
1 I ! ;J,~ Estuari ne Research - University of Southern...
20
GG ~I C' -, '~I""" j5't ; C1:J'-;' , 1 1 I _! ;J,~ Estuari ne Research VOLUME II Geology and Engineering Edited by L, Eugene Cronin Estuarine Research Federation @ Academic Press, Inc. NEWYORK SANFRANCISCO LONDON 1975 A Subsidiary of Harcourt Brace Jovanovich, Publishers
Transcript of 1 I ! ;J,~ Estuari ne Research - University of Southern...

GG~I C'-, '~I"""
j5't; C1:J'-;' ,1 1 I _!
;J,~
EstuarineResearch
VOLUME II
Geology and Engineering
Edited by
L, Eugene CroninEstuarine Research Federation
@Academic Press, Inc. NEWYORK SANFRANCISCO LONDON 1975
A Subsidiary of Harcourt Brace Jovanovich, Publishers

CoPYRIGHT@ 1975, BY ACADEMIC PRESS, INC.ALL RIGHTS RESERVED.
NO PART OF THIS PUBLICATION MAY BE REPRODUCED OR
TRANSMITTED IN ANY FORM OR BY ANY MEANS, ELECTRONICOR MECHANICAL, INCLUDING PHOTOCOPY, RECORDING, OR ANYINFORMATION STORAGE AND RETRIEVAL SYSTEM, WITHOUTPERMISSION IN WRITING FROM THE PUBLISHER.
ACADEMIC PRESS, INC.111 Fifth Avenue, New York, New York 10003
United Kingdom Edition published byACADEMIC PRESS, INC. (LONDON) LTD.24/28 Oval Road. London NWI
Library of Congress Cataloging in Publication Data
International Estuarine Research Conference, 2d,
Myrtle Beach, SoCo, 1973.Estuarine research.
Papers presented at a conference held by theEstuarine Research Federation and cosponsored by theAmerican Society of Limnology and Oceanography and theEstuarine and Brackish Water Sciences Association.
Bibliography: poIncludes index.
CONTENTS: v.1. Chemistry and biology.-v. 20 Geology and engineering.
1. Estuaries---Congresseso 20 Estuarine ocean-ography---Congresses. 3. Estuarine biology---Congresses.I. Cronin, Lewis Eugene, (date) no EstuarineResearch Federation. III. American Society ofLimnology and Oceanography. IV. Estuarine andBrackish-water Sciences Association. V. Title.GC96.I57 1973 551.4'609 75-29370ISBN 0-12-197502-9 (vo2)
PRINTED IN THE UNITED STATES OF AMERICA

SUBMERGENTVEGETATIONFOR BOTTOMSTABILIZATION
by
Lionel N. Eleuterius I
ABSTRACT
Three species of sea grass, Thalassiatestudinum, Cymodocea manatorum, andDiplanthera wrightii were transplanted from natural habitats to barrensubmergent spoil areas and to control areas located adjacent to undisturbedsea-grassbeds. Anchoring devices were developedto hold the transplants in placeon the bottom regardless of water depth. Diplanthera had the greatestpercentage of survival, its growth rate for exceeding that of Thalassia.Cymodocea did not succeed at all. Some characteristics of vegetativemorphology and rate of growth were compared under varying conditions ofsediment deposition and erosion. In viewof its distribution, growth, tolerances,and low rates of deposition, Diplanthera was judged the best candidate forfurther transplant studies. Successful transplants formed established beds incontrol areas. Erosion and sedimentation rates exceeded the growth capacity ofthe plants although no successful transplants were obtained on spoil. Lowtemperatures and prolonged exposure to low-salinity waters apparently affectsea-grass beds and transplants adversely. Available plant nutrient levels ofbottom (substrate) samples were not found to vary appreciably betweenvegetated and barren areas.
INTRODUCTION
Sea-grass beds in the Gulf of Mexico are composed predominantly of the
1. Gulf Coast Research Laboratory, Ocean Springs, Mississippi 39564.
439

LIONEL N. ELEUTERIUS
marine angiosperms Diplanthera wrightii Ascherson (shoal grass), Thalassiatestudinum Konig (turtle grass), and Cymodocea manatorum Ascherson(manatee grass). Their distribution and ecology in various regions of thenorthern Gulf of Mexicohave been studied by Humm (8), Phillips (14, 15, 17),Strawn (24), Moore (12) and Eleuterius (4). Animal communities associatedwith sea-grassbeds in the waters of Florida have been reported upon by Voss andVoss (28), Tabb and Manning (25), Stephens (22), and O'Gower and Wacasey(13). Gunter (6) reported that most of the pink shrimp production in Texascomes from the Gulf opposite Redfish Bay, which has one of the largestJ'halassia communities on the Texas coast. Hoese and Jones (7) studiedvegetated sea bottoms in Texas and the associated larger animals. Christmas et al.(3) made special reference to the abundance of postlarval penaeid shrimp in thegrass beds near Horn Island, Mississippi.They pointed out that more shrimpwere taken at sampling stations that included grass beds than at those withoutthem. These studieshave shown that sea-grasshabitats support a large and variedcommunity of marine animals in the Gulf of Mexico. Similar reports by Wood(27) and Fuse et al. (5) from other parts of the world indicate that vegetated seabottom provides one of the richest and most productive of marine communities.
The ability of vegetation to prevent erosion of the soils of the earth has beenrealized for a long time, and its use in soil-conservation practices related toagriculture, forestry, and highway construction is well documented. For anumber of years Europeans have used emergent vegetation along stream andriver banks to stabilize them against erosive currents (21). More recentlysubmerged vegetation has been used for bottom stabilization of Americanwaterways (19, 20). These reports, of course, relate to fresh-water systems. The\!lseof submergentmarine flowering plants for bottom stabilization has had littlestudy. Kelly et al. (9) transplanted Thalassia in Boca Ciega Bay, Florida, andPhillips (17) transplanted Zostera marina in Puget Sound, Washington.However,natural establishment of Diplanthera on submerged spoil areas from Port Isavelto Port Mansfield, Texas, has been reported by Breuer (1, 2). Stevens (23)reported sea grassesgrowing on spoil in Corpus Christi Bay, Texas. Diplantherahas also been reported to occur on spoil areas in Sarasota Bay, Florida (GordonGunter, personal communication). Strawn (24) reported that at Morehead City,North Carolina, the bay scallop fishery was partially restored by the increase ofDiplanthera. Thus, successful establishment of sea grasseson barren spoil wouldhave two beneficial effects: the stabilization of spoil and the provision ofadditional productive habitat.
There were three primary objectives in the present study: a. development ofeffective methods for transplanting sea grasses; b. determination of the bestspecies for transplanting; and c. identification of some of the limiting ecologicalfactors by studying existing grass beds and experimental areas. This projectbegan in July 1971 and the first transplants were made in the fall of the same
440

ESTUARINE RESEARCH
year.Mississippi Sound is bordered on the south by four barrier islands, and four
river systems contribute directly to the fluctuating salinity gradient across it.Most of the sea-grasscommunities are located north of the barrier islands and inthe island passes. The.grass beds in the shallow waters along the islandsoccupyareas of various size, thus their distribution here is discontinuous. In the deeperwaters grass beds are more extensive in area. The spoil islands used asexperimental areas in this study were created primarily as the result ofmaintenance dredging of the Pascagoula ship channel. General location ofsea-grassbeds and transplanting sites are shown in Figure 1.
MISS'SS,I'I',
roo,.
_.~\)!/'SOU1'l°
..-~c~ P\'~!f>~\,\.
~ .-x= "..""""'.f> "'"
FigUre1. Transplanting sites (x) of sea grasses. The stipled area indicates general locationof naturally established grass beds.
METHODSAND MATERIALS
The three common species of sea grass shown in Figure 2 were transplantedfrom natural sea-bed habitats near Horn Island, Mississippi, to spoil islandsadjacent to the Pascagoula ship channel and to barren areas which served ascontrols adjacent to areas vegetated by seagrasses.
Two types of anchoring devicesdevelopedand used in this study are shown inFigures 2-D, 3-A, and 6-C. The specific details of development of these deviceswill be reported elsewhere (Eleuterius, in preparation). In order to determine thebest time of year to conduct transplanting. operations, transplants were madeseasonally. Vegetative morphology and rate of growth were noted and comparedunder varying conditions of sedimentation and erosion rates. General
441

LIONEL N. ELEUTERIUS
A ~'"
~
Figure2. Three marine angiosperms found in Mississippi Sound used in transplant study.(A) Diplanthera wrightii (shoal grass), (B) Cymodocea manatorum (manateegrass), (C) Thalassia testudinum (turtle grass), (D) T. testudinum attached towire-mesh anchor.
characteristics such as nutrient levels and developmental patterns of sea.grassbeds establishednaturally and by transplantation were compared to barren areas.
Transplant material was dug from natural habitats on low tides and most of itwas transported by boat to the laboratory some twenty.five miles away. During
442

ESTUARINE RESEARCH
Figure3. Transplants, anchors, planting techniques, and study areas along Horn Island.(A) Diplanthera wrightii tied to coated iron rod. Plastio-coated wire was used inattaching transplants to anchor. (B) Tying anchors together in trotline fashion.(C) Wile-mesh anchors arranged along shore to demonstrate transplantingtechnique. (D) Wire mesh with Diplanthera wrightii attached. (E) Study areaalong Horn Island. Pipes were used as permanent markers. (F) Hand emplacedtransplants (attached to construction rods) on barren areas adjacent to grassbeds at Horn Island.
443

1
LIONEL N. ELEUTERIUS
the first year, transplants were moved to adjacent bar'ren areas and replanted thesame day they were dug from the habitats (Fig. 3-F). The sea grassesthat werereturned to the laboratory were stored at room temperature in aerated sea water.Transplants were then selected and attached to anchoring devices cut fromconsturction rod and heavy-gaugewire mesh.
Each wire-meshanchor, approximately 45 cm x 45 cm (18" x 18") of 15 cm(6") mesh, weighed approximately 190 grams, and each iron rod,approximately 20 cm (8") in length, weighed 110 grams. The anchors werecoated with white vinyl paint and transplants were attached to them withplastic-coated copper wire. The rust year all transplants were emplaced by handor broadcast over the study areas. During the second year transplant units weretied together in trotline fashion by nylon cord in order to relocate the anchorsand record the condition of the transplants attached to each (Fig. 3-B, C, D,6-D).
The transplants were laid out in rectangularbelts along the experimental spoilislands and on barren areas adjacent to or within the sea-grasshabitats (Fig. 4-A).
RESULTS
Transplants were made simply by sprigginginitially, but all were pulled fromthe substratum by wave action and currents in control areas and spoil areas. Avariety of anchoring devices, whose emplacement required a person to be in thewater, were used during this project. The ones described herein were developedfor employment in either shallow or deep waters and proved to be especiallysuitable for depths too great to hand plant by wading.
The inshore submerged spoil area composed primarily of mud wascontinously covered by highly turbid waters ranging in salinity from0%0-15%0. The offshore spoil area consisted primarily of sand and wascovered by waters ranging in salinity from 0%0-35%0. Both areas had clamshell intermixed, the proportion, however, was relatively low. No submergedvegetation existed on either area.
No appreciable difference in available nutrients was noted between barrenspoil, sea-grass beds, and adjacent barren areas. Slightly higher averages inphosphorus and potassium were observed in the older portion of sea-grassbeds,probably due to the accumulation of organic matter. Samples from theperiphery of colonieshad lower levels and approximated adjacent barren areas.Phosphorus ranged from 23 kg (50 Ibs) to 80 kg (175 Ibs) per acre. Potassiumranged from 46 kg (100 Ibs) to 136 kg (300 Ibs) per acre. Nitrogen was less than4.5 kg (10 Ibs) per acre for all areas sampled. Organic matter was recorded atapproximately 2% in Cymodocea and Thalassiabeds and slightly over 1% inDiplanthera beds. Adjacent barren areas varied from slightly less than 1% toslightly less than 2%. Hydrogen ion concentration (PH) ranged from 6.2 to 8.2
444
...
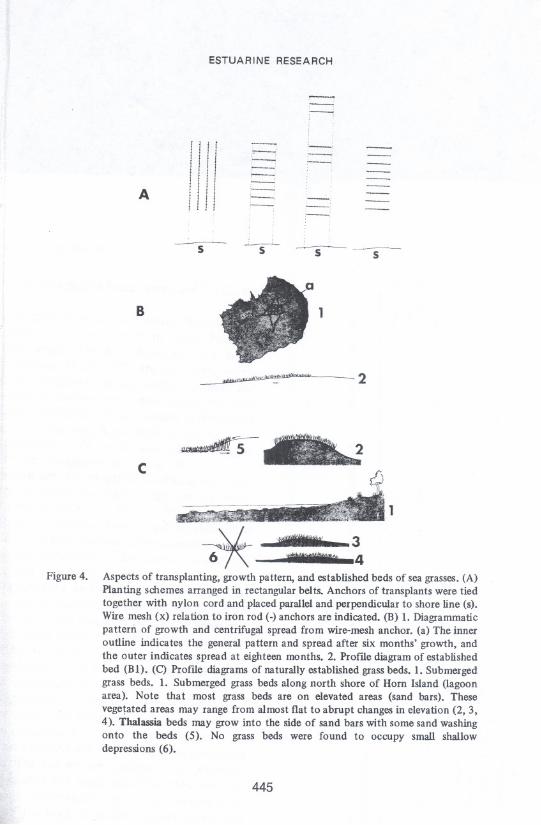
Figure 4.
ESTUARINE RESEARCH
AI
'-
--s-5 s-- ~
B 1
~2
~
1
c
*
445

LIONEL N. ELEUTERIUS
with averagesof 7.2 for grassbeds; 7.6 for adjacent barren areas, and 7.3 for thebarren spoil area at Horn Island.
None of the transplants made on spoil areas were successful. Mortality wasdue to the extremely unstable bottom. The spoil at Horn Island Pass wasreequently exposed to heavy seas which reworked drastically the sandysubstratum. From November 1971 through March 1972, approximately 60 em(24") of sand were deposited over the transplants on the south, southeast, andeastern section of the spoil. No velocity measurements were made, but tidalcurrents through the passesare alwaysconsiderable. These currents at times wereso great that a person could not remain standing in the shallow waters of thespoil areas at Horn Island Pass. Transplants made on the west side of the spoilwere quickly eroded away and no further transplants were made there.
Mortality of transplants on the spoil near the mouth of the Pascagoula Rivermay have resulted from a number of causes; inadequate light due to highlyturbid water, exposure to the great quantity of low-salinity fresh-waterdischarged from the Pascagoula River, or siltation. Survival and establishmentwere attained in transplants of Diplantheraand 'l11alassiain the control area nearHorn Island. The number of successful transplants of Diplanthera was almosttwice that for 'l11alassia.No successwas achieved in transplants of Cymodocea.Survival data showing dates of transplanting and subsequent observation areshown in Table 1. Since most anchors of transplants made by the broadcastmethod could not be found, evaluation of each transplant was not possible. (Atthe time of writing this paper a U. S. Army surplus metal mine detector (modelDT-44D{PRS-3)has been successfully employed to locate anchoring devicesplaced by broadcasting. This sensing device will be used to ascertain rapidly andmore accurately those grass beds established from transplants attached toanchors. This location process is on-goingand the results were not available forincorporation herein.) An effective method of tying the anchors together bynylon cord was developed to aid in relocating the units. Most transplants werekilled by sediment deposition. The predominant sediment along Horn Island andspoil at Horn Island Pass was sand. The inshore area sediment was primarilymud. Some transplants were lost to erosion and many may have been killed byman-made distrubances. Approximately 100 transplants were found entangled inthe nylon cord and anchors, obviously the work of vandals. Others were foundstrewn on high dunes near the study areas. Whether or not the transplants werekilled before or at the time of this disturbance is not known.
Elevational changes due to sedimentation are illustrated in Figure 5.Deposition rates greater than 2.5 cm (1") per month appeared to exceed thegrowth rate of 'l11alassiatestudinum and Cymodocea manatorum and depositiongreater than 5 cm (2") per month apparently exceeded the growth rate ofDiplanthera wrightii. The plants were obviously not able to adjust to rapidincreases in substrate elevation caused by sand deposition and death resulted.
446

TABLE 1.
Surviving, successfully established and mortality of transplants of Diplanthera wrigbtii, Thalassia testudinum and Cymodocea manatorum, forthe period April, 1972 through September, 1973.
.j::.
.j::."
fs
d:h
* Two groups placed in separate area.(BC)=Broadcast method(T) =Anchors tied together with nylon cord.
Location of Number of Number of Number of Number of Number c
Transplant Transplant Anchoring Observation Transplants Transplants units not Observation TrnasplanDate Area Species Units used Date alive Dead located Date establishe
Rod Mesh Rod Mesh Rod Mesh Rod Mesh Rod Me
3 Apr 72 Spoil-Horn Diplanthera (BC) 60* 30* 6 Jut 72 60* 30*Island Pass MaIassia (BC) 60* 30* 60* 30*
Cymodocea (BC) 60* 30* 60* 30*
1 May 72 Spoil-Horn Diplanthera (BC) 30 15 10 Jut 72 30 15Island Pass ThaIassia (BC) 30 15 30 15
Cymodocea (BC) 30 15 30 15
1 May 72 Spoil-Mouth Diplanthera (BC) 30 15 10 Jut 72 30 15Pas. River ThaIassia (BC) 30 15 30 15
Cymodocea (BC) 30 15 30 15
25 Jut 72 Spoil-Horn Diplanthera (BC) 60 30 20 Aug 72 60 30
Island Pass ThaIassia (BC) 60 30 60 30
Cymodocea (BC) 60 30 60 30
3 Apr 72 Horn Island Diplanthera (BC) 30 15 6 Jut 72 20 10 10 20 6 Aug 75 14 8
(lagoon area) ThaIassia (BC) 30 15 15Aug 72 10 6 20 9 7 4
Cymodocea (BC) 30 15 0 0 30 15
25 Jul 72 Horn Island Diplanthera (BC) 60* 15* 15 Aug 72(Lagoon area) Th?\assia (BC) 70* 15*
27 Jul 72 Horn Island Diplanthera (BC) 40* 15 Aug 72ThaIassia (BC) 38*
27 Jul 72 Horn Island Diplanthera (n 25 15 Aug 72 0 25
Thalassia (n 25 0 25
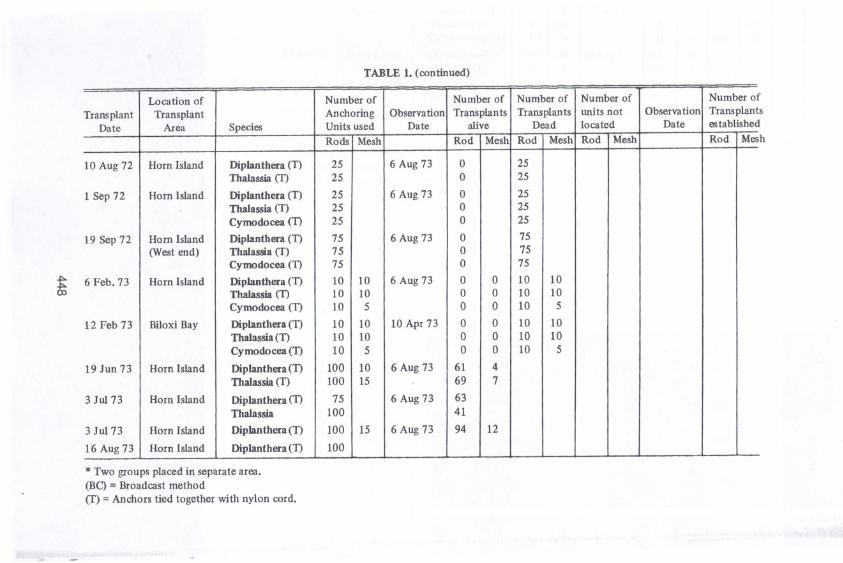
TABLE 1. (continued)
~~00
h
* Two groups placed in separate area.(BC) =Broadcast method<T)=Anchors tied together with nylon cord.
. .Location of Number of Number of Number of Number of Number of
Transplant Transplant Anchoring Observation Transplants Transplants units not Observation TransplantDate Area Species Units used Date alive Dead located Date established
Rods Mesh Rod Mesh Rod Mesh Rod Mesh Rod Mes
10 Aug 72 Horn Island Diplanthera (T) 25 6 Aug 73 0 25Thalassia (T) 25 0 25
1 Sep 72 Horn Island Diplanthera (T) 25 6 Aug 73 0 25Thalassia (T) 25 0 25
Cyrnodocea(TI 25 0 25
19 Sep 72 Horn Island DipJanthera(TI 75 6 Aug73 0 75(Westend) Thalassia <T) 75 0 75
Cyrnodocea(TI 75 0 75
6 Feb. 73 Horn Island Diplanthera (TI 10 10 6 Aug 73 0 0 10 10Thalassia(T) 10 10 0 0 10 10
Cyrnodocea(TI 10 5 0 0 10 5
12 Feb 73 BiloxiBay DipJanthera(TI 10 10 10 Apr 73 0 0 10 10Thalassia(TI 10 10 0 0 10 10
Cymodocea(TI 10 5 0 0 10 5
19 Jun 73 Horn Island Diplanthera(TI 100 10 6 Aug 73 61 4Thalassia(T) 100 15 69 7
3 Ju173 Horn Island Diplanthera(TI 75 6 Aug 73 63Thalassia 100 41
3 Ju173 Horn Island Diplanthera(TI 100 15 6 Aug 73 94 12
16 Aug 73 Horn Island Diplanthera(TI 100

ESTUARINE RESEARCH
Figure5. Sequential stages in the establishment of a bed of Diplanthen wrightii from ananchored transplant. (A) Transplant attached to iron-rod anchor lies on surfaceof substratum initially. (B) Anchor becomes covered by sand soon aftertransplanting. (C) Anchor covered with 5 ern (2 inches) of sediment deposition.(D) Established bed of Diplanthera, approximately six months from time oftransplanting. (E) Well-established bed of Diplanthera; note increase in elevationof bed due to sediment deposition in this example. Successful transplants andestablished beds, however, were achieved with relatively less sediment thanillustrated here.
449

..
~I
LIONEL N. ELEUTERIUS
Few transplants were killed by erosion in the control areas along Horn Island.However, detrimental effects related to transplanting during unfavorable seasonswere obviously responsible for transplant mortality. Some trnasplants died fromunknown causes. Since sediment deposition and erosion were not excessive itappears that detrimental physiological effects are caused by digging, preparing,attaching, or holding the transplants. However, during the first year of thisstudy, from four to six transplants from grass beds were inunediately placed inadjacent barren areas. All of them died. Transplants have been held affIxed toanchoring devices in the laboratory for up to six weeks and growth wasapparent, especiallyon Diplanthera.
During the winter of 1971-72 the seagrassesin MississippiSound did not losetheir leaves and were therefore easily located. However, during the winter of1972-73 all sea grasses observed lost their leaves and the beds were not easilydistinguished from barren areas. The loss of leaves was apparently a result of acombination of low temperatures, and unusual fresh-water discharge from localrivers, and the opening of the Bonnet Carrl'fSpillway from mid-February tomid-June 1973. Only short stubble remained on the surface of the beds. Notransplants were made during the springof 1973 although considerable time wasspent in search of suitable transplant material. Most beds observed appeared tobe dead or dying. However, complete death and erosion of these beds did notoccur and new leaves began to appear in late June 1973. Obviously theenvironmental conditions mentioned above as affecting sea-grassbeds were also amajor factor affecting transplants made from September 1972 to June 1973.Similar leaf loss from sea grasses occurred furing the winter of 1969 and wasattributed at that time to low temperatures since rainfall was not excessive.
During periods when the open (covering) waters were "fresh"(00/00-100/00) the soil waters had consistently higher salinities. Perhaps highersoil-water salinities protected the roots and rhizomes from rapid changes insalinity. Exposure of roots and rhizomes of transplants to this low-salinity watermay have killed them.
The most remarkable and apparent difference between the two successfulspecies, (Diplantheraand Thalassia,)was the prolific growth of the former. Thebest example of growth was achieved by a transplant made in April 1972.Rhizomes had spread centrifugally approximately 124 cm (48") by July 1972and approximately 92 cm (60") by late August 1972. This entailed an averagegrowth rate of 27 cm (10.5") per month. Thus one rhizome grew approximately14 cm (5") per month. Kellyet aI. (9) reported that Thalassiagrew at an annualrate of 20 cm (8") or less in Boca CiegaBay, Florida. By the end of November1972 the bed was estimated to be 2.6 meters (8.5') in diameter. Observationsmade on this same establishedbed on 6 August 1973 showed that its centrifugalspread had increased to 3.7 meters (12') across the greatest width and 2.4meters (8') across the narrowest. This is illustrated diagranunatically in Figure
450

ESTUARINE RESEARCH
4-B. The anchoring device was located approximately 28 cm (11") beneath thesurface of the substratum.
Naturally established grass beds of Thalassiaand Cymodocea are generallyfound in deeper and far less turbulent waters than those of Diplanthera.In deep,calm water continuous stands are formed, but the species are practically neverintermixed. In shallow water, especially along sandy beaches and bars (shoals),Diplanthera generally occupies the shallowestdepths and alwaysextends nearestthe shoreline, being therefore exposed almost continuously to turbulentconditions. Thalassia and Cymodocea beds were found only where sedimentdeposition rates were low and generally in areas where erosion was apparent.Shoal grass was found in areas of heavy sand deposition and also in areas ofheavy erosion. Practically all vegetated areas are elevated as shown in Figure 4-C,especially near the lagoon area of Horn Island. However, extensive beds at themiddle ground and in deeper waters along Horn Island are also found on longgentle slopes.
The ability of Diplanthera to occupy bottom exposed to relatively highenergy is due in part to the extensive rhizome and root mass produced. Theproliferation of these fine rhizomes and roots binds the substratum andcontibutes directly to stabilization. Water movement across stabilized bedscarries away little sediment and in near-shoreareas over which wavesbreak thereis generally a deposition of sediments when leavesare present. Shoals (vegetatedand non-vegetated) are built up to a certain elevation, to which they arerestricted. Thus, they do not get above a certain elevation in relation to meanlow water (MLW). These shoals are generally exposed on tides lower than MLWand of course especially at extremely low tides. Some of the transplantsextended over and between sand bars, and those on the highest elevations didnot become covered with much sediment (Fig. 6-E, 6-F). In many such instances5 cm (2") of sediment was deposited within two months after transplanting andno further deposition appears to have occurred. On barren areas between or onthe side of sand bars various amounts of deposition were recorded and werealways greater than that found on top of the bars. Some bars have less elevationthan others, and appear to be building. Establishment of vegetation on theselesser bars increased the building rate.
The peculiar morphology of Diplanthera also favors the growth of the plantin these high-energy areas. Waves and currents apparently exert less frictionalforce on its narrow leaf blades than they do on the wide blades of Thalassiaandthe relatively large terete blades of Cymodocea. These larger-leavedspecieswereobserved to be quickly pulled from the substratum when transplanted to areasconstantly or periodically exposed to high energy. Another characteristiccontributing to the successof Diplantherawas the flexibility of the blades. Theybend easily, whereas those of Thalassiaand Cymodocea are less flexible andmore easily broken.
451

D
Figure6. Spoil islands, naturally established grass beds, transplanting techniques andestablished beds of DipIanthera wrightii from transplants with wire meshanchors. (A) Barren spoil islands at Horn Island Pass. The submerged spoilshown around the island in the lower portion of the photograph was used astransplanting site. (B) Naturally established sea grass beds north of Horn Island.The beds appear as dark patches on the bottom. (C) Tha1assia testudinum tiedto coated iron rod anchor. Plastic coated copper wire was used to tie transplantsto anchors. (D) Transplants and anchors tied together in trotline fashion to aidin rapid relocation of units for evaluation. (E) F O. DipIanthera wrightii bedestablished from wire meshes. The beds are shown approximately six monthsafter transplanting. These beds have subsequently tripled in area. (Note outlineof wire mesh anchors.)
The rhizomes and roots of Thalassiaand Cymodocea are also larger thanthose of Diplanthera, but their proliferation is not as great and their network ofinterweavingdoes not bind the substratum as well.
452
LIONEL N. ELEUTERIUS
A
B.JL
c

ESTUARINE RESEARCH
DISCUSSION
The environmental conditions that existed during the winter of 1971-72werequite different from those found during the winter of 1972-73. The leafless,non-flowering condition found in the spring of 1973 was in sharp contrast withthe luxurious leaf growth and flower production in the spring of 1972. The greatoutflow of fresh water and resulting low salinitiesprobably affected natural grassbeds and transplants more than any other factor through July of 1973 andcontributed to the inability to duplicate in 1973 the successfulresults obtainedin the spring and summer of 1972.
The low survival rate of transplants shows that the successful transplantingand establishment of sea-grassbeds for bottom stabilization or as a habitat is nota simple matter. However, its survival and rapid growth qualify Diplantheraas abetter candidate for more detailed study than Thalassia.
Diplanthera is found along the Atlantic coast from North Carolina to Floridaand from Florida to Texas, but Thalassiaand Cymodocea do not occur on theAtlantic coast, except for a short extension northward around the tip of extremesouth Florida. Diplanthera apparently has wide environmental tolerances.McMillan and Moseley (11) showed in a comparative study in Texas thatDiplanthera had a greater tolerance to salt than any other speciestested. In aprevious study (4) Diplanthera was found to occur over a larger areaa of bottomthan Thalassiaor Cymodocea in MississippiSound. Of the four marine floweringplants studied only Diplanthera extended into low-salinity waters along themainland shore. It also occurred in areas exposed to highest salinities, forexample, in the shallow waters near the barrier islands and in the open passes.Diplanthera alone is found in the island passes exposed to powerful waves andstrong tidal cunents. These characteristic tolerances allow wide distribution andindicate greater application of research results throughout the south Atlantic andnorthern Gulf of Mexico than would be possible with Thalassiaor Cymodocea.
The particular set of environmental conditions that must exist during the.time of natural establishment may be present only briefly each year or not at allfor several years. Exactly what those conditions are is not known, but it hasherein been ascertained that they are of paramount importance if grassbeds onnatural or man-made sea bottoms are to be successfullyestablished.
There is no doubt that once established, vegetation suppresseserosion of seabottom within limits, but heavy deposition of sand and other sediments erodedfrom adjacent areas may suppress plant growth or kill the beds quickly. Slightaccumulation of sediments in Thalassia and Cymodocea beds appears to becharacteristic, but heavier rates were found in Diplanthera beds. The rapidgrowth of Diplanthera apparently enables it to grow under greater rates ofsedimentation. The spoil upon which transplants were made was not a suitable
453
~

LIONEL N. ELEUTERIUS
habitat for sea grasses, even though a large Diplanthera bed is found nearby.Perhaps more time is needed for the particular spoil to stabilize to physicalforces before conditions within the growth capacity of Diplanthera are reached.Sea bottom areas with slight disturbance, if only seasonal, appear to be morefavorableto the establishmentof vegetation. .
ACKNOWLEDGMENTS
This project was funded by the Corps of Engineers, U. S. Army, undercontract no. DACWOl-71-C.olO5.I thank Dr. Gordon Gunter and Mr. CharlesEleuterius for their reviewand criticism of the manuscript.
REFERENCES
1. Breuer, J. P.1960 Life history studies of the important sports and commercial fish
of the lower Laguna Madre- Texas Game and Fish CommissionMarine Laboratory. Coastal Fisheries Project Reports. 5 p.
2. Breuer, J. P.1961 Life History studies of the marine flora of the Lower Laguna
Madre area. Texas Game and Fish Commission MarineLaboratory. Coastal Fisheries Project Reports. 2 p.
Christmas, J. Y., Gunter, G., and Musgrave, P.1966 Studies of annual abundance of postlarval penaeid shrimp in the
estuarine waters of Mississippi, as related to subsequentcOmmercial catches. Gulf Research Reports, 2(2): 177-212.
3.
4. Eleuterius, L. N.1971 Submerged plant distribution in Mississippi Sound and adjacent
waters. Jour. Miss. Acad. Sel., 17: 9-14.Fuse, S-I., Habe, T., Harada, E., Okuno, R., and Miura, T.
1959 The animal communities in the submerged marine plantvegetations. BulL Mar. BioI. Str., Asamushi Tohoku Univ., 9(4):173-175.
5.
6. Gunter, G.1962 Shrimp landing and production of the State of Texas for the
period 1956-1959, with a comparison with other Gulf states.PubL Inst. Mar.Sci. Univ.Tex., 8: 216-226.
Hoese,H. D. and Jones, R. S.1963 Seasonalityof larger animals in a Texas turtle grass community.
Pub. Ins. Mar.Sci., 9: 37-47.
7.
8. Humm, H. J.1956 Seagrasses of the northern Gulf Coast. Bull Mar. Sci. Gulf Carib.,
6(4): 305-308.Kelly, J. A., Jr., Fuss, C. M., and Hall, J. R.
1971 The transplanting and survival of turtle grass, Thalassiatestudinum, in Boca Ciega Bay, Florida. Fishery Bulletin, 69(2):273-280.
9.
454

10.
ESTUARINE RESEARCH
Marmelstein, A. D.1966 Photoperiodism, and related ecology, in Thalassia testudinum.
M.S. thesis, Texas A & M Univ. 54 p.McMillan, C. and Moseley, F. N.
1967 Salinity tolerances of five marine spermatophytes of RedfishBay, Texas. Ecology, 48: 503-506.
11.
Moore, D. R.1963 Distribution of the sea grass, Tbalassia, in the United States. BulL
Mar. Sci. Gulf Carib., 13(2): 329-342.O'Gower, A. K. and Wacasey, J. W.
1967 Animal communities associated with Thalassia, Diplanthera, andsand beds in Biscayne Bay 1. Analysis of communities in relationto water movements. Bun. Mar. SeL GulfCan'b., 17: 175-210.
12.
13.
19.
Environmental effect on leaves of Diplanthera Du Petit-Thouars.BulL Mar. Sci. Gulf Carib., 10(3): 346-353.
Observations on the ecology and distribution of the Floridaseagrasses. Fla. St. Brd. of Consv. Mar. Lab. Prof. Papers, Ser. no.2: 1-72.
Distribution of seagrasses in Tampa Bay, Florida. Fla. St. Brd. ofConsv. Div. of Salt Water Fisheries, Special Scientific Report No.6:12 p.
On species of the sea grass, Halodule, in Florida. Bun. Mar. Sci.Gulf Can'b., 17(3): 673-676.
Grazing effect on sea grasses by herbivorous reef fishes in theWest Indies. Ecology, 46(3): 255-260.
Ree, W. O.1949 Hydraulic characteristics of vegetation for vegetated water ways.
Agricultural Engineering, 30(4): 184-189.Ree, W. O. and Palmer, V. J.
1949 Flow of water in channels protected by vegetative linings. SoilConservation Technical Bulletin no. 967, U.S.D.A. SoilConservation Service. 115 p.
20.
21. Seibert, P.1968
22.
Importance of natural vegetation for the protection of the banksof streams, rivers and canals. In Freshwater Pub. by Council ofEurope, p. 35-67.
Stephens, W. M.
1966 Life in the turtle grass. Sea Frontiers, 12(5): 264-275.Stevens, H. R., Jr.
1960 A qualitative survey of floral types in Corpus Christi Bay. TexasGame and Fish commission Marine Laboratory Coastal FisheriesProject Reports. 2 p.
23.
24. Strawn, K.1961 Factors influencing the zonation of submerged monocotyledons
at Cedar Key, Florida. Jour. Wildl. Management, 25(2): 178-189.
455
14. Phillips,R. C.1960
15. Phillips,R. C.1960
16. Phillips,R. C.1962
17. Phillips,R. C.1967
18. Randall, J. E.1965

LIONEL N. ELEUTERIUS
25. Tabb, D. and Manning, R.1962 A checklist of the flora and fauna of northern Florida Bay and
adjacent brackish waters of the Florida mainland collected duringthe period of July 1957 through September 1960. BulL Mar. ScLGulf Carib., 11(4): 552-649.
26. Williams, A. B.1958 Substrates as a factor in shrimp distribution. Limnol. Oceanogr.,
3(3): 283-290.27. Wood, E. J. F.
1959 Some east Australian sea grass communities. Proc. LinlL Aoe. N.S. W., 84(2): 218-226.
Voss, G. L. and N. A.1955 An ecological survey of Soldier Key, Biscayne Bay, Florida. BulL
of Mar. ScL Gulf Carib., 146: 204-229.
28.
456


















