
1 Self-Replication versus Self-Elongation Or: How to make long oligonucleotides without enzymes,...
23
1 Self-Replication versus Self- Self-Replication versus Self- Elongation Elongation Or: Or: How to make long oligonucleotides without How to make long oligonucleotides without enzymes, primers, templates, surfaces, or enzymes, primers, templates, surfaces, or stepwise feeding? stepwise feeding? Oliver Thoennessen, Mathias Scheffler & G. von Kiedrowski, Ruhr-University Bochum d COST D27 workshop, Heraklion, Crete, Sept. 30-Oct. 3, 2004
-
date post
22-Dec-2015 -
Category
Documents
-
view
216 -
download
0
Transcript of 1 Self-Replication versus Self-Elongation Or: How to make long oligonucleotides without enzymes,...

1
Self-Replication versus Self-ElongationSelf-Replication versus Self-Elongation
Or:Or:How to make long oligonucleotides without How to make long oligonucleotides without enzymes, primers, templates, surfaces, or enzymes, primers, templates, surfaces, or
stepwise feeding?stepwise feeding?
Oliver Thoennessen, Mathias Scheffler & G. von Kiedrowski, Ruhr-University Bochum
3rd COST D27 workshop, Heraklion, Crete, Sept. 30-Oct. 3, 2004

2
The "standard" pictureThe "standard" picture
Who agrees?
1. Self-Replication
2. Metabolism
3. Mutability
4. Some way of keeping 1-3 connected, viz. compartimentation

3
Chemical self-replicationChemical self-replication
+
katalysierteLigation
Assoziation Dissoziation
ABC C2
CCA B
spontane Ligation
ka
kb

4
Open systems, possible non-catalyzed pathwaysOpen systems, possible non-catalyzed pathways
+2 3 4
+ + + +
x 2
x 2
432 5 61
building blocks for "closed" systems,single-sided reactivivity
building blocks for "open" systems,dual-sided reactivity
templates,non-reactive
complementary
self-complementary

5
Open systems: possible template-directed pathwaysOpen systems: possible template-directed pathways
+ + + + n

6
OP
O
O
ON
N
N
N
NH2
O
O
P
O
O
O
ONH2 N
N
O
NH2
OP
O
O
O
OO N
N
N
NH
O
NH2
P
O
O
O
ONH2 N
N
O
NH2
OP
O
O
O
OO N
N
N
NH
O
NH2
P
O
O
O
ONH2 N
NH
O
O
CH3
OP
O
O
ON
N
N
N
O
O
P
O
O
O
ONH2 N
NH
O
O
CH3
NH2
PO42-NH2 CA
bzw.CAn p
bzw.TGn p
PO42-NH2TG
bzw.TAn p
PO42-NH2TA
bzw.CGn p
PO42-NH2 CG
T G
C G
C A
T A
DimerDimerbuilding blocksbuilding blocks
for an open for an open system:system:
nYRpnYRp

7
Dimer synthesisDimer synthesis
OH
OBOH
O
OP
O
O
OB'2OH
Cl S
B'1
O
OP
O
O
O
Cl
N3
P
O
O
OB2OP
O
O
OB1N3
O
S
O
O
N+
O
OMe SO2ClH
+NEt3
B1 B2
C A C G T A T G
OP
O
O
OB2O
O
P
O
O
O
OB1NH2
OP
O
O
OB2O
O
P
O
O
O
OB1N3
OP
O
O
OB2O
O
P
O
O
O
ONH2 B1
S
1. base protection2. CBr4, PPh3, LiN3, DMF
3. o-ClPhOP(OCE)O2- Et3NH+,
Efimov coupling reagents4. NEt3
A', G'
C', T
1. Efimov:
2. TBAF3. NH3
P(Ph)3
NaIO4,
NaOH
+ P(Ph)3
1. base protection2. DMT-Cl3. o-ClPhOP(OPTE)O2
- Et3NH+,
Efimov coupling reagents4. H+

8
Ligation versus Cyclisation
OHP
O
O
OA/GO
O
P
O
O
O
OC/TNH
2
NC
N NH
+
NH
NH
+
NH
O
OHP
O
O
OA/GO
O
P
O
O
O
OC/TNH
2NH
NH
+
NH
O
OP
O
OA/GO
O
P
O
O
O
OC/TNH
2
XYn p
DimerisierungA:nXYp
- EDU
P
O
O
OA/GO
O
P
O
O
O
OC/TN
H
n
OHP
O
O
OA/GO
O
P
O
O
O
OC/TN
HP
O
O
OA/GO
O
P
O
O
O
OC/TNH
2
EDCnXYp
- EDU
CyclisierungB:- EDU
O
O
O
NH
P
PO
O
OO
OO
A/G C/T
+ H+
+ H2O
EDC
EDU
- H+
A
+
B
Oligomerisierung
12-Ring
CA-Cyclus
c( XY ) = 1-10 mM Cyclisierung
c( XY ) > 20 mM Oligomerisierung
n p
n p
B:
A:
Reaktionsbedingungen:
0.2 M EDC in 0.1 M HEPES-Puffer, 2° - 30°C

9
Oligomerisation of nCGp-dimersOligomerisation of nCGp-dimers
10 15 20
0
250
500
750
1000
1250
1500
2
4 6 8 10 1216
14 221820 24
26
nCGp
120 min
240 min
3 min
60 min
Abs
orpt
ion
[mV
]
time [min]
C G
C GC GC GC G
C GC GC G
C G C G
2-mer
4-mer
6-mer
8-mer

10
Reactivity of nYRp building blocks Reactivity of nYRp building blocks
Reaktivität: nCGp >> ( nTAp nTGp ) > nCAp
2mer, 30°C
0
10
20
30
40
50
0 100 200 300 400Zeit [min]
c[m
M]
nTGp,30°C
nTAp,30°C
nCGp,30°C
nCAp,30°C
4mer, 30°C
0
1
2
3
4
5
0 100 200 300 400Zeit [min]
c[m
M]
(nTGp)2,30°C
(nTAp)2,30°C
(nCGp)2,30°C
(nCAp)2,30°C
Dimer Tetramer
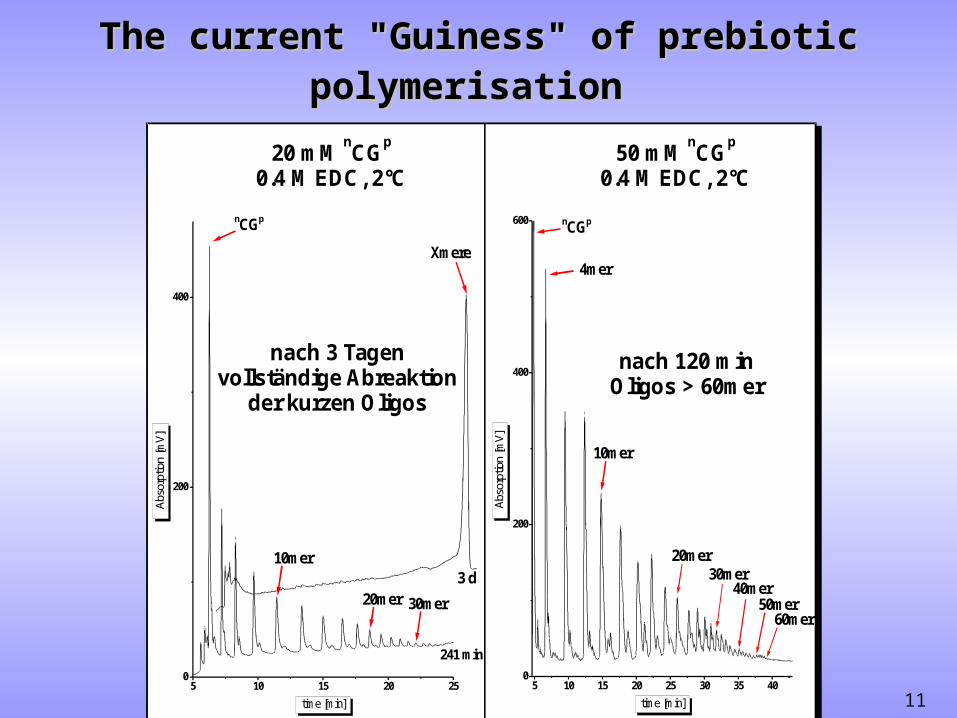
11
The current "Guiness" of prebiotic polymerisationThe current "Guiness" of prebiotic polymerisation
5 10 15 20 25 30 35 400
200
400
600
4mer
60mer
40mer50mer
20mer30mer
10mer
nCGp
Abs
orpt
ion
[mV
]
time [min]
50 mM CG0.4 M EDC, 2°C
n p
5 10 15 20 250
200
400
Xmere
3 d20mer 30mer
10mer
nCGp
241 min
Abs
orpt
ion
[mV
]
time [min]
20 mM CG0.4 M EDC, 2°C
n p
nach 3 Tagenvollständige Abreaktion
der kurzen Oligos
nach 120 minOligos > 60mer

12
No template effects in reactions using No template effects in reactions using single-sided building blocks single-sided building blocks
pteG C n pG C N 3
H OC G C G O H H OC G C G O H
G C G C O H
pteG C npG C G C G C O H
H OC G C G C G C G O HH OC G C G C G C G O H
ppteG C n G C G C G C O H
pteG C npG C G C O H
H OC G C G C G O H
pteG C npG C N 3
2 + 2an 4
2 + 4an 6
2 + 6an 8
H OC G C G C G O H
ppteG C n
HO(CG)3p + nCGpte je 20 m M + Te m plat HO(CG)4
o, 2° C,0.4 M EDC / 0.1 M HEPES, Produk t: HO(CG)3
pnCGpte
0
2
4
6
8
10
0 50 100 150 200 250Zeit [min]
c[m
M]
+0% Tem plat
+10% Tem plat
+20% T em plat
+40% T em plat

13
Earlier results from Zielinski & Orgel: Earlier results from Zielinski & Orgel:
Nature 1987: Experiments on a self-replicating tetraribonucleotide analogue confirmed our "square-root law". EDC as the source of energy, efficient replication in:
GCn + pGC --> GCnpGC
J. Mol. Evolution, a few years later: No self-replication at all in a slightly different system:
CGn + pCG --> CGnpCG
Speculations about the involvement of "slidomers".

14
Efficient oligomerisation via sliding, Efficient oligomerisation via sliding, concatenation, and concatomer ligation? concatenation, and concatomer ligation?
free oligomers
straight duplexes
slidomer duplexes
concatomerC GC GC G
G CG CG C
C G C G
G C G C
C GC GC GC G C G
G CG CG CG C G C
C G C G
G C G C
C GC GC GC G
G CG CG CG C
10-mer 4-mer 8-mer6-mer 4-mer
10-mer 4-mer 8-mer6-mer 4-mer
C GC GC G
G CG CG C
C GC GC G
G CG CG C
C G C G
G C G C
C GC GC GC G
G CG CG CG C
C G C G
G C G C
C GC GC GC G C G
G CG CG CG C G C
C G C G
G C G C
C GC GC GC G C G
G CG CG CG C G C
C GC GC G
G CG CG C
C GC GC GC G
G CG CG CG C
C G C G
G C G C
duplexation
sliding
aggregation

15
How a concatomer might lookHow a concatomer might look
C GC GC G
G CG CG C
C G C G
G C G C
C GC GC GC G C G
G CG CG CG C G C
C G C G
G C G C
C GC GC GC G
G CG CG CG C
10-mer 4-mer 8-mer6-mer 4-mer
10-mer 4-mer 8-mer6-mer 4-mer

16
Better base stacking via slidomer Better base stacking via slidomer concatenationconcatenation
CG dimer GC dimer CGCG slided duplex

17
Thermodynamic data support Thermodynamic data support slided concatomersslided concatomers
Stabilitätsverhältnis von Slidomer zu Duplex inAbhängigkeit von der Bausteinanzahl, B-Form
-120
-100
-80
-60
-40
-20
0
20
40
60
0 10 20 30Anzahl der Bausteine
G° (S
lid
om
er-
Du
ple
x)/
G° (D
up
lex
)in
%
dCGCG dCGCGCG dCGCGCGCG
pGCnpGCn
G0 = 0.22 kcal·mol-1 G0 = -4.19 kcal·mol-1
G = -14.24 kcal·mol-1
G0 = -8.60 kcal·mol-1
G37°C,Total = -26.81 kcal·mol-1
G = -7.12 kcal·mol-1 G = -7.12 kcal·mol-1
Energiegewinn durch Slidomerfortsetzung
pGCnpGCn
nCGpnCGp
nCGpnCGp nCGpnCGpnCGp
pGCnpGCnpGCn
nCGpnCGpnCGpnCGp
pGCnpGCnpGCnpGCn
nCGpnCGpnCGpnCGp
pGCnpGCnpGCnpGCn pGCnpGCnpGCn
nCGpnCGpnCGp
G37°C,Total = -12.57 kcal·mol-1

18
Two possible modes of ligationTwo possible modes of ligation
GC GC
CGCGCG CGCGCG
GCGCGCGC
CG CG
GC GC
CG CGCGCG
GCGCGC GC
CGCGCG
GCGCGC
CG CG
GC GC
GCGCGCGC GC
CGCGCGCG CG

19
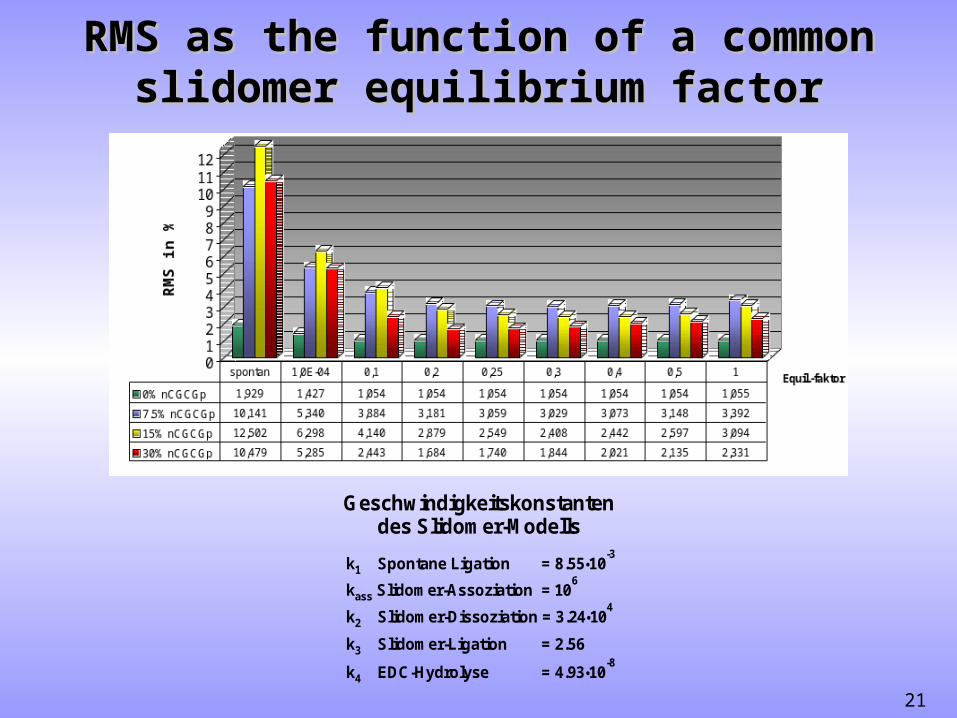
SimFitting supports slidomer modelSimFitting supports slidomer model
RMS = 12.5% RMS = 2.4%
n pCGCGn pC G + 7.5%, 15%, 30%
3rd order ligation slidomer model
rate parameter
k EDC hydrolysis = 4 4.93 10•-8
s -1
k 3rd order ligat. = M s1-2 -18.55 10•
-3
M s-1 -1k slidom er ass. = 10ass
6
k slidomer diss. = 2 3.24 10 s•4 -1
k slidom er ligat. = 3 2.56 M s-3 -1

20
Reaction modelReaction modelSpontanes Ligationsmodell
Nr. Reaktionsgleichung Simfit-Kurzform k
1 nCGp + nCGp + EDC → (nCGp)2 + EDU n2 + n2 + EDC → n4 + EDU k1
2 nCGp + (nCGp)2 + EDC → (nCGp)3 + EDU n2 + n4 + EDC → n6 + EDU k1
3 nCGp + (nCGp)3 + EDC → (nCGp)4 + EDU n2 + n6 + EDC → n8 + EDU k1
4 nCGp + (nCGp)4 + EDC → (nCGp)5 + EDU n2 + n8 + EDC → n10 + EDU k1
5 nCGp + (nCGp)5 + EDC → (nCGp)6 + EDU n2 + n10 + EDC → n12 + EDU k1
6 nCGp + (nCGp)6 + EDC → (nCGp)7 + EDU n2 + n12 + EDC → n14 + EDU k1
7 nCGp + (nCGp)7 + EDC → (nCGp)8 + EDU n2 + n14 + EDC → n16 + EDU k1
8 (nCGp)2 + (nCGp)3 + EDC → (nCGp)4 + EDU n4 + n4 + EDC → n8 + EDU k1
9 (nCGp)2 + (nCGp)4 + EDC → (nCGp)5 + EDU n4 + n6 + EDC → n10 + EDU k1
10 (nCGp)2 + (nCGp)5 + EDC → (nCGp)6 + EDU n4 + n8 + EDC → n12 + EDU k1
11 (nCGp)2 + (nCGp)6 + EDC → (nCGp)7 + EDU n4 + n10 + EDC → n14 + EDU k1
12 (nCGp)2 + (nCGp)7 + EDC → (nCGp)8 + EDU n4 + n12 + EDC → n16 + EDU k1
13 (nCGp)3 + (nCGp)3 + EDC → (nCGp)6 + EDU n6 + n6 + EDC → n12 + EDU k1
14 (nCGp)3 + (nCGp)4 + EDC → (nCGp)7 + EDU n6 + n8 + EDC → n14 + EDU k1
15 (nCGp)3 + (nCGp)5 + EDC → (nCGp)8 + EDU n6 + n10 + EDC → n16 + EDU k1
16 (nCGp)4 + (nCGp)4 + EDC → (nCGp)8 + EDU n8 + n8 + EDC → n16 + EDU k1
Erweiterung für Slidomermodell
Nr. Reaktionsgleichung Simfit-Kurzform k
(nCGp)2 + (nCGp)2 → (nCGp)2/(nCGp)2-Slidomerduplex
(nCGp)2 + n(CG)2p → (nCGp)2/
n(CG)2p- Slidomerduplex 17
n(CG)2p + n(CG)2
p → n(CG)2p/n(CG)2
p- Slidomerduplex
n4 + n4 → n4s 106
18 (nCGp)3 + (nCGp)3 → (nCGp)3/(
nCGp)3- Slidomerduplex n6 + n6 → n6s 106
19 (nCGp)4 + (nCGp)4 → (nCGp)4/(
nCGp)4- Slidomerduplex n8 + n8 → n8s 106
20 (nCGp)5 + (nCGp)5 → (nCGp)5/(
nCGp)5- Slidomerduplex n10 + n10 → n10s 106
21 (nCGp)6 + (nCGp)6 → (nCGp)6/(
nCGp)6- Slidomerduplex n12 + n12 → n12s 106
22 (nCGp)7 + (nCGp)7 → (nCGp)7/(
nCGp)7- Slidomerduplex n14 + n14 → n14s 106
23 (nCGp)8 + (nCGp)8 → (nCGp)8/(
nCGp)8- Slidomerduplex n16 + n16 → n16s 106
(nCGp)2/(nCGp)2-Slidomerduplex → (nCGp)2 + (nCGp)2
(nCGp)2/n(CG)2
p-Slidomerduplex → (nCGp)2 + n(CG)2p 24
n(CG)2p/n(CG)2
p-Slidomerduplex → n(CG)2p + n(CG)2
p
n4s → n4 + n4 k2
25 (nCGp)3/(
nCGp)3-Slidomerduplex → (nCGp)3 + (nCGp)3 n6s → n6 + n6 k2·10-4
26 (nCGp)4/(
nCGp)4-Slidomerduplex → (nCGp)4 + (nCGp)4 n8s → n8 + n8 k2·10-8
27 (nCGp)5/(
nCGp)5-Slidomerduplex → (nCGp)5 + (nCGp)5 n10s → n10 + n10 k2·10-12
28 (nCGp)6/(
nCGp)6-Slidomerduplex → (nCGp)6 + (nCGp)6 n12s → n12 + n12 k2·10-16
29 (nCGp)7/(nCGp)7-Slidomerduplex → (nCGp)7 + (nCGp)7 n14s → n14 + n14 k2·10-20
31 (nCGp)2/(nCGp)2-Slido + (nCGp)2/(nCGp)2-Slido + 2
EDC → (nCGp)4/(nCGp)4-Slido + 2 EDU
n4s + n4s + 2 EDC
→ n8s + 2 EDU
k3 (nCGp)2/n(CG)2p-Slido + (nCGp)2/(nCGp)2-Slido + 2
EDC → (nCGp)4/((nCGp)2(n(CG)2p))-Slido + 2 EDU
(nCGp)2/n(CG)2p-Slido + (nCGp)2/n(CG)2p-Slido + 2
EDC → (nCGp)4/(n(CG)2p)2-Slido + 2 EDU
(nCGp)2/n(CG)2p-Slido + n(CG)2p/n(CG)2p-Slido + 2
EDC → ((nCGp)2n(CG)2p)/(n(CG)2p)2-Slido + 2
EDU
n(CG)2p/n(CG)2p-Slido + n(CG)2p/n(CG)2p-Slido + 2
EDC → (n(CG)2p)2/(n(CG)2p)2-Slido + 2 EDU
32 (nCGp)2/(nCGp)2-Slido + (nCGp)3/(nCGp)3-Slido +
2 EDC → (nCGp)5/(nCGp)5-Slido + 2 EDU
n4s + n6s + 2 EDC
→ n10s + 2
EDU
k3
33 (nCGp)2/(nCGp)2-Slido + (nCGp)4/(nCGp)4-Slido +
2 EDC → (nCGp)6/(nCGp)6-Slido + 2 EDU
n4s + n8s + 2 EDC
→ n12s + 2
EDU
k3
34 (nCGp)2/(nCGp)2-Slido + (nCGp)5/(nCGp)5-Slido +
2 EDC → (nCGp)7/(nCGp)7-Slido + 2 EDU
n4s + n10s + 2
EDC → n14s +
2 EDU
k3
35 (nCGp)2/(nCGp)2-Slido + (nCGp)6/(nCGp)6-Slido +
2 EDC → (nCGp)8/(nCGp)8-Slido + 2 EDU
n4s + n12s + 2
EDC → n16s +
2 EDU
k3
36 (nCGp)3/(nCGp)3-Slido + (nCGp)3/(nCGp)3-Slido +
2 EDC → (nCGp)6/(nCGp)6-Slido + 2 EDU
n6s + n6s + 2 EDC
→ n12s + 2
EDU
k3
37 (nCGp)3/(nCGp)3-Slido + (nCGp)4/(nCGp)4-Slido +
2 EDC → (nCGp)7/(nCGp)7-Slido + 2 EDU
n6s + n8s + 2 EDC
→ n14s + 2
EDU
k3
38 (nCGp)3/(nCGp)3-Slido + (nCGp)5/(nCGp)5-Slido +
2 EDC → (nCGp)8/(nCGp)8-Slido + 2 EDU
n6s + n10s + 2
EDC → n16s +
2 EDU
k3
39 (nCGp)4/(nCGp)4-Slido + (nCGp)4/(nCGp)4-Slido +
2 EDC → (nCGp)8/(nCGp)8-Slido + 2 EDU
n8s + n8s + 2 EDC
→ n16s + 2
EDU
k3
40 2 (nCGp)2 + (nCGp)2/(nCGp)2-Slido + 2 EDC
→ (nCGp)3/(nCGp)3-Slido + 2 EDU
2 n2 + n4s + 2
EDC → n6s + 2
EDU
k3
41 2 (nCGp)2 + (nCGp)3/(nCGp)3-Slido + 2 EDC
→ (nCGp)4/(nCGp)4-Slido + 2 EDU
2 n2 + n6s + 2
EDC → n8s + 2
EDU
k3
42 2 (nCGp)2 + (nCGp)4/(nCGp)4-Slido + 2 EDC
→ (nCGp)5/(nCGp)5-Slido + 2 EDU
2 n2 + n8s + 2
EDC → n10s + 2
EDU
k3
43 2 (nCGp)2 + (nCGp)5/(nCGp)5-Slido + 2 EDC
→ (nCGp)6/(nCGp)6-Slido + 2 EDU
2 n2 + n10s + 2
EDC → n12s + 2
EDU
k3
44 2 (nCGp)2 + (nCGp)6/(nCGp)6-Slido + 2 EDC
→ (nCGp)7/(nCGp)7-Slido + 2 EDU
2 n2 + n12s + 2
EDC → n14s + 2
EDU
k3
45 2 (nCGp)2 + (nCGp)7/(nCGp)7-Slido + 2 EDC
→ (nCGp)8/(nCGp)8-Slido + 2 EDU
2 n2 + n14s + 2
EDC → n16s + 2
EDU
k3
46 EDC → EDU EDC → EDU k4
21
RMS as the function of a common RMS as the function of a common slidomer equilibrium factorslidomer equilibrium factor
Geschwindigkeitskonstantendes Slidomer-Modells
k EDC-Hydrolyse = 4 4.93 10•-8
k Spontane Ligation = 1 8.55 10•-3
k Slidomer-Assoziation = 10ass6
k Slidomer-Dissoziation = 2 3.24 10•4
k Slidomer-Ligation = 3 2.56

22
"Template" addition even inhibits "Template" addition even inhibits polymerisationpolymerisation
nCGp 20 m M + Tem plat H OCGCGOH , 2°C,0.4 M EDC/0.1 M HEPES, Produkt: 8m er
0 ,0 0
0 ,0 2
0 ,0 4
0 ,0 6
0 ,0 8
0 2 0 4 0 6 0Zeit [m in]
c[m
M]
8m er, 0%T
8m er, 10%T
8m er, 20%T
8m er, 40%THOGCGCOHHOGCGCOH
pnCG HOCGCG OH HOCGCG OH
HOGCGCOHHOGCGCOH
pHOCGCG OH nCGpnCG HOCGCG OH
HOGCGCOH
Modes of inhibitionby "non-reactive" template
4mer1.
6mer2.
8mer3.
nCGp
HOCGCG OH
HOCGCGCG OH
HOCGCGCGCGOH
20m M10%, 20%, 40%
Templat+

23
Summary and possible significanceSummary and possible significance The current picture to make long prebiotic oligomers is by primer-extension on a solid support (clay) via feeding with nucleotide-phosphorimidazolides (Ferris & Orgel, "crepes scanario"). Traces of 50-mers can be detected after several weeks and daily replenishment of the imidazolides.
"Self-elongation" as an alternative picture: In the presence of the dehydration reagent EDC, the dimer nCGp yields high molecular weight oligomers (quantitatively for n >> 40) after 3 days.
"Self-elongation" and "self-replication" may be different sides of the same coin. Exactly the same reason that caused poor self-replication in a comparable system causes efficient polymerisation in our system.
Eigen, Hartman, and others have speculated that the earliest "genes" were rich in C and G, or even CG-repeats. Our experiments indicate that one may neither need templates nor surfaces to arrive at such structures.
Outlook: Co-oligomerization experiments with nYRp are expected to result in materials still rich in CG but "being doped" with other bases. Such materials may have the capacity to fold into discrete secondary structures.


















