
2, - Journal of...
10
JOURNAL OF BACTERIOLOGY Vol. 87, No. 2, p. 293-302 February, 1964 Copyright X 1964 by the American Society for Microbiology Printed in U.S.A. PIGMENTATION AND TAXONOMY OF THE GENUS XANTHOMONAS MORTIMER P. STARR AND WILLIAM L. STEPHENS' Department of Bacteriology, University of California, Davis, California Received for publication 26 August 1963 ABSTRACT STARR, MORTIMER P. (University of California, Davis), AND WILLIAM L. STEPHENS. Pigmentation and taxonomy of the genus Xanthomonas. J. Bac- teriol. 87:293-302. 1964.-The colonies formed by phytopathogenic bacteria of the genus Xantho- monas are typically yellow in color. This chromo- genesis stems from the presence in all yellow xanthomonads of a particular carotenoid pigment. This unique "Xanthomonas-carotenoid," which has not been found in any yellow nonxanthomonad, is characterized herein to the extent required for its recognition by relatively simple screening pro- cedures. In general, the occurrence of a carotenoid "alcohol" with absorption maxima at 418, 437, and 463 myu (petroleum ether)-in a gram-negative, polarly-flagellated, oxidative, rod-shaped bac- terium-would suggest placement of that micro- organism in the genus Xanthomonas. Many of the phytopathogenic bacteria are pigmented, and this chromogenesis is an im- portant determinative trait used practically in the isolation and identification of these micro- organisms. Nevertheless, with a few exceptions (Starr, 1944, 1958; Starr and Saperstein, 1953), only incidental attention has been devoted to the nature of their pigments. Most of the chromogenic phytopathogenic bacteria are referred to the genera Pseudomonas and Xanthomonas of the order Pseudomonadales. In naming the genus Xanthomonas, Dowson (1939) emphasized that the yellow color is one of the more significant characteristics of its species. This emphasis upon production of a yellow growth, and its use as a primary deter- minative feature for these bacteria, prompted a general study of the yellow coloring matter of Xanthomonas, which had been shown earlier (Starr, 1944) to be carotenoid in nature. Our work on the localization of this pigment in the 1 Present address: Department of Biology, Chico State College, Chico, Calif. cytoplasmic membrane of X. juglandis appeared recently (Stephens and Starr, 1963). The present contribution substantiates our belief that a unique carotenoid occurs universally in the yellow members of the genus Xanthomonas, and that it is not found in a representative collec- tion of yellow nonxanthomonad bacteria, in- cluding several which are erroneously referred to Xanthomonas. The possible taxonomic and deter- minative implications of these observations are discussed. The precise chemical nature of the unique Xanthomonas pigment is considered here only to the extent required for its recognition by relatively simple screening procedures. MATERIALS AND METHODS Cultures. Cultures were obtained from the International Collection of Phytopathogenic Bacteria (ICPB) maintained at the Department of Bacteriology, University of California at Davis, from the general stock culture collection of the same department, and from the American Type Culture Collection (ATCC). The forowing species (names, as received) and strains were used: Cellulomonas biazote (2074); C. fimi (2544); Erwinia anasas (ICPB-EA133); E. lathyri (ICPB-EL102); Flavobacterium acidificum (ATCC-8366); F. ferrugineum (ATCC-13524); F. heparinum (ATCC-13125); F. meningisepticum (ATCC-13253); F. resinovorum (ATCC-12524); P. rimaefaciens (ICPB-PR107); P. syringae (ICPB-PS169); Pseudomonas sp. (safflower-662- 11-1-yellow); X. badrii (ICPB-XB103); X. begoniae (ICPB-XB8); X. beticola (ICPB- XBl09Smr); X. carotae (ICPB-XC 139); X. cassiae (ICPB-XC141); X. corylina (ICPB- XC12); X. desmodii-gangeticii (ICPB-XD106); X. geranii (ICPB-XG10); X. holcicola (ICPB- XH3); X. hyacinthi (ICPB-XH1 10); X. juglandis (ICPB-XJ103); X. lespedezae (ICPB-XL2); X. manihotis (ICPB-XM12); X. nakatae-olitorii (ICPB-XN101); X. oryzae (ICPB-XO101); X. pelargonii (ICPB-XP8); X. phaseoli (ICPB- 293 on June 22, 2018 by guest http://jb.asm.org/ Downloaded from
Transcript of 2, - Journal of...

JOURNAL OF BACTERIOLOGYVol. 87, No. 2, p. 293-302 February, 1964Copyright X 1964 by the American Society for Microbiology
Printed in U.S.A.
PIGMENTATION AND TAXONOMY OF THE GENUS XANTHOMONAS
MORTIMER P. STARR AND WILLIAM L. STEPHENS'
Department of Bacteriology, University of California, Davis, California
Received for publication 26 August 1963
ABSTRACT
STARR, MORTIMER P. (University of California,Davis), AND WILLIAM L. STEPHENS. Pigmentationand taxonomy of the genus Xanthomonas. J. Bac-teriol. 87:293-302. 1964.-The colonies formed byphytopathogenic bacteria of the genus Xantho-monas are typically yellow in color. This chromo-genesis stems from the presence in all yellowxanthomonads of a particular carotenoid pigment.This unique "Xanthomonas-carotenoid," whichhas not been found in any yellow nonxanthomonad,is characterized herein to the extent required forits recognition by relatively simple screening pro-cedures. In general, the occurrence of a carotenoid"alcohol" with absorption maxima at 418, 437, and463 myu (petroleum ether)-in a gram-negative,polarly-flagellated, oxidative, rod-shaped bac-terium-would suggest placement of that micro-organism in the genus Xanthomonas.
Many of the phytopathogenic bacteria arepigmented, and this chromogenesis is an im-portant determinative trait used practically inthe isolation and identification of these micro-organisms. Nevertheless, with a few exceptions(Starr, 1944, 1958; Starr and Saperstein, 1953),only incidental attention has been devoted to thenature of their pigments.Most of the chromogenic phytopathogenic
bacteria are referred to the genera Pseudomonasand Xanthomonas of the order Pseudomonadales.In naming the genus Xanthomonas, Dowson(1939) emphasized that the yellow color is oneof the more significant characteristics of itsspecies. This emphasis upon production of ayellow growth, and its use as a primary deter-minative feature for these bacteria, prompted ageneral study of the yellow coloring matter ofXanthomonas, which had been shown earlier(Starr, 1944) to be carotenoid in nature. Ourwork on the localization of this pigment in the
1 Present address: Department of Biology, ChicoState College, Chico, Calif.
cytoplasmic membrane of X. juglandis appearedrecently (Stephens and Starr, 1963).The present contribution substantiates our
belief that a unique carotenoid occurs universallyin the yellow members of the genus Xanthomonas,and that it is not found in a representative collec-tion of yellow nonxanthomonad bacteria, in-cluding several which are erroneously referred toXanthomonas. The possible taxonomic and deter-minative implications of these observations arediscussed. The precise chemical nature of theunique Xanthomonas pigment is considered hereonly to the extent required for its recognition byrelatively simple screening procedures.
MATERIALS AND METHODS
Cultures. Cultures were obtained from theInternational Collection of PhytopathogenicBacteria (ICPB) maintained at the Departmentof Bacteriology, University of California atDavis, from the general stock culture collectionof the same department, and from the AmericanType Culture Collection (ATCC). The forowingspecies (names, as received) and strains wereused: Cellulomonas biazote (2074); C. fimi (2544);Erwinia anasas (ICPB-EA133); E. lathyri(ICPB-EL102); Flavobacterium acidificum(ATCC-8366); F. ferrugineum (ATCC-13524);F. heparinum (ATCC-13125); F. meningisepticum(ATCC-13253); F. resinovorum (ATCC-12524);P. rimaefaciens (ICPB-PR107); P. syringae(ICPB-PS169); Pseudomonas sp. (safflower-662-11-1-yellow); X. badrii (ICPB-XB103); X.begoniae (ICPB-XB8); X. beticola (ICPB-XBl09Smr); X. carotae (ICPB-XC 139); X.cassiae (ICPB-XC141); X. corylina (ICPB-XC12); X. desmodii-gangeticii (ICPB-XD106);X. geranii (ICPB-XG10); X. holcicola (ICPB-XH3); X. hyacinthi (ICPB-XH1 10); X. juglandis(ICPB-XJ103); X. lespedezae (ICPB-XL2); X.manihotis (ICPB-XM12); X. nakatae-olitorii(ICPB-XN101); X. oryzae (ICPB-XO101);X. pelargonii (ICPB-XP8); X. phaseoli (ICPB-
293
on June 22, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

STARR AND STEPHENS
XP104); "X." stewartii (ICPB-SS1, SS11, SS12, -SS18); X. taraxaci (ICPB-XT11); "X." trifolii(ICPB-XT109, "Bacterium herbicola" 2554);"X." uredovorus (ICPB-XU104); X. vasculorum(ICPB-XV24); X. vesicatoria (ICPB-XV3). Thequotation marks indicate our belief that thesespecies ("X." stewartii, "X." trifolii, and "X."uredovorus) are not correctly assigned to thegenus Xanthomonas.
Cultivation of the bacteria. Before use in thisstudy, each culture was checked for purity,Gram stain reaction, cell morphology, motility,and flagellation. The xanthomonad stock cultureswere grown on "YDC" agar slants consisting of1.0% Difco yeast extract, 1.0% glucose, 2.0%finely divided CaCO3, and 1.5% agar. These weretransferred to Difco nutrient broth (100 ml per250-ml Erlenmeyer flask); the flasks were incu-bated at 28 C on a rotary shaking machineoperating with an amplitude of 1.5 in. at a rateof 160 rev/min. After 24 hr, the 100-ml cultureswere checked microscopically for purity, and thentransferred in toto aseptically to 500 ml of nutrientbroth contained in 2-liter Erlenmeyer flasks,which were incubated for 24 hr under the sameconditions. After a second purity check, the cellswere harvested in a Servall centrifuge (typeSP/X) at 3,440 X g; a typical xanthomonadyielded about 4 g of moist packed cells per literof culture.
Of the 20 nonxanthomonad cultures, 18 werecultivated in a similar manner; however, becausemost of these cultures appeared to form lesspigment than the true Xanthomonas spp., addi-tional flasks of culture medium were used, andthe cells were harvested in a Sharples Super-Centrifuge at 50,000 rev/min. F. ferrugineumand Pseudomonas sp. "662-11-1-yellow" did notproduce pigmented cells when cultivated in theabove manner; they were grown on YDC agarplates at 30 C and, after sufficient growth, thepigmented cells were scraped off.
Solvents. The following solvents were allreagent grade of Allied Chemical Corp., NewYork, N.Y.: petroleum ether (b.r. 30 to 60 C),methyl alcohol (absolute), diethyl ether (an-hydrous), and benzene. The petroleum ether wasfurther purified by passage through a silicic acidcolumn, and the diethyl ether was redistilledand stored over reduced iron.
Extraction of the pigments. Approximately 50ml of absolute methanol were added per g ofmoist bacterial cells, and the container was im-
mersed in hot water to bring the methanolquickly to the boiling point and thus extract thepigments in a very few minutes. The solution wasthen cooled, and centrifuged to remove the cells.Direct controls revealed that this brief exposureto an elevated temperature produced no observa-ble injurious effects on the pigments.
Separation of types of pigments. The methanolextract of each organism was partitioned betweenimmiscible solvents to separate the carotenoidsinto four empirical groups: "hydrocarbons,""alcohols," "esters," and "acids." It must beemphasized that these terms, as applied to crudepigment preparations, do not necessarily relateto actual chemical structure-rather, they meanthat the crude pigment partitions in the mannerexpected of pure carotenoid hydrocarbons,alcohols, esters, or acids. There is a substantialeffect of impurities upon this partition behavior,and much confusion has resulted from any morerigorous interpretation of such partition tests.This partition procedure (Fig. 1) is a modificationof the scheme of Sobin and Stahly (1942) whichin turn derives from the method of Kuhn andBrockmann (1932). We recommend close ad-herence to the following procedural details asessential for reproducibility.
Petroleum ether was added to the methanoliccell extract to make a monophasic system, afterwhich enough water was added to give a 90%alcohol concentration (neglecting, however, the<2% water in the cell extract). The mixturewas shaken in a separatory funnel, but verygently to avoid formation of an emulsion. Toinsure complete separation, the petroleum etherlayer was extracted repeatedly with fresh samplesof 90% methanol, and the methanol phase wassimilarly partitioned several times against freshpetroleum ether. This partition test should firstbe performed on a small sample; if all pigmentsremain in the methanol (i.e., are hypophasic),the bulk of the methanolic extract may then besaponified and partitioned against diethyl ether.The petroleum ether layer was then washed
with distilled water by means of a continuous-flow device, dehydrated with sodium sulfate,and taken to dryness in vacuo with a Rincoevaporator. The dried material may containcarotenoid hydrocarbons and esters. This residuewas then dissolved in 3% KOH-methanol, andsaponified at 40 C for 3 hr. An equal volume ofpetroleum ether was added, followed by enoughwater to give a 90% alcohol concentration; then
294 J. BACTERIOL.
on June 22, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

XANTHOMONAS PIGMENTATION AND TAXONOMY
To the METHANOLIC EXTRACT OF CELLS, add an equal volume of petroleum ether (b.r.,30 to 60 C), followed by enough distilled water to give a methanol concentration of 90%. Shake gentlyin a separatory funnel.
PETROLEUM ETHER EPIPHASE(contains "hydrocarbons" and "esters")
Evaporate to dryness. Saponify with 3% meth-anolic KOH (40 C for 3 hr). Add an equal volumeof petroleum ether and enough water to again givea 90% methanol concentration.
METHANOL HYPOPHASE(contains "alcohols" and "acids")
Add methanolic KOH to final concentration of3% KOH, hold at 40 C for only 5 min (mild saponi-fication). Add an excess of diethyl ether, followedby water, and shake gently in a separatory funnel.
PETROLEUMETHER POST-
SAPONIFICATIONEPIPHASE
(contains "hydro-carbons")
METHANOL POST-SAPONIFICATIONHYPOPHASE(contains freed"alcohols")
DIETHYL ETHEREPIPHASE
(contains "alcohols")
AQUEOUSHYPOPHASE(contains saltsof "acids")
Acidify, and extractfreed "acids" with di-ethyl ether.
FIG. 1. Scheme for empirical separation of bacterial carotenoid pigments.
the mixture was shaken very gently in a separa-tory funnel. The petroleum ether epiphase wasremoved, washed, dehydrated with sodium sul-fate, and dried in vacuo. The individual carote-noid hydrocarbons (i.e., carotenes) in the residuewere then separated by column chromatography.The de-esterified carotenoid alcohols in the
KOH-methanol hypophase were transferred intodiethyl ether by the addition first of that solventand then of distilled water. The ether layer waswashed free of the methanol-KOH with water,freed of water by treatment with a saturatedsolution of NaCl followed by the addition ofanhydrous sodium sulfate, and then dried invacuo. These de-esterified carotenoid alcoholswere then separated by adsorption chromatog-raphy.
Returning now to the original methanolichypophase (or the original methanolic cell ex-tract), we deviated slightly from the Sobin-Stahlyprocedure, which calls for addition of just enoughKOH to make the solution alkaline to litmus.Instead, KOH-methanol was added to themethanolic solution of the pigments to a finalKOH concentration of 3% and the solution washeated gently (40 C for 5 min); then 2 volumes ofdiethyl ether were added to make a monophasicsystem, followed by enough water to separatetwo layers. This mixture was shaken very gently
in a separatory funnel. The hypophase wasdrained off and, if still colored, was partitionedrepeatedly against fresh samples of diethyl'ether. The ether layer, which contained thecarotenoid alcohols, was withdrawn, washed,.dehydrated, and evaporated to dryness by themethods previously mentioned. It must beemphasized that the best possible extraction ofthe carotenoid alcohols (i.e., xanthophylls)requires that the water be added to the homo-geneous solution of diethyl ether and methanolicpigment extract. Strain (1938) showed thatalcohol or acetone solutions of xanthophyllsform colloidal solutions when diluted with waterand, moreover, the pigments cannot be extractedfrom these solutions with solvents that are im-miscible with water. The formation of thesecolloidal solutions occurs most easily in thepresence of the saponified materials and thecomplex substances which are in the alkalinelipid extracts.Any carotenoid acids would remain in the
aqueous phase in the form of their salts, andmight be recovered by the addition of dilutehydrochloric acid, followed by extraction withdiethyl ether.The method described above was used on the
crude methanol extracts primarily to isolatecarotenoids of similar solubility properties, and
VOL. 87, 1964 295
on June 22, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

STARR AND STEPHENS
thereby to effect a certain amount of purificationprior to chromatography. It should again bestressed that partition tests performed upon suchcrude lipid extracts can give quite different resultsfrom those performed on pure carotenoids, andthat the designations "hydrocarbon," "alcohol,""ester," and "acid" are merely a convention.The presence of other lipids interferes with thepartitioning of the different carotenoid types.Partition tests used in the chemical characteriza-tion of a carotenoid should be performed on a
pure sample.Chromatographic adsorption. After separating
the carotenoids into the aforementioned groups,the individual carotenoids of each group were
isolated by column chromatography. The ex-
tracts were adsorbed from dry benzene solutionsonto columns of magnesia-Celite, 1:1 by weight(MgO, Baker's chemically pure analyzed;Celite, L-665-A of Johns-Manville, New York,N.Y.). We used Pyrex chromatography tubes(16 by 130 mm) with coarse-porosity fritteddiscs sealed in the inner member of the 1%5ground joint. Columns were packed dry withgentle suction, then washed with benzene forcedthrough the adsorbent with nitrogen. Loadingof the samples onto the columns and develop-ment of the chromatograms were also accom-
plished with nitrogen under positive pressure.
Development was generally achieved with ben-zene. After separation of the pigments, thecolumns were allowed to run dry, and the zones
were separated mechanically and eluted withmethanol.
Spectrometric analysis. All spectrometric anal-yses were performed with a Beckman recordingspectrophotometer (model DK-2). The absorp-tion maxima of the individual carotenoids were
determined in petroleum ether, benzene, andabsolute methanol. The spectrophotometer was
calibrated with a Hanovia mercury lamp andchecked periodically with the wavelength calibra-tion standards listed in Beckman instructionmanual 305-A.
RESULTSBehavior upon partition tests. The procedure
described for separating the four major types ofcarotenoids can be abridged somewhat for thepigments of Xanthomonas species. Since theseparticular pigments were always hypophasicwhen a 90% methanolic crude extract was
partitioned against petroleum ether, this parti-
tion need be tried only on a small sample toverify the absence of epiphasic pigments. Noepiphasic pigments were found in any xanthomonads, and the bulk of the crude methanolextract was further processed as though it werethe "original methanolic hypophase."
Chromatograms and absorption maxima. Table1 contains the results of the chromatographicand spectrophotometric observations of thepigments found in the xanthomonads. Thedesignations given to the pigments in this tableare arbitrary and serve merely to identify thespecific carotenoids. Zones are numbered in theorder in which they appear on the columns fromtop to bottom.Of the 20 Xanthomonas cultures examined, all
19 yellow-pigmented species contained carote-noids; only the colorless X. manihotis was devoidof these pigments. All carotenoids found in eachof the yellow species had partition behaviors,in crude extracts, like those of carotenoid alco-hols. In every case, the pigment in at least onezone of the chromatogram, when eluted andexamined photometrically, showed an absorptionspectrum with maxima of (418), 437, and 463m,u in petroleum ether; (435), 454, and 481 m,uin benzene; and (420), 441, and (468) m,u inmethanol. (Parentheses around an absorptionmaximum indicate a shoulder or broad centralpeak centered around that approximate wave-length.) More often, the pigments in two separatezones of the column possessed this particular setof maxima. In such cases, the pigments in theupper zone (designated type 2) appear to beoxidized or oxidatively deteriorated forms of thepigment in the lower zone, the "universal pig-ment" of the yellow xanthomonads.
Pigments 1 and 2 are further distinguishablefrom one another on the basis of spectral con-figuration (Fig. 2). It should not be inferred thatthese are two distinctive carotenoids. Althoughthe lower zone (pigment 1) is spectrally homo-geneous, a spectral gradient is apparent whenthe upper zone is mechanically divided intosections. This gradient (Fig. 3) ranges from pig-ments with bathochromic (high-wavelength)peaks and hypsochromic (low-wavelength) shoul-ders of equal height through pigments withbathochromic peaks much lower in absorbancethan the corresponding hypsochromic shoulders.In extreme examples of this latter case, thebathochromic peak actually disappears, and thehypsochromic shoulder blends into the principal
296 J. BAOTERIOL.
on June 22, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

XANTHOMONAS PIGMENTATION AND TAXONOMY
TABLE 1. Absorption maxima of the carotenoids of Xanthomonas
Adsorption behaviorb . Absorption maxima (m,s)dSpecies Culture designationa - _ .
Zone Color to Petroleum ether Benzene Methanol
X. badrii ....I ICPB-XB103
X. begoniae....I ICPB-XB8
X. beticola.....
X. carotae......
ICPB-XB109Smr
ICPB-XC139
X. cassiae..... ICPB-XC141
X. corylina...X. desmodii-
gangeticii....
ICPB-XC12
ICPB-XD106
X. geranii.....1 ICPB-XG1O
X. holcicola.... ICPB-XH3
X. hyacinthi... ICPB-XH110X. juglandis... ICPB-XJl03
X. lespedezae... ICPB-XL2
X. manihotis...X. nakatae-
olitorii.......
ICPB-XM12
ICPB-XN101
X. oryzae. ICPB-XO101
X. pelargonii ... ICPB-XP8
X. phaseoli....1I ICPB-XP104
X. taraxaci..... ICPB-XT11
X. vasculorum ..I ICPB-XV24
X. vesicatoria. ICPB-XV3
IIII
III
III
IIIIII
III
IIIIIII
IIII
III
II
IIII
III
III
III
III
III
II
RedOrangeRedOrangeOrangeYellowRedOrangeYellowRedOrangeOrange
RedOrangeRed-orangeOrange-yellowRedOrangeOrangeRedOrangeRedOrange
OrangeYellowRed-orangePale orangeRedOrangeOrangeYellowRedOrangeOrangePale yellowRedOrange
512212
13412
2
2
212121122121
2121312121121121
(415) 433 459(418) 437 465(418) 437 463(418) 437 463(418) 437 463(418) 437 463(401) 420 445(398) 417 442(418) 437 463(418) 437 463(418) 437 463(418) 437 463
(418) 437 463(418) 437 464(418) 437 463(418) 437 463(418) 437 463(418) 437 463(418) 437 463(418) 437 463(418) 437 463(418) 437 463(418) 437 463
(418) 437 463(418) 437 463(418) 437 463(418) 437 463(401) 420 443(418) 437 464(418) 437 463(418) 437 463(418) 437 463(418) 437 463(418) 437 463(418) 437 465(418) 437 463(418) 437 463
(433) 451 479(435) 454 481(435) 454 481(435) 454 481(435) 454 481(435) 454 481(415) 436 460(412) 431 457(435) 454 481(435) 454 481(435) 454 481(435) 454 481
(435)(435)(435)(435)(435)(435)(435)(435)(435)(435)(435)
(435)(435)(435)(435)(415)(435)(435)(435)(435)(435)(435)(435)(435)(435)
454 481454 481454 481454 481454 481454 481454 481454 481454 481454 481454 481
454 481454 481454 481454 481436 461454 482454 483454 481454 483454 483454 483454 483454 483454 483
(419) 438 (462)(420) 441 (468)(420) 441 (468)(420) 441 (468)(420) 441 (468)(420) 441 (468)(425) (445) -
(425) (443) -(420) 441 (468)(420) 441 (468)(420) 441 (468)420) 441 (468)
(420)420(420)(420)(420)(420)(420)(420)(420)(420)(420)
441 (468)441 (468)441 (468)441 (468)441 (468)441 (468)441 (468)441 (468)441 (468)441 (468)441 (468)
(420) 441 (468)(420) 441 (468)(420) 441 (468)(420) 441 (468)(426) (445) -
(420) 441 (468)(420) 441 (468)(420) 441 (468)(420) 441 (468)(420) 441 (468)(420) 441 (468)(420) 441 (468)(420) 441 (468)(420) 441 (468)
a ICPB = International Collection of Phytopathogenic Bacteria, Department of Bacteriology,University of California, Davis (M. P. Starr, Curator).
I Zones are numbered in the order in which they appear on the columns, from top to bottom. Basedupon partition tests of crude pigments as explained in the text, all pigments could be categorized-somewhat arbitrarily as is the custom in carotenoid research-as "alcohols."
¢ The numbers given to the pigments serve to identify the specific carotenoids in the text.d Parentheses around an absorption maximum indicate a shoulder or broad central peak centered
around that approximate wavelength.
297VOL. 87, 1964
on June 22, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

STARR AND STEPHENS
D400 425MrL
FIG. 2. Absorption spectra in petroleum ether ofpigments types 1 and 2 of Xanthomonas.
I
m | :(-)z
-cxA
400 425 450 475MEL
FIG. 3. Diagrammatic representation of the spec-
tral gradient which is apparent when a single chro-matogram zone containing pigment type 2 is mechan-ically divided into successive sections. The differentspectral configurations (in petroleum ether) are
arranged in the order in which they occur in thechromatogram zone, from top to bottom.
peak, thus presenting a single broad-band spec-
trum. The pigment designated type 1 has a
bathochromic peak relatively high in absorbancecompared to its hypsochromic shoulder.Under certain conditions, pigment type 1 can
be converted into pigment type 2. When thisconversion takes place, there is an accompanying
change in chromatographic behavior. Upondevelopment with benzene, pigment 1 movesdown a MgO-Celite column, while pigments oftype 2 remain at the top of the column. Occa-sionally, intermediate types are found: thoseforms which have a spectral configuration likepigment 1, but remain at the top of the chromato-graph column like pigment 2, are designatedpigment 12; those forms which have the spectralconfiguration of pigment 2, but chromatographlike pigment 1, are designated pigment 21.
Table 1 shows that X. geranii yielded twobands, when chromatographed on MgO-Celiteand developed with benzene. The top band helda carotenoid alcohol with a pigment 2 spectralconfiguration, while the lower band containedpigment 1. This was the most common patternencountered in the survey. Other species con-taining only pigments 1 and 2 were X. beticola,X. desmodii-gangeticii, X. holcicola, X. juglandis,X. lespedezae, X. nakatae-olitorii, X. oryzae, X.phaseoli, and X. taraxaci.Chromatograms of X. corylina always con-
tained a single band, which proved to be acarotenoid alcohol with the pigment 2 configura-tion. X. carotae forms three carotenoid alcohols,one with the spectral configuration and chromato-graphic behavior of pigment 1, and the othertwo, pigments 3 and 4, with absorption maximaat much lower wavelengths. X. begoniae and X.cassiae each yielded two bands when chromato-graphed on MgO-Ce]ite; the pigments in bothtop and bottom bands had the spectral configura-tion of pigment 2. The top bands actually con-tained pigment 2; because the pigment in thebottom bands chromatographed like pigment 1,it was considered to be pigment 21. X. hyacinthihad a single pigment with type 1 spectral con-figuration, but which remained (like pigment 2)at the top of a MgO-Celite column after develop-ment with benzene; hence, it was pigment 12.X. vasculorum and X. vesicatoria contained pig-ment 12 and, in addition, they each had pigment1. X. pelargonii contained two carotenoid alco-hols; namely, pigment 1 and a carotenoid verysimilar to the pigment 3 found in X. carotae. X.badrii had two carotenoid alcohols; besides pig-ment 1, there was found pigment 5, with absorp-tion maxima at wavelengths slightly lower thanthose of pigments 1, 12, 2, and 21.
Chromatograms and absorption maxima of
298 J . BACTERIOL .
on June 22, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

XANT'HOMONAS PIGMENTATION AND TAXONOMY
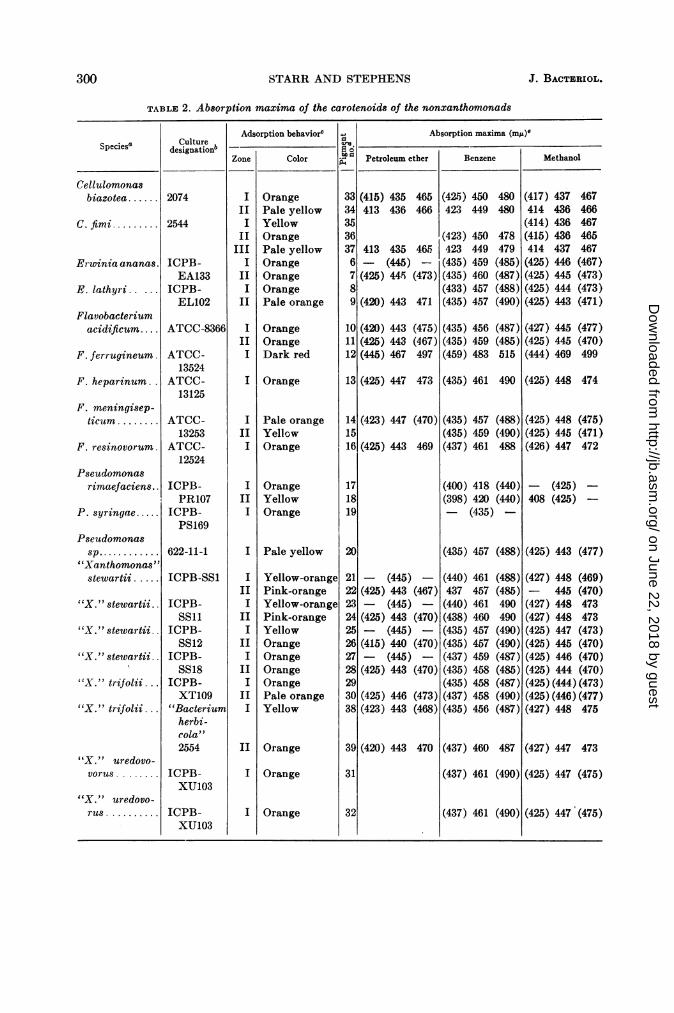
nonxanthomonads. Table 2 contains the resultsof the chromatograms and the absorption maximaof all pigments found in the survey of yellow-pigmented species from the genera Cellulomonas,Erwinia, Flavobacterium, and Pseudomonas, andin three doubtful species of Xanthomonas: X.stewartii, X. trifolii (including a strain receivedas Bacterium herbicola), and X. uredovorus. Thenumbers given to the pigments in this table arearbitrary and are used merely to label the severalcarotenoids. Zones are numbered in the order inwhich they appear on the columns from top tobottom. Parentheses around an absorptionmaximum indicate a shoulder or a broad centralpeak centered around that approximate wave-length.
These 20 cultures of yellow nonxanthomonadseach contained carotenoids, but none of themcontained the "Xanthomonas-carotenoid." Thepartition behavior of all these carotenoids incrude lipid extracts was like that of carotenoidalcohols, which was true also for the Xantho-monas pigments. However, the carotenoids ofthe nonxanthomonads move into the diethylether phase from alkaline aqueous methanolmuch more rapidly than does the Xanthomonaspigment. The fine details of the spectral absorp-tion curves of the xanthomonad and nonxantho-monad pigments differ. Furthermore, the "Xantho-monas-carotenoid" has a relatively greatersharpness of the absorption peaks in petroleumether rather than in methanol; most of the pig-ments from nonxanthomonads reverse this pat-tern and have the sharper resolution in methanol.Although all the pigments described here parti-tion in crude lipid extracts like carotenoidalcohols, other types of carotenoids were de-tected in additional control cultures treated inthe same manner. Carotenoids which partitionlike hydrocarbons and alcohols were found inRhodospirillum rubrum; pigments which parti-tion like carotenoid hydrocarbons, alcohols, andesters were found in an unidentified fruitingmyxobacterium.
Since the primary purpose of this survey wasto determine whether the Xanthomonas pigmentscould be found outside that genus, these otherpigments are not described here in any detail.The partition and adsorption behavior, andabsorption maxima of each pigment, are pre-sented in Table 2. It is quite possible that the
same pigment was isolated from more than oneof the examined strains; however, no attemptswere made to check duplication, and the pig-ments from each organism were given separatenumbers. The most significant information gainedfrom this survey of nonxanthomonads was theabsence of the "Xanthomonas-carotenoid" fromyellow bacteria other than Xanthomonas.
DISCUSSION
In everyday phytobacterial practice, the yel-low color of Xanthomonas colonies is the keydiscriminating feature which-rationally ornot-is used to distinguish the xanthomonadsfrom other phytopathogenic or saprophyticbacteria isolated from plants (Burkholder andStarr, 1948). This pigmentation can now beattributed to formation of presumably oxy-genated carotenoids with absorption-spectrummaxima at 418, 437, and 463 m,u in petroleumether. All 19 yellow Xanthomonas spp. examinedin the present study were found to produce thisdistinctive pigment, which exists in a cluster ofrelated forms. An alcoholic carotenoid with thesecharacteristic properties has hitherto not beenreported in the literature (Fox, 1953; Goodwin,1955; Karrer and Jucker, 1950; Zechmeister,1934). This unique "Xanthomonas-carotenoid"is not produced by any of the other yellow non-xanthomonad bacteria examined in the presentstudy (Cellulomonas, Erwinia, Flavobacterium,Pseudomonas, and a few incorrectly designated"Xanthomonas" spp.), nor by any of the yellowphytopathogenic corynebacteria previously ex-amined by our group (Starr and Saperstein, 1953;Saperstein, Starr and Filfus, 1954).
Insofar as our present study is representative,the demonstration that distinctive carotenoidsoccur universally in the genus Xanthomonas-and, moreover, that they are restricted to thatgroup-has considerable taxonomic significance.First of all, one could conclude that the forma-tion of a carotenoid with absorption maxima at418, 437, and 463 m,u (petroleum ether)-by agram-negative, polarly-flagellated, oxidative, rod-shaped bacterium-would strongly suggest place-ment in the genus Xanthomonas. Certainly ourexperience to date makes it likely that possessionsolely of other carotenoids might provide apracticable basis for excluding yellow bacteriafrom the genus Xanthomonas. Arguing in another
299VOL. 87, 1964
on June 22, 2018 by guesthttp://jb.asm
.org/D
ownloaded from
STARR AND STEPHENS
TABLE 2. Absorption maxima of the carotenoids of the nonxanthomonads
Adsorption behaviorc Absorption maxima (ms)6Culture Zon | o r , Ptl m t rn n | h oSpecies5 designationb 6- - ________ _______
Zone Color Io0 Petroleum ether Benzene Methanol
Erwinia ananas.
E. lathyri.....
Flavobacteriumacidifictum....
F. ferrugineum.
F. heparinum.
F. meningisep-ticum ........
F. resinovorum.
Pseudomonasrimaefaciens..
P. syringae.....
Pseudomonassp............
"Xanthomonas"stewartii ....
"X." stewartii.
"X." stewartii.
"X." stewartii.
"X." trifolii.
"X." trifolii...
"X." uredovo-vorus.
"X. " uredovo-rus ICPB-
XU103
I Pale yellow
Yellow-orangePink-orangeYellow-orangePink-orangeYellowOrangeOrangeOrangeOrangePale orangeYellow
II Orange
(415) 435 465 (42.5) 450 480413 436 466 423 449 480
435 465f(445) -
445 (473)
(420) 443 471
(420) 443(425) 443(445) 467
(475)(467)497
(423) 450423 449(435) 459(435) 460(433) 457(435) 457
(435) 456(435) 459(459) 483
478479(485)(487)(488)(490)
(487)(485)515
(417) 437414 436(414) 436(415) 436414 437(425) 446(425) 445(425) 444(425) 443
467466467465467(467)(473)(473)(471)
(427) 445 (477)(425) 445 (470)(444) 469 499
131(425) 447 473 1(435) 461 490 1(425) 448 474
(423) 447
(425) 443
(470)
469
- (445) -
(425) 443 (467)- (445)-(425) 443 (470)- (445) -
(415) 440 (470)- (445) -
(425) 443 (470)
(425) 446 (473)(423) 443 (468)
(435) 457 (488)(435) 459 (490)(437) 461 488
(400) 418 (440)(398) 420 (440)- (435) -
(425) 448 (475)(425) 445 (471)(426) 447 472
- (425) -
408 (425) -
(435) 457 (488)1(425) 443 (477)
(440) 461 (488)437 457 (485)(440) 461 490(438) 460 490(435) 457 (490)(435) 457 (490)(437) 459 (487)(435) 458 (485)(435) 458 (487)(437) 458 (490)(435) 456 (487)
(427) 448 (469)- 445 (470)(427) 448 473(427) 448 473(425) 447 (473)(425) 445 (470)(425) 446 (470)(425) 444 (470)(425) (444) (473)(425)(446)(477)(427) 448 475
391(420) 443 470 (437) 460 487 (427) 447 473
(437) 461 (490)1(425) 447 (475)
r(437) 461 (490)
Cellulomonasbiazotea ...... 2074
C. fimi.. 2544
IIII
IIIII
IIII
II
IIII
OrangePale yellowYellowOrangePale yellowOrangeOrangeOrangePale orange
OrangeOrangeDark red
413
(425)
33343536376789
101112
I Orange
IIII
IIII
Pale orangeYellowOrange
OrangeYellowOrange
ICPB-EA133
ICPB-EL102
ATCC-8366
ATCC-13524
ATCC-13125
ATCC-13253
ATCC-12524
ICPB-PR107
ICPB-PS169
622-11-1
ICPB-SS1
ICPB-SS11
ICPB-SS12
ICPB-SS18
ICPB-XT109
"Bacteriumherbi-cola"2554
ICPB-XU103
141516
171819
20
2122232425262728293038
31
IIII
III
III
III
III
I Orange
300 J . BACTERIOL.
II Orange (425) 447 (475)321
on June 22, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

XANTHOMONAS PIGMENTATION AND TAXONOMY
direction, the fact that all yellow Xanthomonas"species" form the same carotenoids might beused to support the contention (Starr, 1959)that many of the present "species" are indeedphytopathogenic formae speciales of a limitednumber of true species. We must note that un-pigmented mutant strains of normally yellowXanthomonas spp. have been encountered occa-sionally. Breed, Murray, and Smith (1957)assigned seven normally achromogenic species toXanthomonas; we have already shown that oneof these-X. manihotis-forms no carotenoids.Colorless xanthomonads, whether naturallyoccurring or artificially induced, pose the usualsystematic dilemma which emerges whenever toogreat reliance is placed upon a single determina-tive trait; however, this quandary should notdetract from the basic utility of the foregoingtaxonomic recommendation.
This basic utility has already been demon-strated. Through the years, yellow bacteria ofuncertain systematic position have been placedin the genus Xanthomonas. X. stewartii, X.trifolii, and X. uredovorus are three examples ofwhat we and others consider to be erroneousplacement. In the present study, all strains ofthese three species were found to be devoid of the"Xanthomonas-carotenoid," thus supporting theirexpulsion from the genus Xanthomonas whichhad been advocated on other bases (Burkholderand Starr, 1948; Hayward and Hodgkiss, 1961;Dye, 1962, 1963; Starr and Stephens, 1963).
ACKNOWLEDGMENT
This study was supported in part by researchgrant AI-01067 from the National Institutes ofHealth, U.S. Public Health Service.
LITERATURE CITEDBREED, R. S., E. G. D. MURRAY, AND N. R. SMITH.
1957. Bergey's manual of determinative bac-teriology, 7th ed. The Williams & Wilkins Co.,Baltimore.
BURKHOLDER, W. H., AND M. P. STARR. 1948. Thegeneric and specific characters of phytopatho-genic species of Pseudomonas and Xantho-monas. Phytopathology 38:494-502.
DOWSON, W. J. 1939. On the systematic positionand generic names of the gram negative bac-terial plant pathogens. Zentr. Bakteriol.Parasitenk. Abt. II 100:177-193.
DYE, D. W. 1962. The inadequacy of the usual de-terminative tests for the identification ofXanthomonas spp. New Zealand J. Sci. 5:393-416.
DYE, D. W. 1963. The taxonomic position of Xan-thomonas uredovorus Pon et al. 1954. New Zea-land J. Sci. 6:146-149.
Fox, D. 1953. Animal biochromes and structuralcolors. Cambridge University Press, Cam-bridge, England.
GOODWIN, T. W. 1955. Carotenoids. Ann. Rev. Bio-chem. 24:497-522.
HAYWARD, A. C., AND W. HODGKISS. 1961. Taxo-nomic relationships of Xanthomonas uredo-vorus. J. Gen. Microbiol. 26:133-140.
KARRER, P., AND E. JUCKER. 1950. Carotenoids.Elsevier Publishing Co., Inc., New York.
KUHN, R., AND H. BROCKMANN. 1932. Bestimmungvon Carotinoiden. Z. Physiol. Chem. 206:41-64.
SAPERSTEIN, S., M. P. STARR, AND J. A. FILFUS.1954. Alterations in carotenoid synthesis ac-companying mutation in Corynebacteriummichiganense. J. Gen. Microbiol. 10:85-92.
SOBIN, B., AND G. L. STAHLY. 1942. The isolationand absorption spectrum maxima of bacterialcarotenoid pigments. J. Bacteriol. 44:265-276.
STARR, M. P. 1944. Studies of phytopathogenic bac-a The quotation marks indicate our belief that these three species ("X." stewartii, "X." trifoliji,
and "X." uredovorus) are not correctly assigned to the genus Xanthomonas.b ICPB = International Collection of Phytopathogenic Bacteria, Department of Bacteriology, Uni-
versity of California, Davis (M. P. Starr, Curator). ATCC = American Type Culture Collection,Washington, D.C. Cultures 2074, 2544, and 2554 are part of the general collection maintained by theDepartment of Bacteriology, University of California, Davis. Culture 622-11-1 was received from D. C.Erwin, University of California, Riverside.
¢ Zones are numbered in the order in which they appear on the columns, from top to bottom. Basedupon partition tests of crude pigments as explained in the text, all pigments could be categorized-somewhat arbitrarily as is the custom in carotenoid research-as "alcohols."
d The numbers given to the pigments serve merely to identify the specific carotenoids for possiblefuture reference. The pigments from each organism in this table were given separate numbers; however,it is quite possible that the same pigment was isolated from more than one of the examined strains.
e Parentheses around an absorption maximum indicate a shoulder or broad central peak centeredaround that approximate wavelength.
301VOL. 87, 1964
on June 22, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

302 STARR AND
teria. Cornell University, Abstracts of theses,1943, p. 349-350.
STARR, M. P. 1958. The blue pigment of Corynebac-terium insidiosum. Arch. Mikrobiol. 30:325-334.
STARR, M. P. 1959. Bacteria as plant pathogens.Ann. Rev. Microbiol. 13:211-238.
STARR, M. P., AND S. SAPERSTEIN. 1953. Thiamineand the carotenoid pigments of Corynebac-terium poinsettiae. Arch. Biochem. Biophys.43:157-168.
STEPHENS J. BACTERIOL.
STARR, M. P., AND W. L. STEPHENS. 1963. Pig-mentation and taxonomy of the genus Xan-thomonas. Bacteriol. Proc., p. 11.
STEPHENS, W. L., AND M. P. STARR. 1963. Localiza-tion of carotenoid pigment in the cytoplasmicmembrane of Xanthomonas juglandis. J. Bac-teriol. 86:1070-1074.
STRAIN, H. H. 1938. Leaf xanthophylls. CarnegieInst. Wash. Publ. 490, 1-147.
ZECHMEISTER, L. 1934. Carotenoide. JuliusSpringer, Berlin, Germany.
on June 22, 2018 by guesthttp://jb.asm
.org/D
ownloaded from


















