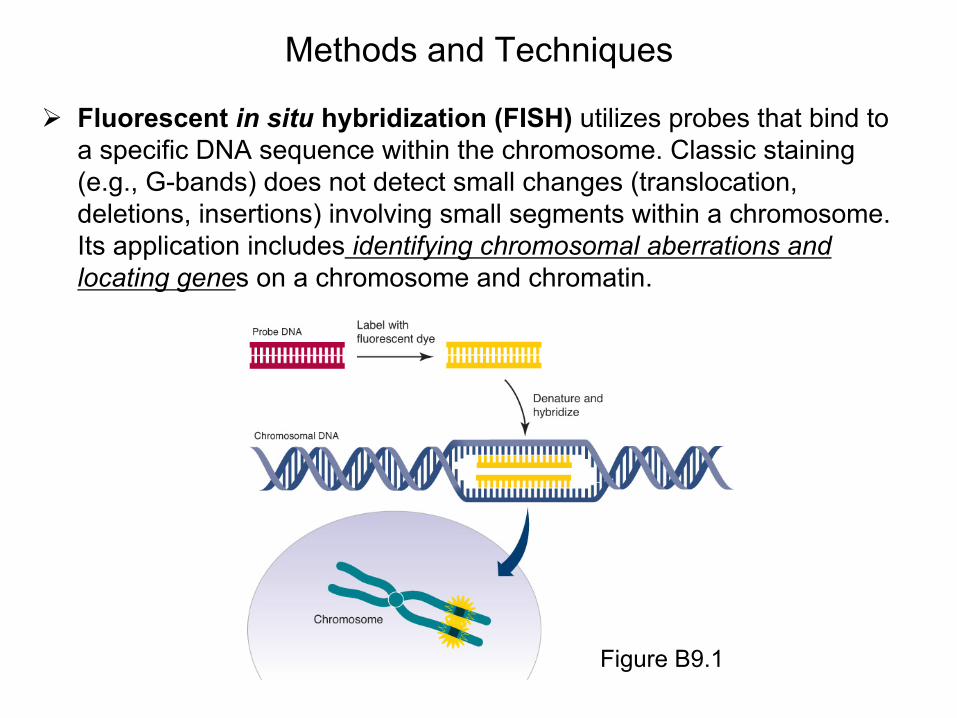
Methods and Techniques
Fluorescent in situ hybridization (FISH) utilizes probes that bind to a specific DNA sequence within the chromosome. Classic staining (e.g., G-bands) does not detect small changes (translocation, deletions, insertions) involving small segments within a chromosome. Its application includes identifying chromosomal aberrations and locating genes on a chromosome and chromatin.
Figure B9.1

Methods and Techniques
Improvement of FISH for more robust analysis
• Chromosome painting: use multiple fluorescent probes binding all along a particular chromosome (visualize one chromosome), unable to see all 23 chromosomes in a unique color because there are not enough fluorescent dyes with sufficient color differences to mark all 23 chromosomes in a unique color.
• Spectral karyotyping (SKY): simultaneously visualize all pairs of chromosomes in different colors. Chromosome-specific probes are labeled with different assortment of colors. Analyzed and presented with pseudo colors to each different color combination.
Figure B9.2

• Polytene chromosomes are found in the interphase nuclei of specific tissues of the larvae of dipteran (two-winged) insects such as Drosophila melanogaster. They possess increased diameter and greater length compared to their usual condition.
• Polytene chromosomes are generated by successive replications of a chromosome without separation of sister chromatids (endoreduplication): can duplicate up to a DNA content x1024 of a individual chromosome.
• They are composed of bands and interbands (note that bands and interbands here are different from G-bands).
• Bands contain most of the mass of DNA and are intensely stained; but interbands are lightly stained
9.7. Polytene Chromosomes Form Bands That Expand at Sites of Gene Expression
Figure 9.14
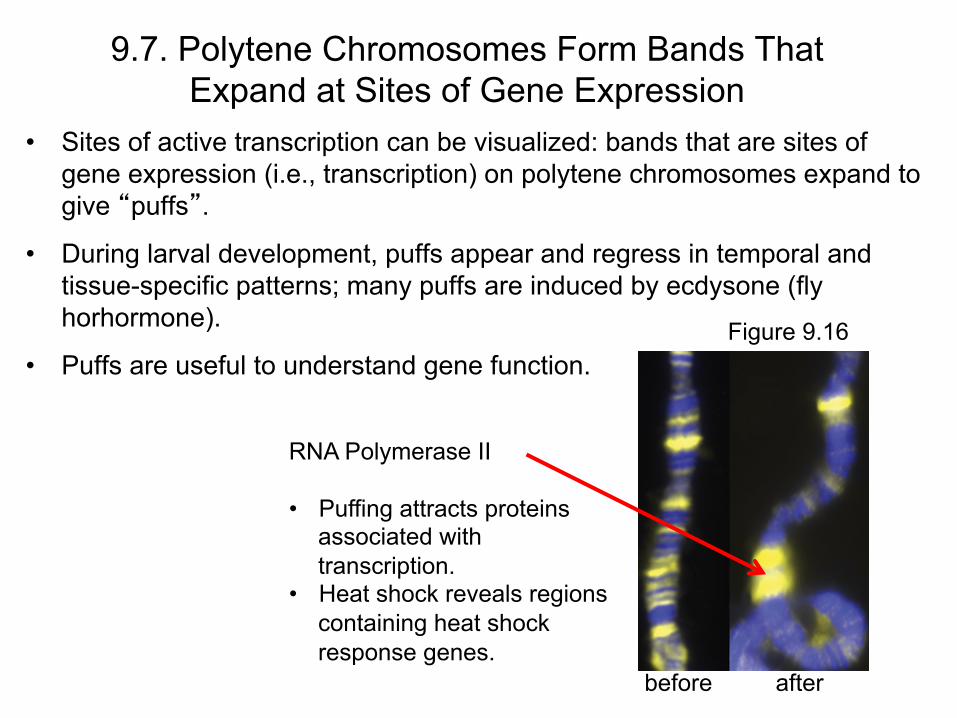
• Sites of active transcription can be visualized: bands that are sites of gene expression (i.e., transcription) on polytene chromosomes expand to give “puffs”.
• During larval development, puffs appear and regress in temporal and tissue-specific patterns; many puffs are induced by ecdysone (fly horhormone).
• Puffs are useful to understand gene function.
RNA Polymerase II • Puffing attracts proteins
associated with transcription.
• Heat shock reveals regions containing heat shock response genes.
before
9.7. Polytene Chromosomes Form Bands That Expand at Sites of Gene Expression
Figure 9.16
after
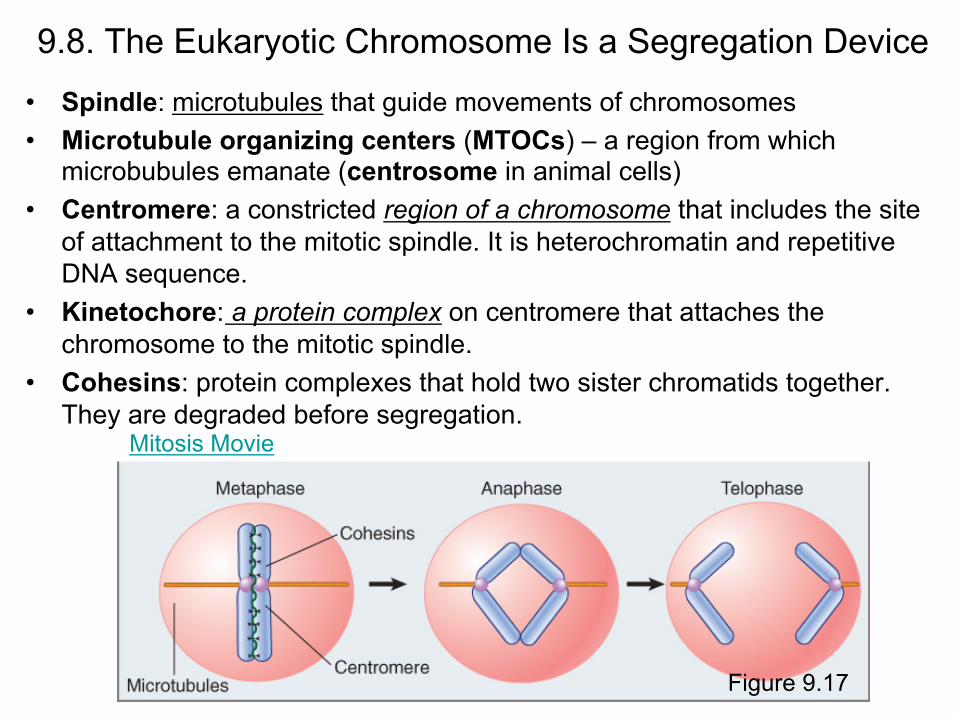
9.8. The Eukaryotic Chromosome Is a Segregation Device
• Spindle: microtubules that guide movements of chromosomes • Microtubule organizing centers (MTOCs) – a region from which
microbubules emanate (centrosome in animal cells) • Centromere: a constricted region of a chromosome that includes the site
of attachment to the mitotic spindle. It is heterochromatin and repetitive DNA sequence.
• Kinetochore: a protein complex on centromere that attaches the chromosome to the mitotic spindle.
• Cohesins: protein complexes that hold two sister chromatids together. They are degraded before segregation.
Figure 9.17
Mitosis Movie
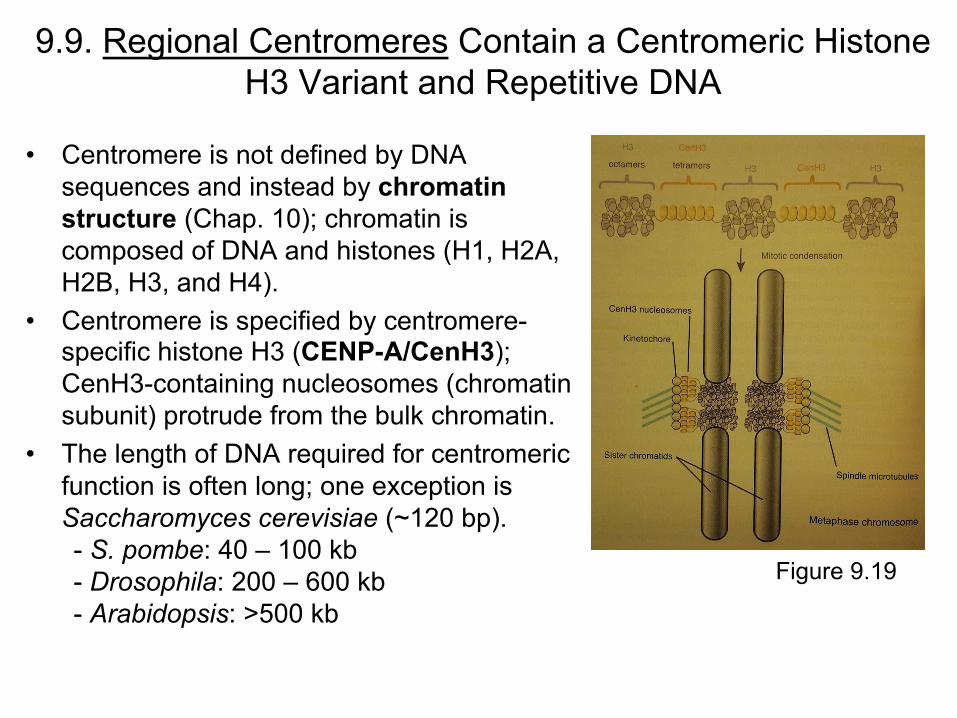
9.9. Regional Centromeres Contain a Centromeric Histone H3 Variant and Repetitive DNA
• Centromere is not defined by DNA sequences and instead by chromatin structure (Chap. 10); chromatin is composed of DNA and histones (H1, H2A, H2B, H3, and H4).
• Centromere is specified by centromere-specific histone H3 (CENP-A/CenH3); CenH3-containing nucleosomes (chromatin subunit) protrude from the bulk chromatin.
• The length of DNA required for centromeric function is often long; one exception is Saccharomyces cerevisiae (~120 bp). - S. pombe: 40 – 100 kb - Drosophila: 200 – 600 kb - Arabidopsis: >500 kb
Figure 9.19

• The sequences required for centromeric function fall within ~120 bp (CEN region) provides a useful tool to identify proteins that bind centromere and form kinetochore.
• Every chromosome of S. cerevisiae has a CEN region. Fragments from different chromosomes are interchangeable; CEN does not distinquish one chromosome from another.
• Three cell cycle-dependent elements (CDEs) in the CEN region: - CDE-I: 9bp, highly conserved. - CDE-II: >90% A-T rich sequence of 80-90 bp, its function depends on the length rather than its exact sequence. - CDE-III; 11bp, highly conserved, some mutations are tolerated but CCG are essential.
9.10. Point Centromeres in S. cerevisiae Contain Short, Essential Protein-Binding DNA Sequences
Figure 9.20. The S. cerevisiae centromere has a long A-T stretch flanked by short conserved sequences.
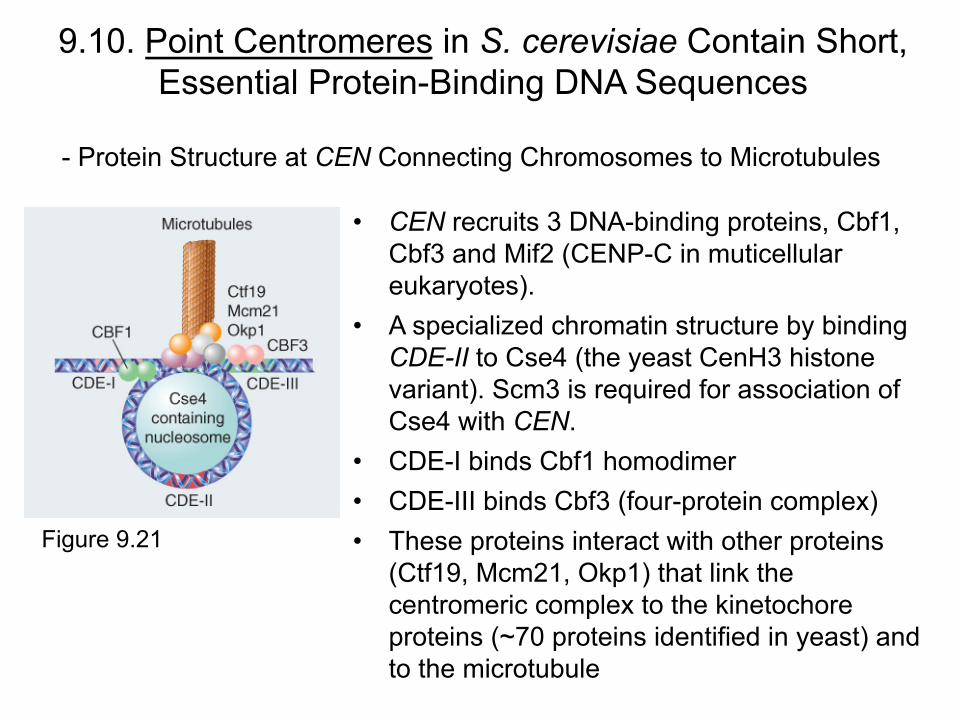
9.10. Point Centromeres in S. cerevisiae Contain Short, Essential Protein-Binding DNA Sequences
Figure 9.21
- Protein Structure at CEN Connecting Chromosomes to Microtubules
• CEN recruits 3 DNA-binding proteins, Cbf1, Cbf3 and Mif2 (CENP-C in muticellular eukaryotes).
• A specialized chromatin structure by binding CDE-II to Cse4 (the yeast CenH3 histone variant). Scm3 is required for association of Cse4 with CEN.
• CDE-I binds Cbf1 homodimer • CDE-III binds Cbf3 (four-protein complex) • These proteins interact with other proteins
(Ctf19, Mcm21, Okp1) that link the centromeric complex to the kinetochore proteins (~70 proteins identified in yeast) and to the microtubule

9.11. Telomeres Have Simple Repeating Sequences That Seal the Chromosome Ends
• The telomere is required for the stability of the eukaryotic chromosome (linear DNA).
• The cell must be able to distinguish the telomere (stable) from free DNA ends caused by damage.
• A long series of short, tandemly repeated sequences (100 -1000 repeats).
• A telomere consists of a simple repeat, Cn(A/T)m; n>1, m is 1 to 4.
• Highly conserved among eukaryotes.
• Extension of 3’ overhang G-T-rich strand.
Figure 9.22
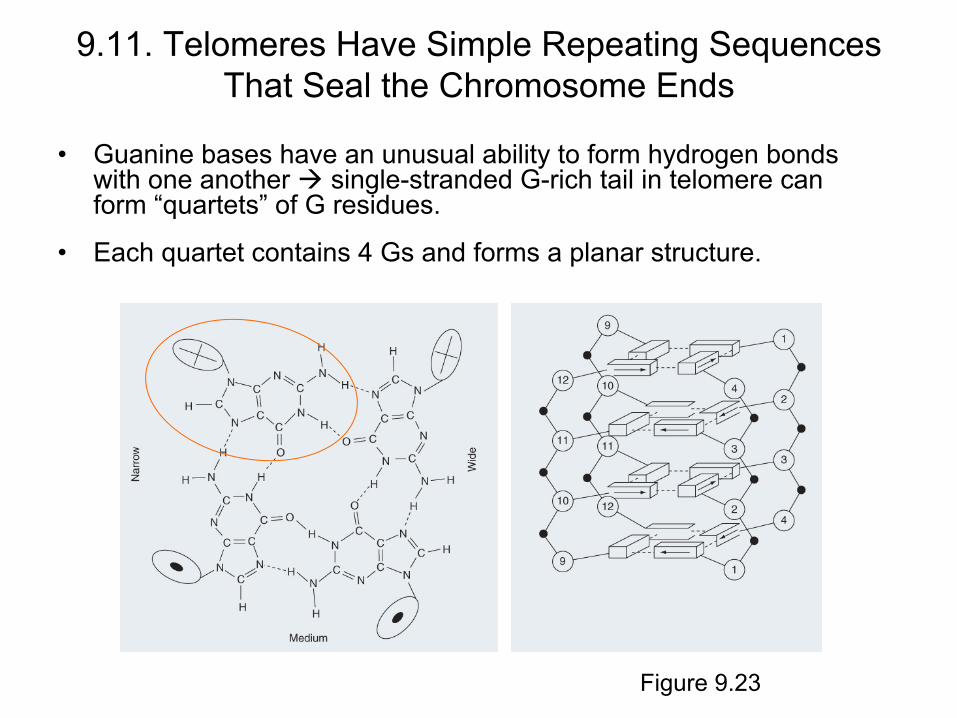
• Guanine bases have an unusual ability to form hydrogen bonds with one another single-stranded G-rich tail in telomere can form “quartets” of G residues.
• Each quartet contains 4 Gs and forms a planar structure.
9.11. Telomeres Have Simple Repeating Sequences That Seal the Chromosome Ends
Figure 9.23
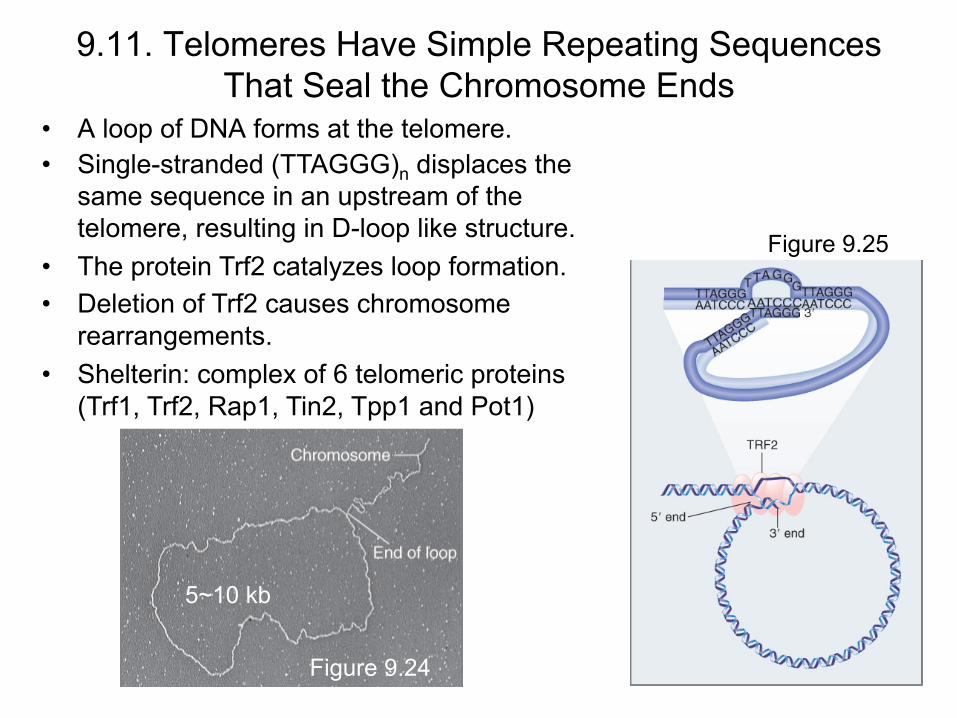
• A loop of DNA forms at the telomere.
5~10 kb
9.11. Telomeres Have Simple Repeating Sequences That Seal the Chromosome Ends
Figure 9.24
• Single-stranded (TTAGGG)n displaces the same sequence in an upstream of the telomere, resulting in D-loop like structure.
• The protein Trf2 catalyzes loop formation. • Deletion of Trf2 causes chromosome
rearrangements. • Shelterin: complex of 6 telomeric proteins
(Trf1, Trf2, Rap1, Tin2, Tpp1 and Pot1)
Figure 9.25

• Telomere has the ability to be extended, by addition of telomeric repeats to the end of the chromosome in every replication cycle.
• Telomerase uses the 3′–OH of the G + T telomeric strand to prime synthesis of tandem TTGGGG repeats.
• The RNA component (150~1300 nt) of telomerase has a sequence of 15~22 nt identical to two repeats of the C+A-rich repeating sequence.
• One of the protein subunits is a reverse transcriptase that uses the RNA as template to synthesize the G + T-rich DNA sequence.
• Discontinuous synthesis. • Telomere length (5~15 kb in human and ~300
bp in yeast) is determined by Est1 and Est3 in yeast but not by telomerase.
9.11. Telomeres Have Simple Repeating Sequences That Seal the Chromosome Ends
Telomere Extension by Telomerase Figure 9.27