
Ascending Combinatorial Auctions Andrew Gilpin November 6, 2007.

121. Gilpin, M., G.A.E. Gall and D.S. Woodruff. Ecological dynamics and agricultural landscapes. In: Integrating Conservation Biology and Agricultural Production. Special Issue of Agriculture, Ecosystems and Environment 42:27-52. (1992b).
BOOK CHAPTER

I:UAgriculture. Ecosystems and Environment. 42 (1992) 27-52Elsevier Science Publishers B.V.. Amsterdam
Ecological Dynamics and Agricultural Landscapes
MICHAEL GILPINl, GRAHAM A.E. GALL 2 and DAVID S. WOODRUFfl
I Depanmenr of Bi%Kl, University of California, La Jolla; CA 92093 (U.S.A.)2 Depanmenr of Anima/ Science, University of California, Davis, CA 95616 (U.S.A.)
ADSTRACf
Gilpin, M., Gall, G.A.E. and Woodruff, 0.5., 1992. Ecological dynamics and agriculturallandscapcs. Agric.Ecosyslems Environ., 42: 27-52.
The planet Eanh is having difficulty under the stresses imposed by the diverse demands of an everincreasing human population. The problem appears to stem not only Crom population pressures but alsoCrom an imbalance between the needs and desires oC society. Agriculture, broadly defined to includefarming. rlShing,forestry and grazing systems. plays a significant role in the management of land, water, andbiological resources. This paper provides an analysis of opponunities for interaction between ecologicalscience and agriculture. The tong-term stability or agric\lhure is dependent on natural sources or genetiCmaterial Many pans oC society see a connict between conservation of biological resources and theirexploitation by agriculture. Agricultural production is essential to SOCietyand also can provide stewardshipof biological resources beyond the limits of those directly associated with production 0( a commodity.However, interdisciplinary effort is needed in the development of strategies, and societal support ofagriculture nlust take a Cormthat encourages and rewards agriculture Cor this stewardship.
INTRODUcrION
There is some truth to the view that something is wrong on planet Earth:its living systems are out of balance. Biological diversity is threatened.Natural habitat is being lost. Some predict a spate of extinctions unmatchedsince the end of the age of the dinosaurs (Table 1). And it's all the fault ofone species: Homo sapiens. We human beings are too numerous. We areputting too much pressure on the planet's biological support systems.
Nonetheless this perspective is over-simplified, blame-leveling, explainslittle, and offers no hope, save of drastically cutting back the humanpopulation, of reversing these mounting difficulties. Human numbers arepart of the problem, yet there are places, such as northern India, where aquite dense human population has coexisted with high biological diversity formillennia. Thus, at least part of the problem is due to human culture, nothuman population density per se.
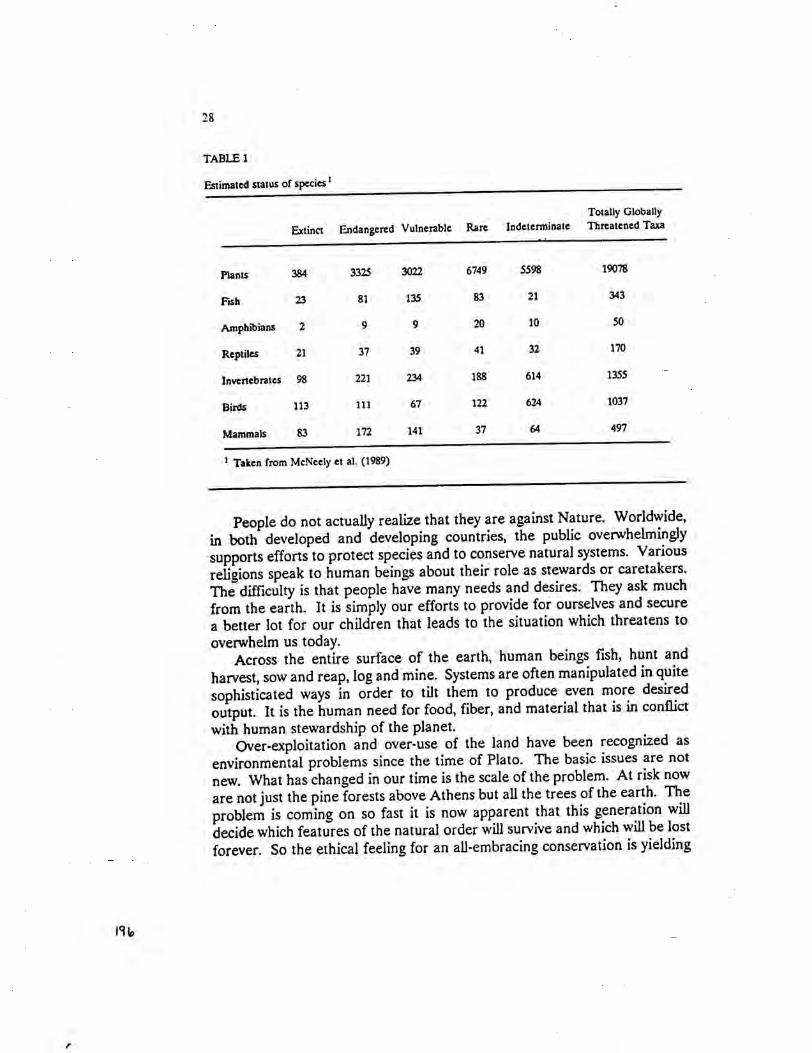
28
TABLE 1
~timated status of species 1
Totally Globally
Extinct Endangered Vulnerable Rare Indeterminate Threatened Taxa
Plants 384 3325 3022 6749 5598 19078
Fish 23 81 135 83 21 343
Amphibians 2 9 9 20 10 50
Reptiles 21 37 39 41 32 170
Invertebrates 98 221 234 188 614 1355
Birds 113 111 67 122 624 1037
Mammals 83 172 141 37 64 497
1 Taken from McNeely et al. (1989)
People do not actually realize that they are against Nature. Worldwide,in both developed and developing countries, the public overwhelminglysupports efforts to protect species and to conserve natural systems. Variousreligions speak to human beings about their role as stewards or caretakers.The difficulty is that people have many needs and desires. They ask muchfrom the earth. It is simply our efforts to provide for ourselves and securea better lot for our children that leads to the situation which threatens tooverwhelm us today.
Across the entire surface of the earth, human beings fish, hunt andharvest, sow and reap, log and mine. Systems are often manipulated in quitesophisticated ways in order to tilt them to produce even more desiredoutput. It is the human need for food, fiber, and material that is in conflictwith human stewardship of the planet.
Over-exploitation and over-use of the land have been recognized asenvironmental problems since the time of Plato. The basic issues are notnew. What has changed in our time is the scale of the problem. At risk noware not just the pine forests above Athens but all the trees of the earth. Theproblem is coming on so fast it is now apparent that this generation willdecide which features of the natural order will survive and which will be lostforever. So the ethical feeling for an all-embracing conservation is yielding
,

to the applied science of conservation biology, which pretends to offer ascientific basis for the choices we face.
This paper is an analysis, from a systems perspective, of the kind ofthinking the ecological sciences bring to these problems. Particular attentionis given to dimensions of time and space, and an attempt is made to showwhich processes dominate or control other processes.
CONSERVATION BIOLOGY
A decade ago, conservation biology had its birth in crisis. Manyvertebrate species were seen as teetering on the brink of extinction. Zoosbecame emergency rooms for species survival, with the genetic anddemographic systems of the patient the focus of clinical care. There are nowprocedures for handling these cases, yet only one species in ten thousand willever be selected for such treatment. The costs are too great, especially thecosts of physiological and natural history knowledge. Somehow, these at-riskspecies must be provided for in ways that demand less hands-on care.
Species are placed in zoos only when their population size in nature isno longer minimally effective. Species populations only drop to such lowsizes when the integrity of the processes of whole communities of species hasnot been preserved. And these ecological communities are threatened onlywhen there is a failure at the level of ecosystems and biogeographiclandscapes. Thus, mismanagement of the natural order can have ahierarchical character in which a mistake can compound itself throughsuccessive levels. This is the understanding which must guide the humanresponse to species risk. At question is not only how to act, but where andhow soon.
Conservation biology is now attempting to meet its challenges at thesehigher levels, involving land use and whole ecosystems. It is attemptingconstructively to engage the processes that threaten biological diversity. Todo this, conservation biology must draw on the knowledge that has beenaccumulated about these higher-level processes. To a large extent,conservation biology must simply reach out and borrow knowledge ratherthan create its own. In this sense, it is an eclectic discipline, However, thereare processes of "intermediate" size and duration that have not been wellresearched by academic ecology and about which critical knowledge iswanting. At these scales, conservation biology must add to the general storeof knowledge.
29
1~7

30
AORICULWRE
Agriculture, broadly construed to include fishing, forestry, and grazingsystems, is the dominant interface between human beings and nature. It isthe fundamental mechanism by which nature is manipulated. Agriculture isthe direct cause of the loss of natural habitat. When misguided, agriculturecan lead to both over-exploitation and the transport and introduction ofexotic species. And because it is central to the problem of threatenedbiological diversity, it is pivotal to the solution.
Agriculture can be divided into three types, graded by the intensity andexpense of the human inputs. First, requiring essentially no human inputs,are fisheries, grazing systems dominated by domestic animals, and forests notsubject to clear cutting and replanting. These systems can yield their productindefinitely if exploited properly. Second, there are the low input farmingsystems which, with proper management, might also be sustainable;developed-world forestry often fits in here. Third, there are the high inputsystems seen in the developed world where the system is dependent onhuman management and costly inputs of energy and chemicals.
Human beings can fail in the management of any of these three classesof systems and can rapidly degrade them to ecological uselessness. How thefailure happens, however, is different.
The zero-input exploitation systems are the most complex from abiological perspective and the simplest from a management perspective.They are all essentially one-dimensional. That is to say, to maximize theyield of lumber per year is the goal in forest exploitation systems; and thesteady-state stocking rate, in sheep equivalents, tells one what is mostimportant about grazing systems. These systems, since they involve specieswith long life-times and relatively slow growth rates, are slow to respond toand recover from management action and disturbance.
The other two classes of agriculture partially or wholly supplant thenatural landscape. They are, therefore, ecologically less complex and havefewer species. Because of human action on the landscape - plowing,flooding, seeding, harvesting or burning at the end of the growing season -its structure can change rapidly. That is, land that was in soybeans one yearcan be turned to corn the next year, with great consequence for animalsdependent on that system. Despite these features, agricultural systems aremore complex than natural systems from the management standpoint, sincethe farmer can control and manipulate them in a variety of different ways,such as selecting different soil treatment strategies, fertilizers, herbicides,pest control tactics, timing of activities, and product removal techniques.
,

31
Because the fundamental time scale of agriculture is the growing season,usually a year, many kinds of failures can quickly be corrected. Givensuboptimal performance from a system, a farmer can switch to a differentcultivar or application of herbicides in a subsequent season. Thus, modernagriculture has the appearance of a successful, progressive enterprise and,in fact, western agriculture is one of the most economically efficient systemsever invented by humans.
One example of a series of failures leading to success is seen in theBritish experience in Ceylon in the late nineteenth century. They firstattempted to grow coffee in their plantation system. However, the combinedattacks of Hemileia tastatrix and the native golunda rat wiped it out. Thenext step was to try cinchona, the source of quinine. The climate provedsuboptimal and the system was unable to compete with the Javan crop.Finally, they tried tea. This turned out to be a successful plantation crop.
When managers of high input agricultural systems try to sustain thesystem in a particular state for a long period of time, a feedback loop candevelop which requires more inputs with each passing year. This dependen-cy state is similar to the drug addiction into which a human being may fall.
Modern agriculture suffers from its own successes. Outside of Africa,the triumph of the green revolution has produced surpluses in mostcountries: mountains of butter, lakes of wine and milk. Until humanpopulation growth catches up with its food supply or distribution problemsare solved, such conditions offer a window of opportunity for adaptiveexperimentation.
One final category of the landscape consists of those areas of earth thatare now unproductive or otherwise degraded because of past mistakes inexploitation or agriculture. Degraded ecosystems are often quite simple,sometimes to the point of virtual sterility, and may have a permanence notunderstood by the public. The loss of soil or nutrients, the presence ofbiocides, or a dearth of proper colonists species may so constrain thesesystems that they must be lifted as it were, with great human effort andunderstanding, from the stable configuration of processes into which theyhave descended. Thus, these systems may have very long time scales ofrecovery.
BIOLOGICAL DIVERSITY
In a word, the goal of conservation is biological diversity. Yet biologicaldiversity is many different things. At different levels of biological organiza-tion, it involves features running from genes through species to ecosystems

2,00
,
32
and landscapes. Biological diversity can be measured over various spatialscales, and it may be quantified in a variety of ways.
At the level of species diversity, ecologists and biogeographers have threedistinct categories of measures: alpha diversity, the number of species at apoint in space; beta diversity, the rate of addition and subtraction of speciesas one moves along an environmental gradient; and gamma diversity, thedifference of species lists from site to site. We might extend this scheme andcall the total number of species on the planet or a continent omega diversity.It is important to understand that one can increase alpha diversity whilelowering gamma and omega diversity. It should not be the goal of conserva-tion biology to supplant rare endemics with a suite of otherwise commonspecies.
The biological significance of diversity among landscapes and ecosystemshas not been well quantified, though they are doubtless of great significance,at least aesthetically. One important biological aspect of a landscape ispatchiness. But it is often the species that operationally defines this orprovides our perception of patches.
Biological diversity is not stationary in time and space. There are ebbsand flows. The major features of these fluctuations are driven by changesin climate. Pollen studies in lakes of North America have shown how greatthe change of ecological communities has been since the Pleistoceneglaciation. And in Africa, which did not suffer glacial intrusion, the changehas been no less profound. The Kalihari and Sahara deserts were joined20,000 years ago and the tropical systems were squeezed onto a couple ofmountain ranges to the east and west. Lakes in the Rift Valley werehundreds of meters lower during the drought. This helps to explain thegreatly reduced beta diversity of the Zaire Basin compared to Amazonia.In the Zaire Basin, the species of plants and animals have extended theirrange a hundred-fold since the time of the great deserts, and have keptpretty much the same ecological structures through the range of thisexpansion. Amazonia has far greater heterogeneity from place to place.The origin of this is still uncertain.
Biological diversity and agricultural interactions
Although agriculture is concerned directly with only a small fraction ofthe genetic and biological diversity of the planet, as presently practiced itinfluences the entire biosphere. There is widespread agreement thatenvironmentally destructive and exploitative practices (with their increasingly

33
unacceptable societal and environmental costs) must be replaced by moresustainable food, fiber, and timber production systems.
To further complicate issues, this shift in agricultural practice will haveto be accomplished during a period of unprecedented change. First, thephysical environment is being transformed at a rate' hitherto unknown.Alterations of global climate and sea level will have a major impact on thefuture geographic distribution of agricultural activities. At the same time,the demand for agricultural products will likewise be changing at everincreasing rates.
If the primary imperative in the management of agricultural lands is theprovisioning of our own species, then in the next 20-30 years we will have todevelop sustainable agricultural systems capable of producing double or tripletoday's yields. This will be necessitated because in the coming decades, thehuman population will double and most tropical forests could be destroyed.These occurrences will further exacerbate both the human and agriculturalpredicaments.
Yet in this apparent conflict between agriculture and the environment,there are seeds of hope. Our need to produce more food, fiber, and forestryproducts is not necessarily incompatible with the conservation of biologicaldiversity. There is as much to be learned from the positive aspects of theinteraction between agriculture and nature as there is from the negativeaspects. Much is to be gained from examining agriculture's biological andhistorical roots. And there are many reasons to believe that futureagricultural systems can be designed to alleviate the problems encounteredthroughout the world today.
Traditionally, agriculture was totally dependent on nature. Naturalpopulations of plants and animals were exploited directly or, to varyingdegrees, domesticated. Future sustainable agricultural systems will likewisebe dependent on nature; the crucial linkage in the future, as in the past,involves genetics. Natural biological diversity does not stand outside ofagricultural diversity as it is broadly defined but, rather, is part of it. Andspatial and temporal dimensions are important considerations of this mix.
Agriculture is presently based on the genetic resources of a handful ofhighly select species and their wild relatives. The genetic dependence ofdomesticated plant and animal development on their wild relatives isincreasing as we recognize the wealth inherent in hitherto unincorporatednatural germplasm. With rapid progress in genetic engineering and theforeseeable ability to move genes between unrelated species, the genes ofwild plants and animals become the capital which must be banked orconserved if they are to return dividends in agriculture.
101

34
Agriculture and conservation biology are not mutually exclusiveenterprises. In fact, they can be linked positively on both local, regional, andglobal scales. Unfortunately, the populations and species that carry thegenes upon which future agriculture depends are rapidly disappearing justwhen the tools which can make effective use of them are being developed.This adds a great urgency to our attempts to shift "to more sustainableagricultural systems that are more compatible with biological conservation.
One of the most important ways agriculture can contribute to conserva-tion is through its legacy of research. Much of what we know about themanagement of plants and animals and their communities is a result ofagricultural research. Just as basic science promises to revolutionizeagriculture in the near future, so too the legacy of hundreds of years ofapplied research and experience by agriculture promises to contributepositively to the conservation of nature and natural resources. Agricultureand conservation biology should not be adversaries; each is important inbiosphere management. The human species is dependent on both disciplinesfor its prosperity and its future.
Natural biological diversity and agriculture interact dynamically, whatevertheir spatial configuration. Each directs outputs to, and receives inputs from,the other. In some cases, these input/output relationships may be betweenclosely adjacent landscape elements, but they may also have a far greaterspatial dimension. For example, at the short spatial range, the tropicalrainforests surrounding Gatun Lake in the middle of the-Panama Canal havebuffered the runoff of seasonal rains; their removal is causing more extremevariation resulting in water shortages during the dry season. At the greatestspatial reach is the general and gradual build up of carbon dioxide in theearth's atmosphere, which is produced in part by the burning of tropicalforests to clear land for agriculture and also by the consumption ofpetrochemicals necessary to supply and operate the system.
Neither natural biological diversity nor agriculture is neutral to the other.Concerning the damage spoken of above, it is important to remember thatsometimes one may benefit the other. Natural elements of biologicaldiversity within exploited and agricultural systems may yield a number ofbeneficial services to these systems. These have relevance to the vitalquestion of agricultural sustainability. Biological diversity may provide arefuge for biological control agents such as the predatory insects that preventoutbreaks of herbivorous pest insects.
Biological diversity stabilizes soils and prevents their loss. Biologicaldiversity regulates water flow and can also attenuate the effects of wind,thereby greatly lowering rates of erosion. Biological diversity generallybuffers disturbance and gives human beings greater time to deal with it.

35
Biological diversity also provides early warning of building toxicity, as thebirds of Rachel Carson's Silent Spring bore witness. At the genetic level,biological diversity is the source of novelty. The wild relatives of domesticcultivars have survived eons of change and challenge and have the answersto innumerable environmental problems stored in their genetic heritage,
Sources of ecological knowledge
Like other scientists, ecologists observe and compare. They accumulatedata on pattern and process. But their study systems span a spectacularrange of scale, from molecules to the biosphere. They know most at thegenetic and population levels, where there is more replication and wheretime scales are shorter. They know least at the level of the biosphere wherethere are a limited number of biomic types, but which have behaviors thatoccur over millennia.
Ecological experimentation in the laboratory is difficult and for fieldstudies, the attainment of the various statistical desiderata is often lacking.Even at small spatial scales, the replication of natural systems is often notcontrolled. Each island in an archipelago differs in numerous ways from theothers. For the student of bird diversity, for example, these differences canonly be ignored as noise or else factored out in the roughest statisticalmanner. All ecological knowledge of natural systems is subject to much thesame limitations.
Ecologists, like other scientists, build models. These models, however,are normally not intended as the basis for quantitative prediction. Rather,they summarize the categories of behavior that have been seen repeatedover many different systems. The models are .for logical prediction, nottemporal or spacial prediction.
The temporal and spatial scale of ecological knowledge acquisition hasbeen limited to the graduate student life-time because few systems have beenstudied in complimentary and overlapping fashion by teams of scientists.This one-person, one-question approach to ecological science producesstrong biases. Scientists want to study processes that behave fast and arespatially common because they need to harvest some novel publicationbefore their financial support dries up.
Some of the best knowledge in ecology comes from after-the-fact reviewsof major human disturbances and manipulations. These can be viewed asunwitting perturbation experiments. For example, the Smokey the Bearapproach to fire suppression in North America has revealed more about fire.ecology than ever could have been learned from the study of undisturbed

36
systems, that is, systems where fires had their pristine temporal and spatialpattern. There are many other such processes about which more and moreknowledge is being gained as human beings interfere with them. Theprospect of restoring degraded systems and working within agriculturalsystems to optimize biological diversity can certainly be seen as offeringunmatchable opportunities for the acquisition of new' knowledge.
The importance of diversity
Diversity can be measured at different levels of biological complexity(genetic diversity, species diversity, ecosystem diversity), all of which areimportant to the future of agriculture on the planet. The reader unfamiliarwith the basic arguments for the importance of conserving biological diversitymay want to consult volumes prepared by the U.S. Congress, Office ofTechnology Assessment (1987) and Wilson (1988). To see the importanceof the biological issues in national and international perspective refer toMyers (1984), Repetto (1985), and Western and Pearl (1989).
Genetic diversity involves the natural variability of the genes of individualorganisms and the collective variability of the genes of all members of apopulation, strain, race or species. A healthy population of a typical plantor animal species is characterized by genetic diversity; within it, there areindividuals of many different genetic constitutions (genotypes) (Table 2).This variety of genetic types is an indication that no single genotype is ideallysuited to all the conditions under which the population lives or all conditionsunder which the population might live. The variety of genotypes within thepopulation enables it to be successful over a range of ecological conditionsand to be able to adapt to changes in these conditions over time.
Animal and plant species used in agriculture typically have a muchnarrower genetic base. So much so that in species such as wheat, millionsof acres may be sown with a single homogeneous cultivar. This hasconsiderable advantages under the highly standardized conditions offertilization, growing season, pesticide application, and harvest methods ofmodern agriculture. But it has enormous disadvantages if a virulentpathogen evolves to attack the population because all individuals in thatpopulation are vulnerable. Similarly, a lack of genetic variability willdiminish a population's ability to survive normal climatic variations.
This reduced ability to adapt or evolve makes the conservation of withinspecies genetic variability of fundamental importance to future agriculturalpractices. For a more detailed discussion of the term "genetic conservation"as it applies to actions at this level of biological conservation, the reader is

37
TABLE 2
Levels of hetcrozygosity (H) and proportion of polymorphic: loci (P) (or various taxa I
Hetcrozygosity Polymorphism .'
species mean s.d. species mean s.d.
Venebrala 551 0.054 0.059 596 0.226 0.146
Invencbrata 361 0.100 0.091 371 0.375 0.219
Plants 56 0.075 0.069 75 0.295 0.251
t Summarized from Nevo (1984).
referred to the recent reviews by Frankel and Soule (1980), Oldfield (1989),and Orians et al. (1990). Brown et al. (1989) contains numerous discussionsof the application of population genetics in forestry and crop improvement,and accounts of the problems and practices of conserving species leveldiversity are given by Ehrlich and Ehrlich (1981), Schon ewald-Cox et al.(1983). Wilson (1988), Woodruff (1989). and Fiedler and Jain (1991). Everysingle. species-level extinction diminishes the options and prospects forhumane management of the biosphere.
THE BIOSPHERE
The scope of interest in conservation biology is immense. The field ofstudy is best thought of as a great hierarchy of entities and processes (Fig.1). Within the complete organizational structure of nature, the principalfocus of integrating conservation biology and agricultural practices rangesfrom the larger landscape ecosystem down to the individual organism. Allof this is contained within the planet Earth and is dependent on its stability.
The biosphere, the 30 kilometer thick zone of biological activity sur-rounding the planet, is the environment for life on earth. In the biosphere.energy balances are crucial for the continuity of its processes. It seemsstrange to some scientists that the temperatures on the earth's surface haveremained in a relative tight range for the last three billion years, neitherfreezing nor boiling life before it had the chance to produce the species thatbecame self-conscious of this surprising stability. This global homeostasis isremarkable because the sun has changed its solar flux by as much as 30%over this period of time. Some scientists believe that the biosphere has self-

38
W!!.lliI I I
IEARTH
ATMOSPHERE GEOSPHERE BIOSPHERE HYDROSPHERE
BIOSPHERE
I I1 ... > N
BIOMES
1 23 •••••••••••••• _> nECOSYSTEMS
COMMUN ITIES
I IQUILDS
IPOPULATIONS
, 23 •••••••••• > nINDIVIDUALS
I IORGANS
I IFig. 1. Hierarchical structure of conservation biology. Adapted from Western and Pearl (1989).
regulatory properties (the Gaia hypothesis) with which it can accommodatedisturbance.
Two properties largely control the capture of solar energy by our planet:the composition of the atmosphere and the reflectance of the surface of theearth. An atmosphere with elevated levels of carbon dioxide absorbsreflected infrared radiation and produces a greenhouse effect. A whitersurface - ice or sand - reflects heat and cools the planet.
Global modelers are now attempting mathematically to mimic theseprocesses at this ultimate spatial scale. To do so, they need to understandthe inputs from ecological and agricultural processes. The output from sucha model will predict the climate of the future and thereby point to whatchanges ecological and agricultural systems will need to make in response.
Unfortunately, the scales of knowledge in these enterprises are different.The unit in a climatological model might be 300 to 400 square kilometers,a size dictated by the capacity of computers available. This is considerablylarger than units normally studied by ecologists - so processes that occur
,

39
at such a scale will be referred to as "mesoscale" processes; their study iscrucial for linking ecology to climatology.
If climate changes equal anything like some of the predictions now beingadvanced - e.g. a 9°e increase in average temperature and a 50% drop inrainfall in the North American Midwest - there will be massive displace-ment of systems to different locations on the earth's surface wheretemperature and rainfall patterns are more conducive to their survival. Atthis point in time, no one can identify these locations. Today's storehousesof diversity may be tomorrow's deserts.
Populations and species
The new applied science called conservation biology seeks to integratethe findings of traditional agriculture and resource management with recentadvances in population genetics and ecology. In conservation biology,traditional population concepts are typically replaced by metapopulationmodels, and population structure and subdivision are recognized as havingcrucial effects on demography.
A local population of a species is typically at equilibrium with itssurroundings when it is at a level that its total density cannot exceed. Whenthe number of individuals falls below this level, there-is an average per .capltarate of growth. But all local populations receive from their environment,random shocks of varying intensity, and these stochastic inputs subject allpopulations to a probability of extinction. Only by being part of a gridworkof similar local populations, from which immigrants and colonists can arrive,can a population persist for an appreciable length of time. The collection oflocal populations over a gridwork of environmental patches is called ametapopulation.
Both local populations and metapopulations have thresholds of size andspatial arrangement below which they suffer catastrophic increases in theirprobability of extinction. Because local populations under such conditionsare not indefinitely elastic, at some point they snap quickly to extinction.Many such cases have been studied with a variety of techniques, and semi-quantitative predictions can be made of where these catastrophic thresholds- involving the joint action of genetics, demography, and spatial arrange-ment - are likely to be crossed. The thresholds are different for differentspecies and, unfortunately, there are no magic minimum viable populationnumbers that apply equally to all cases.
Local extinction.of components of a metapopulation are now viewed asnormal occurrences. The field of population vulnerability analysis (PYA)

40
focuses on the relative significance of genetic, demographic, and habitatfragmentation processes in causing such extinctions. Increasingly, predictionsabout the survivability of populations must be made with reference to aspecific time scale: single generation, ten generations, a hundred genera-tions, a thousand generations. Proposals to manage -populations for veryshort-term gain can have exceedingly deleterious long-term consequences.
In practice, agriculturalists conserve populations and species rather thanspecific genes of economic importance. In addition, there is widespreadrecognition of the need to protect many domesticated varieties and breedsand other threatened natural species of great potential value to forestry andmedicine.
Agriculturalists should be aware that conservation and evolutionarybiologists occasionally employ a different taxonomy for the population unitsof concern. Agriculturalists have traditionally recognized wild species,subspecies, land races, breeding lines, varieties, stocks, strains, and cultivars.Recently, genetically engineered lines have been added to this lexicon. Incontrast, conservation biologists recognize demographic and evolutionaryunits - clones, outbreeding populations and species. The traditionalconcepts of the wild-type genotype and of the subspecies as an evolutionarycategory have been progressively abandoned. Biological species areenvisioned as groups of populations that share a genetic and evolutionarycohesion based on the ability of individuals to discriminate between membersof their own species and members of other species. Such evolutionaryconcepts are directly applicable to agricultural situations.
The difficulty in implementing conservation programs is a consequenceof the number of species involved. There are presently 30,000 threatenedplant species and an additional 10,000 plant species of potential economicvalue that may require u situ conservation unless suitable in situ conserva-tion measures are developed. The National Marine Fisheries Service(U.S.A.) has implemented 30 management plans in a move towardconserving stocks of 300 species of fish, crustaceans, and molluscs, themajority of which are still being overfished.
At the international level, conservation efforts for a few hundredespecially appealing animal species have been organized by such non-government organizations as the I.U.C.N. (Species Survival Commission with100 specialist groups) and the American Association of Zoological Parks andAquariums (40 Species Survival Plans).
The number of animal and plant populations and species whose survivaldepends on intervent.lve management far exceeds present levels of resourcecommitment. Woodruff (1989) has reviewed the problems of conserving
'lo'i3

41
animal genes and species noting the strengths and weaknesses of existingmanagement plans.
Communities
Within the mesoscale regions of the earth, there are smaller, more easilyunderstood units. Communities are objects of study in which speciesidentities are known and in which the densities of their members may bedetermined. In fact, sometimes the community is represented as a set ofequations of motion for the densities of the individual species populations.Yet communities have many emergent properties that are also the properstudy of community ecology.
The web of feeding relationships is one area of active study. Feeding isnot random but has a structure along which members of the community canbe arrayed in sequential fashion - feeding patterns tend to form discrete"intervals" rather than overlapping on their prey species in a randomarrangement.
Alternative and mutually exclusive collections and arrangements ofspecies may be possible at any point in space. That is, the character of thecommunity is not completely set by extrinsic factors. The forces generatedbetween the species themselves_can_haveoverridingimportance. The historyof the system is also important. Shifts between communities can be verysudden, while the time spent in such "domains of attraction" may be quitelong.
Communities are networks of balanced forces. Though their appearancemay be static, they are fully dynamic. When a force is applied to one speciesor one.component in a community, actions and interactions spread to theother components through many direct and indirect linkages. Often the finalconsequence of the removal of one species on the condition of anotherspecies I may be different from what would be predicted from cursoryexamination of the system. For example, the removal of one competitor canlead to]the extinction, not the increase, of another competitor.
There is another consideration that leads to the conclusion that theprediction of community dynamics will be difficult. Outside of their stableconfiguration (if one exists), communities are likely to be governed bynonlinear relationships. This makes communities very sensitive to smallchanges in conditions; trajectories of community behavior that are initiallyquite similar can diverge to very different final behaviors.
All communities have species components that provide structure for thesystem. These are sometimes called "keystone" species. That is, the great

42
majority of the species in the community adjust their densities and spatialdistributions to features of the keystone species. The keystone speciesprovide scope and stability to other species, but not conversely. Thecommunication of information and adjustment among species is typicallyrapid. In most cases this allows ecologists to ignore .the presence of non-keystone species and to focus their research and dynamic modelling on thefew, key species.
Ecosystems and landscapes
Ecosystems differ from communities most importantly in the way theyare perceived by the scientists who study them. Simultaneous equations ofmotion for hundreds and thousands of species populations cannot be studiedprofitably. Consequently, ecologists lump, or aggregate the individualcomponents, and knowledge of community processes is used as a guide tohow this aggregating should be carried out.
Ecosystem ecologists use the language of systems analysis common to allscience where complexity is a fundamental reality. Almost indispensabletools provided by systems analysis are those allowing for control andmonltoring.of.Inputs and outputs, stresses and responses. There is.very littlethat can be learned by simply observing the steady state values of a systemat equilibrium.
The characterization of complex behavior is necessarily subjective. Onetries for a simplicity that is consistent with an accurate description ofbehavior; the description is adjusted when confronted with surprisingbehavior. Unfortunately, all too often the behavior of ecosystems isunexpected, especially when the ecosystem is stressed from external activitiesassociated with exploitation and agriculture. However, much is learned fromsuch confrontations and the knowledge gained will guide the management ofsimilar situations in the future.
The equilibria I, balance-of-nature view of ecosystems under which anyshock could be absorbed, and any stress accommodated, is still commonlyheld and often implicitly accepted by society and its managers. For example,if the river system has been able to absorb an amount X of sewage, then itcan absorb amount 2X. Another manifestation of this view is that societiesoften take spatial scale as irrelevant since the same kind of equilibriumobtains everywhere. This view has been proved false in virtually everysituation where it has been tested experimentally.
The new paradigm of ecosystem structure recognizes the existence ofmultiple stable states, and it emphasizes variability and spatial heterogeneity.
'J..IO

43
Behavior is only expected to be continuous over small domains of environ-mental variation. Sharp, discontinuous adjustments are expected at the endsof these ranges. Most experience with this new paradigm has come fromsystems where human beings have played a great role.
Consider, for the moment, changes in climate. Such changes have beenprofound even before human modification of biochemical cycles began toplay a role. The second of the two views of systems dynamics mentionedabove demands a simple management prediction: indefinite, slow adjust-ments in agricultural practices cannot be expected, even under gradualmodification of climate - at some point, very abrupt adjustments will haveto be made. It is precisely at these points where a large pool of alternativecultivars and management strategies are necessary if the changes are to besurvived with minimum disruption to support processes.
Considering a different level of spatial perception, the instability justdiscussed can become the basis for regional stability. Many stable systemsdepend on a random pattern of small spatial scale unstable transitions. Eachsmall spatial patch in the system may take on a variety of behaviors over along period of time. But only by playing such different roles over time canthe patch be properly renewed and refreshed to continue in the game. Forceit into anyone role for too long and it will eventually collapse to somedegraded form in which it may remain indefinitely.
MIGRATION AND DISPERSAL
The movement of animals and plants over large distances is a processthat "cuts through" and interacts with all the "horizontal" layers of organiza-tion discussed above, from genes to ecosystems, and must be consideredseparately.
Organisms move for a variety of reasons. Often they move back andforth over a set route to take advantage of seasonal patterns of foodavailability; this is called migration. Large-scale migration is seen in suchvarious organisms as butterflies, birds, whales, and savanna grazers. Asecond kind of movement is permanent, within the lifetime of the individualorganism; it is called dispersal. Typically, one sex and age class will leave theplace of its origin to settle elsewhere. The evolutionary explanation for thisis that genetic lines survive because they have 'hedged their bet' on long-termsurvival by playing in many different and thus uncorrelated environments.This reduces the risk of 'betting' everything on the constancy of theenvironment at orte point in space. Another possibility is that genetic lines
2.1\

'2.1'2..
,
44
with the 'urge' to move are successful because this behavior balances someof the negative values of inherent small population sizes.
Migration presents a management problem at the level of individualspecies, because it means that the chain of survival is only as strong as itsweakest spatial link. Over-harvesting, pollution assaults, or habitat loss atanyone of the areas of a species' occupancy can doom the entire system atall its locations. Migration also can contribute to dispersal in the sense thatseasonal movement tends to "mix"individuals from various origins within thepopulation. For example, the monarch butterfly and whooping crane requireprotection by different national governments.
When the migrating species plays a keystone role in one of the spatialsystems of which it is a part, the management becomes quite uncertain andrisky. The predatory activity of birds on the larvae of herbivorous insectscan often be a key environmental process, as it is, for example, in the forestsof Canada. The birds suppress the population growth of herbivorous insectsand keep their numbers in balance with regard to food supply, i.e. the forestitself. However, these birds migrate to tropical areas in Central and SouthAmerica to over-winter. With accelerating deforestation in the tropicalareas, there is increased risk of defoliation to Canadian coniferous forests.The bulldozer in Bolivia threatens economic and ecological disaster inCanada (Terborgh, 1990).
SMALL POPULA nONS
Many populations of agricultural importance are either numerically smalland geographically isolated or numerically large but derived from very fewfounding individuals. Both types of populations are effectively small in agenetic sense. The genetic behavior of effectively small populations isqualitatively different from that of large out-breeding and geographicallywidespread populations, and more active intervention is required for theirmanagement.
The genetic vulnerability of rare or threatened species is often a directconsequence of habitat destruction and range fragmentation. The eventualloss of the species may be due either directly to the lack of adequate habitator indirectly due to genetic consequences of range fragmentation andreduction in population size (demographic bottlenecks). The analogiesbetween natural species and agricultural strains or cultivars are notaccidental, from a management perspective; both are based on the same setof genetic principals.

45
Inbreeding - the mating of closely related individuals - is rarely aproblem in nature. But, under some management regimes, it can effectivelycripple small populations. Outcrossing populations that suddenly decline innumbers usually experience reduced viability and fecundity known asinbreeding depression. Inbreeding produces increased homozygosity ofrecessive deleterious mutants and, by chance in small populations, thesealleles become fixed. Breeding programs for managed populations must bedesigned to counter the effects of exposing lethal and deleterious recessivegenes in homozygous form.
In large natural populations, close inbreeding rates are typically less than2% per generation and over centuries of experience animal breeders havelearned to keep the rate at approximately half that level. The consequencesof closer inbreeding are tragically illustrated in isolated human populationsor in mammals in zoos where juvenile mortality is greatly increased foroffspring of related rather than unrelated parents.
In contrast, management involving gradual inbreeding or reduction innumbers allows selection to purge deleterious recessive alleles as theybecome homozygous. These populations, and those of species that normallyreproduce by self-fertilization, suffer little inbreeding depression. However,the hybrid vigor seen when inbred lines are crossed shows that they still carrysome genes with poor performance when in the homozygous state. This isa continuing problem for managers of agriculturally important populations.
The second major problem associated with small populations involvestheir inexorable loss of genetic variability. In small populations, randomfluctuations in allele frequencies lead to the occasional loss of rarer allelesand the fixation of commoner ones. Such generation-to-generation fluctua-tions are the result of chance: different individuals play out their lives withor without reproducing successfully. Such genetic drift, as it is knowntechnically, reduces genetic variation and leads to increased homozygosityand loss of ability to evolve.
A fundamental concept is that effective population size, N., is closelylinked to this principle of genetic drift. Sewall Wright's notion of effectivepopulation size is somewhat awkwardly defined as the number of individualsrequired in an ideal population that would be expected to experience geneticdrift at the same rate as the actual population. Unequal numbers of malesand females, increased variance in family size, and temporal fluctuations innumbers all cause effective population size to be much less than the actualcensus count simply because not all individuals contribute equally to futuregenerations. In the absence of factors promoting genetic variation, such asmutation and dispersal, the expected rate of loss of heterozygosity is1/(2N. + 1) per generation. The same rate of loss is predicted for the

46
variance in polygenic characters, many of which are of agricultural impor-tance. Simply stated, loss of variability increases as the genetic effectivenessof the population of breeding individuals decreases. The survival of twoindividuals would produce progeny with only 25% of the original variation.Although little genetic variation may be lost in anyone generation, smallnumbers sustained for several generations can severely deplete variability-most variability should be lost within 2N. generations. When thesetheory-based relationships are translated back into real numbers it becomesclear that many current management practices strip populations of theirvariability within relatively few generations. Thus, genetic drift can imperila mismanaged population within tens rather than hundreds of generations.
During the last decade there has been considerable debate about theoptimal values of N. for a managed population. For outcrossing naturalpopulations, two values, N. = 50 and N. = 500, have been proposed based ontheoretical considerations and different management objectives (Frankel andSoule, 1981). Studies of mutation rates in a few polygenic characters suggestthat a population with N. =. 500 should maintain its intrinsic geneticvariability indefinitely. Similarly, it is argued that N. = 50 was a require-ment for short-term survival. -
Definitions of minimum viable population size based on such magicnumbers are now seen as unsound (Lande, 1988). The numbers based onthe theoretical consideration of a few characteristics of the organism may notbe relevant to the behavior of the whole genome. Genetic uniformitarianismcannot be assumed; other types of genetic variation are equally important inestimating minimum viable population size. Thus, there is no simple answerto the optimal effective population size question other than the trivial onethat large is better than small. Clearly, a reduction in the degree to whichpopulations are geographically isolated would effectively increase populationsize. Thus, an integration of agricultural practices and biological resourcemanagement strategies offers great possibility. The agricultural landscapecould be the largest nature preserve human beings ever dreamed of.
EXPLOITATION STRESS
This section addresses explanations for the degradation and collapse ofexploitation systems, whether fisheries, forests, or grazing systems. Thesesystems are one-dimensional from the standpoint of the manager in thesense, there is a single product to be maximized. The product may not admitinternal variation, but there is usually a linear scaling factor between theseinternal classes. For example, an old-growth douglas fir may be twice as
,

47
valuable per unit volume as a six year-old tree. And a skipjack tuna may beworth 1.3 yellowtail tunas.
Managers of such systems invariably adapt a method of maximumsustainable yield. One thing managers are especially adverse to is variationin output. Typically, they have a capital investment in extraction equipmenton which they have to pay interest, a constant per year amount. Disasterlooms if there are years or seasons where the yield, in monetary terms, isbelow the carrying costs for capital. Therefore, attempts are made tohomogenize the behavior of the system in time. This does not necessarilyrequire spatial homogenization, but it often leads inexorably to this state.
The Sahel is the semiarid region between the hyper-arid Sahara Desertand the regions of the African Savanna. Over the last 40 years, a series ofefforts has been undertaken to make it more productive in the hope offeeding the growing human populations in the region. Prior to World WarII, most of the region was engaged in migration-based pastoralism involvingcattle. After the war, peanut farming spread north into the drier regions,displacing pastoralists. With the elimination of cattle in these farmingregions, natural fertilization of the food crops ended. The farmers were toopoor to supply artificial fertilizers so yields started to decline.
A second homogenization process contributed to the problem. Thepastoralists who inhabited the driest regions of the Sahel were forced tomove more often than normal because of a lack of groundwater. Politiciansthought it better to have them stay in a given place so wells were drilled toprovide permanent water. This provided better health services to bothhumans and cattle, causing population growth. But the elimination ofseasonal migration meant that the grassland surrounding the permanentwater was soon overexploited.
A train of ecological consequences was set running on a track towarddisaster. Changed grazing patterns led to a succession of plant speciesfavoring shallow-rooted and unpalatable forms. More soil was exposed, andthere was compaction from human and domestic animal activity makingconditions unfavorable for plant establishment. A process of decertificationwas established.
This decertification is apparently changing local weather for the worse.Sandy, denuded soils reflect a greater fraction of solar energy than do soilscovered with vegetation. Given that local rainfall is produced by the heatingand lifting of moist tropical air, the cooler soil conditions mean even lessrainfall, which only contributes further to decertification. A management-induced, self-sustaining drought has degraded what once was a stable systembased on a migratory pattern of grazing.
~J5

48
There is possibly no way out of this cycle short of massive intervention.Small spatial solutions are worthless, for as soon as effort to create them isrelaxed, the area is again engulfed by the regional equilibrium. Only aregion-wide solution will work. Some propose that the humans and thegrazers would have to be removed from the region that spans 10 differentcountries and then the soil would have to be artificially fertilized so that itwould turn green after periods of greater than average precipitation.
Similar stories have occurred in fisheries systems. The great whales,although not totally eliminated, have been pushed to such low populationdensities that their per capita growth rate is reduced to near zero. Part ofthe problem no doubt arises because of difficulties finding mates, which isassociated with uneven age structures and reduced rates of social encounter.The dungeness crab population off the shore of Northern California waspushed low enough that an egg parasite, which had previously been swampedby the crab's reproductive output, is now able to hold the crab stably at thislow level.
Overexploitation of multispecies fisheries often leads to the eliminationof all but one of the species. This is the direct consequence of thecelebrated "principle of competitive exclusion" that says two species cannotbe limited by the same factor. Since exploitation by humans becomes thedominant limiting factor -for all the-species, only the species best able torespond to this pressure will survive. This is the reason that in denselypopulated regions of the earth, carp is the common table fish.
DEPENDENCY
A human culture can become "locked-in" to a particular form ofexploitation. We call this dependency. Among other things, it means thatthe human culture is part of a stable ecological system and that any changeof human behavior could produce profound changes in the non-humansystem processes, or vice versa. The Irish peasants of the eighteenth andnineteenth centuries became dependent on the potato. They were notinitially dependent on the potato, but their life style, demographic patterns,political and economic systems, and land tenure practices adjusted to thebounty of the potato. They were part of a system that only a disaster ofmajor proportions seemed able to change.
The nomads of the Sahel were culturally dependent on a wanderingpastoralism. But; at least in the short run, this dependency was mainlycultural so that external political forces could greatly alter this pattern. The

49
system is now in a transition phase and what equilibrium it is tending towardis not yet clear.
Developed world farmers are often dependent on a particular agriculturalsystem. This dependency is relative and not the product of a lack ofecological knowledge. In fact, it may be the consequence of an assumptionthat we understand more than we think we do about ecological processes.
Many classes of developed world farmers are locked into modes ofproduction because of a great investment in capital. The consequent need topay the interest costs of this investment on a timely basis results in mostmanagement decisions being made at the margin, so to speak. Thesefarmers are so deeply committed to a practice that the extra cost of a late-season pesticide application is minor and taken as a matter of course.
This economic dependency is in many ways no better or worse than otherforms of dependency - but in one quite foreseeable way it is probablyunsustainable. If this dependency were true and the economic constraintsvalid, there might be no problem. But through governmental policies tosupport agriculture, these constraints are often fictional and deny trueeconomic constraints that ultimately must be felt.
Petroleum inputs are one obvious case. The cost of petroleum-basedfertilizers and herbicides, and also the energy inputs to farm machinery andtransportation, represent only extraction costs of petroleum, not its trueworth. Petroleum extraction rates will soon start to decline and the price ofthese resources to farmers will greatly increase, making their applicationunprofitable. What replacement will there be for them?
Water is another commodity that is often not costed properly. The SanJoaquin Valley of California, especially the west side, depends on irrigationwater that has to be pumped uphill. The laws of thermodynamics being whatthey are, this consumes the bulk of the energy that the same water earlierproduced as it dropped through the turbines of great dams. With risingenergy prices, the public will soon have to face choices on how it consumesits hydroelectric power.
There are, however, other costs associated with irrigation. Irrigationinvariably leads to the build-up of salts in the soils. The fall of civilizationshas been based on this simple fact. To forestall the accumulation of salts inthe soil, farmers must apply additional water that carries off salts through adrainage system. This waste water must be collected and disposed. In 1957,the State of California came to grips with this problem and drew up a planto transport such waste water north to the Sacramento River for disposaLIn 1960,Congress expanded the project to encompass a 300 kilometer masterdrain for the wh~le valley. In 1964, however, a consultant predicted the

50
"total destruction of wildlife" should the waters from this drain reach the SanFrancisco Bay.
Construction on the drain began in 1970. The intention was to dischargethe waste water into the Sacramento-San Joaquin River Delta, but areservoir was needed to regulate the flow. Land was purchased in westernMerced County that became the Kesterson Reservolc.Because this was alsosensitive wildlife habitat, the U.S. Fish and Wildlife Service agreed toparticipate in the management of the drainage water.
What has happened since is unmitigated disaster. The connection of thedrainage canal to the San Francisco Bay was never completed. KestersonReservoir became a progressively more poisonous evaporation pond, and theFish and Wildlife Service now must try to keep wildlife out of it. Nosolution is yet in view.
HUMAN SCALES AND HUMAN RESPONSE
In discussing the theory of scale, it is important to recognize the roleplayed by human beings in natural and agricultural systems. Various humancultural forms are scattered in a mosaic all over the earth. Different cultureshave different time horizons in viewing nature and formulating political andmanagement plans. With pressing mortgage payments, a farmer may not beable to see past the current growing season. The politicians of democraciesoften cannot see past the next election. Cultural patience may depend onreligion. Patterns of land tenure are extremely variable. The Japanesehorticulturalist may make a living off a few hectares, while the Texas ranchermay control an area of thousands of square kilometers.
The Irish potato famine of 1845-1848 speaks to temporal scale issues inhuman adjustment. The potato was brought to Europe in the early 1600'sby Sir Walter Raleigh. Over the next two and one-half centuries it grew topervasive popularity among the Irish peasantry for a variety of reasons,including: it was well-suited to the deep soil of Ireland; it was not particularlysensitive to war (of which there were many during this period) sincetrampling and burning could only destroy the above-ground part of the plant;it required a low input of external materials and was, thus, suited to poorpeople; its nutrients were complimentary to those produced by dairy cattle,making a baked potato with sour cream a complete meal; it was even thesource of an alcoholic intoxicant, poteen.
The Irish peasants adjusted slowly to the blessings of this highlyproductive crop by increasing their population growth rate, typically through

51
early marriage. Politically, they divided their land holdings to roughly ahectare per family.
Spatially, however, Ireland was far from homogeneous. A great deal ofthe land in Ireland was held by absentee English landlords who practicedcash-crop agriculture and pasturage. There was also a group of Monteneginsfrom Serbia who practiced a very different agriculture. Both of thesecomponents of the system were perfectly unaffected by the potato famine.
The potatoes of Ireland, and all Europe, had gone through a smallgenetic bottleneck during their introduction and were geneticallydepauperate. In 1845, a blight struck and the crop failed. There was wide-spread hunger. "Bad luck" the peasants thought, and replanted potatoes thenext year. Again, the blight struck. There was starvation. But again theyreplanted potatoes.
They were spared the blight in 1847, by which point they had made verylittle adjustment, but it returned in 1848. This exceeded the limit of theresiliency of the system. There was great starvation and mass emigration toAmerica; 50% of the population died or emigrated. Finally, the episodegreatly altered the subsequent social and political structure of Ireland. Landholdings were consolidated and passed to the eldest son. The populationgrowth rate was stabilized, principally through delayed marriage and a slowrate of emigration. The population level stabilized at 50% of its mid-nineteenth century-level.
While the adjustment of the Irish has proven to be permanent, it is notcomforting that it took so violent a disaster to achieve it. The greatproblem, which is outside the province of ecological theory, is how tomobilize an adaptive response by a society before a disaster strikes.Understanding ecological processes is important, but understanding is notsufficient to bring about change.
REFERENCES
Brown, A.D.H., Oegg. M.T., Kahler, A.L and Weir, B.S. (Editors), 1989. Plant Population Genetics,Breeding. and Genetic Resources. Sinauer Assoc., Sunderland, MA.
Ehrlich, P.R. and Ehrlich, A.H., 1981. Extinclion: The Causes and Consequences of the Disappearance 0(Species. Random House, NY.
Fiedler, P.L and Jain, S.K., 1991. Conservalion Biology. The Theory and Practice of Nature Conservation,Preservation and Mana,ement. Chapman and Hall.
Frankel, O.H. and Soule, M.E., 1981. Conservalion and EVOlution.Cambridge University Press, NY.Lande, R., 1988. Genetics and demography in biological conservalion. Science, 241: 1455-1460.McNeely, JA, Miller, K.R., Reid, W.V., Millermeier, R.A. and Werner, T.B., 1990. ConservinC the
World's Biological Dive~i!y. JUCN, Gland, Switzerland; WRl, CI, WWF-US, and the World Bank,Washington, DC.
Myers, N., 1984.The Primary Source: Tropical Forests and Our Future. Nonon, NY.

52
Neva, E., 1984. Patlems of genetic: variation in natural populations. In: 0.5. Mani (Editor), Evolutiona,yDynamics of Genetic Diversity. Springer-Verlag, NY.
Oldfield, M., 1989. Conservation of Genetic Resources. University of Washington, Seattle.Orians, G.H. (Editor), 1990. Conservation of Genetic Resources. University of Washington Press, Seattle,
WArRepetto, R. (Editor). 1985. The Global Possible. Yale University Press, New Haven, cr.Sc:honewald-Cox, e., Chambers, S.M., MacBryde, B. and Thomas, L. (Editors), 1983. Genetics and
Conservation. A Reference for Managing Wild Animal and Plant Populations. Benjamin/Cummings.Menlo Park, CA.
Soul~, M.E. (Editor), 1986. Conservation Biology. Sinauer Assoc., Sunderland, MA.Soul~, M.E. (Editor), 1987. Viable Populations for Conservation. Cambridge University Press, I''Y.Terborgh, J., 1990. Where Have All the Birds Gone? Princeton University Press, Princeton, NJ.U.S. Congress, Officc o( Technology Assessment, 1987. Technologies to Maintain BiolOgical Diversity.
OTA-F-330. U.S. Govemment Printing Office, Washington, De.Westem, D. and Pearl, M.e. (Editors), 1989. Conservation (or the Twenty-First Century. Oxford
University Press, NY.Wilson, E.O. (Editor), 1988. Biodiversity. National Academy Press, Washington, De.Woodruff, 0.5., 1989. The problems of conserving gcnes and species. In: D. Western and M.e. Pearl
(Editors), Conservation (or the Twenty-First Century. Oxford University Press, NY, pp. 76-88.
1.1.0


















