
0 Gallons: Epiphytes Contextualized For Landscape Architects
73
0 Gallons: Epiphytes Contextualized For Landscape Architects. Brandon Cornejo LA 461-02 Dave Watts
-
Upload
brandon-cornejo -
Category
Documents
-
view
221 -
download
2
description
Senior Capstone Project
Transcript of 0 Gallons: Epiphytes Contextualized For Landscape Architects

0 Gallons:Epiphytes Contextualized For Landscape Architects.
Brandon CornejoLA 461-02Dave Watts


1.
IntroductionPurpose / Project Synopsis
Literature Review
Vascular and Non-Vascular EpiphytesWater and EpiphytesColonization and EpiphytesTree Architecture and EpiphytesEnvironmental Changes and EpiphytesEpiphytes and their BenefitsEpiphytes and DesignReferencesImage Sources
Experiment
Background / Experiment Summary / SubjectLimitations / DelimitationsExperiment ControlsCuttingsAttachment Locations Surface OrientationCare / DocumentationGrowth Data Table Results of Experiment / DiscussionExperiment Documentation Photos
Expert Commentary
Conceptual Images
25
81214182022242930
3234353637383940424344
64
67
Table of Contents

Introduction If one looks at the world’s diversity of plants and compares that to the amount of plants humans are utilizing in the landscape, one would notice a serious deficiency in the diversity of plant types in the landscape. While trees are heavily studied as large scale ecological servants in urban environments, many other types of plants are critically underutilized. This underutilization stems from the traditional ways landscape architects think about various surfaces. Landscape architects are the designers of a brilliant amount of surfaces. There is distinction, however, between surfaces’ abilities to foster plant life. The first major distinction divides landscape surfaces into hardscapes and softscapes. Traditionally, hardscapes, in comparison to softscapes which are comprised of soils or soil-like media for plant life, have not been designed to host plant life. With increasing development, landscape architects are challenged with density and an increasingly disparate ratio between immpermeable hardscape to softscape landscape areas in urban areas (Elliot 2015). Because of this, landscape architects are often forced to fit trees into 6’X6’ openings along a sidewalk. This example highlights the traditional idea of softscape as void within hardscape that allows for plants to grow. What if we challenged the fact that plants have only been able to grow in softscape-landscaped areas? What if plants could grow on hardscape surfaces?
There is an interesting group of plants, which can provide solutions for this plant-less hardscape challenge. These plants are called epiphytes. “Epi,” the Latin root for “over”, and “phyte,” a plant or plant-like organism, make up ‘epiphyte,’ a term used to describe a large group of many plants from many genera which grow “over” other plants. A definition describes epiphytic plants as plants that “have given up any contacts with water and nutrient-supplying substrates” (Luttge 1989).
Epiphytes, essentially, are plants that can grow without soil or a soil medium. Figure A. displays the aerial roots of a vanda orchid, an epiphytic orchid often horticulturally grown without a growing medium. Unlike mistletoe, a common parasitic plant, which derives nutrients and water from a host tree, epiphytic plants, gain their “epiphytism” from a non-parasitic relationship where it does not derive nutrients and water from the host plant. In other words, the plant uses the plant it grows over, the “host” plant, for support rather than for nutrients. In many cases, however, epiphytes have their way of growing on other things besides plants too. Epiphytic plants obtain “all essential elements and water from the air and produces its own food by photosynthesis” (Shacklette 1973). Interestingly, adaptations to a variety of qualities related to substrate-less habitats, light, and water, give them a variety forms (Luttge 1989).
Their functionality within their aerial habiats, makes epiphytes truly remarkable and important plants. For example, a wealth
Figure A. Vanda Orchid Roots

of small-scale interconnected ecosystems exists in the canopy of a tropical rainforest. This wealth in tropical rainforests has its foundation in the presence of epiphytic plants. Many organisms rely on the presence of epiphytic plants for survival. Water holding abilities and provision of habitat are just a few services that epiphytic plants provide for many organisms that dwell in tree canopies. In certain areas, epiphytes contribute heavily to the percentage of leaves there are on a tree. For example, a tree providing shade cover would provide even more shade cover if there is a presence of epiphytic plants exists. In addition to more leaves, there are more flowers too, increasing energy available to pollinators. Because of these provisions to a variety of organisms, epiphytic plants are considered great indicators of biodiversity. Epiphytic plants, in fact, act as a “diversity amplifier” through their provision of shelter and humidity (Mendez Castro 2014).
Where nature and human developments interact, these plants can be seen growing on man-made surfaces. Because of the ability by some epiphytes to tolerate desiccation, or dehydration, the growth of epiphytes over man-made surfaces, most commonly witnessed on telephone and electrical wires (Figure B) in the tropics, is not uncommon (Claver 1983).This ability to grow directly on surfaces that are not reminiscent of any soil or soil-like media, offers unique, unexplored opportunities to designers.
Research suggests that a determining factor of germination success of epiphytic plants is the architecture of trees which host epiphytic plants (Ruiz-Cordova 2014). This provides a base for the study of epiphytic plants’ preferences of surfaces to interact with. By observing qualities of trees which host epiphytic plants, one could postulate the same qualities replicated in man-made surfaces. Designed natural and unnatural surfaces can be looked into as possible repositories for
which epiphytes can exist. These surfaces could then possibly be emulated in an urban environment as potential hosts for epiphytic plants. Epiphytes, ultimately, could bring life to hardscapes where traditional planted systems have not thrived.
By incorporating surfaces that can host epiphytic plants, greater botanical diversity is feasible without incorporation of additional soil-planted areas. No longer would a tree canopy consist of a monoculture of a single tree’s foliage, but would also consist of interesting assemblages of plants that coexist among the branches of trees. A biodiverse system could blend with urban systems more readily with planning, design, and technology efforts. This study of epiphytic plants hopes to promote further research into these types of plants in order to better connect the urban fabric with a natural one. The intergration of these types of plants offers opportunities for non-human life forms to exist within man-made forms. Epiphyte ability to coexist with urban development could supplement the qualities of open space of urban areas, without the need of soil.
3.
Figure B. Epiphytic bromeliads and tillandsia growing on telephone wires.


Purpose The purpose of this project is to provide a general overview of the diverse world of epiphytes, provide evidence that suggests epiphyte benefits to a human landscape, and question the feasibility of anticipating the colonization epiphytic plants on surfaces of designed landscape infrastructure.
Research will suggest the typologies of surfaces that comprise elements within landscape infrastructure to promote and enhance epiphytic establishment on these surfaces. Research will also suggest the feasibility of alternatively designed landscape systems as a “host” for epiphytic plants.
5.
Project SynopsisThe project consists of three parts: a literature review communicating epiphytic plant ecology by describing landscape-associated relationships, an experiment testing the growth of particular epiphyte cuttings while attached to various hardscape materials, and a summary of an interview and nursery tour from Anyd’s Orchids, a nursery known for their epiphytic orchid production.


Literature Revie
w
7.

Figure 1.

There are two broad terms used to identify plants at a large scale level: vascular and non-vascular. This categorization is especially helpful in conversations regarding epiphytes. This distinction helps communicate an epiphyte’s general scale, habitat range, and water requirement.
Vascular plants are plants that use specialized “plumbing” called xylem and phloem to transport water and nutrients throughout the plant. Ten percent of the world’s vascular plants are epiphytic (Zotz 2009). While there are some outlier species of epiphytes that grow outside of the tropics (mostly ferns), most of the world’s vascular epiphytes are concentrated in the tropics and within “noteworthy temperate ‘hotspots’ in New Zealand, Chile, and the Himalayas” (Zotz 2009). These plants have evolved vastly since their ancestral beginnings as algae, to have diverse functional forms (See Figure 1). What makes vascular epiphytes interesting are the forms and processes which are each unique from each other. For example, the image to the left shows a orchid cacti, with succulent leaves to store water, while the two bromeliads pictured have incurved leaves which capture and hold rainwater. The genus tillandsia, a group which are commonly referred to as airplants offers a brilliant amount of forms through which adaptations have manifested themselves in speciation and form. The Tillandsia durattii have recurving leaves which, with age, “dessicate, shrinking into tight ringlets around a convenient twig or
Vascular Epiphytes and Non-Vascular Epiphytes
Figure 2. Tillandsia duratii’s recurving leaves help support itself on this shrub.
Figure 3. Tillandsia latifolia, a viviparous tillandsia, offsetting, growing in a “biodiversity hotspot.”
9.


Figure 6.
A fern, a vascular
epiphyte, growing on
moss, a non-vascular
epiphyte in Limekiln State
Park (CA).
branch” (Isley 2009) (See Figure 2). Tillandsia latifolia has evolved to embrace a unique form of asexual reproduction called “viviparity,” a term used to describe tillandsia which bear offsets on their inflorescences (Isley 2009) (See Figure 3). A less evolved and, hence, less varied in form group of plants are the non-vascular plants. Most non-vascular plants are epiphytic. Examples of non-vascular plants include lichen (for the purposes of this research project we will classify lichen, a symbiotic relationship between fungi and algae, as an epiphyte) and mosses. Non-vascular plants have not evolved as much as vascular plants from their ancestral forms as algae. Non-vascular plants are typically small, simple plants with few cells. Despite their simple nature, non-vascular plants are some of the most robust plants in the botanical world because of their ability to dehydrate and revive, a trait that most vascular plants do not have. Non-vascular epiphytes are much more efficient at fog or water absorption than vascular epiphytes (Stanton 2014). Their range includes varied occurrences in the polar treeline to heavy occurrences in the tropics (Zotz 2009). Thusly, there are non-vascular epiphytes which can live in frigid temperatures and non-vascular epiphytes which can take scorching temperatures as well.
Despite their notable differences,
Left: Figure 4. Licken in Big Sur, CA. Above: Figure 5. Moss gorwing over bricks in shade.
vascular epiphytes and non-vascular epiphytes are commonly linked in biological relationships. In many cases non-vascular epiphytes provide a necessary surface for vascular epiphytes to germinate. Mosses and lichen add texture to areas of tree bark where vascular epiphytes may not be able to germinate (See Figures 4 and 5 for non-vascular coverage). Research regarding the importance of non-vascular epiphytes to the establishment or recovery of an ecosystem founded by the presence of epiphytic plants suggests the non-vascular epiphytes as crucial first step to a more biodiverse succession of epiphytic plants (Acuna-Tarazona 2015). The textural qualities of non-vascular epiphytes make them great surfaces for which vascular epiphyte seeds can germinate, providing humidity and structural support for vascular epiphyte seedlings. A “non-vascular epiphyte layer could facilitate the recovery” of a vascular epiphytic plant community far faster than a recovery without a non-vascular plant foundation (Acuna-Tarazona 2015). A study about the germination of epiphytic orchid species observed successful germination where moss succession had occurred (Kindlmann 2014). Non-vascular plants and vascular plants are essentially linked in many epiphytic systems.
Taking a step back, an epiphyte ecosystem may not essentially link a non-vascular epiphyte with a vascular epiphyte counterpart, but the coexistenece of both in similar vascular epiphyte habitat is obligatory (See Figure 6).
11.

Figure 6. Illustration of the biodiversity a bromeliad can

Standing water within some bromeliads is enough to host a plethora of organisms, which themselves can act as pollinators. Tree frogs, for example, can begin their lives, as a tadpole, within the interior of a bromeliad and consume other organisms also supported by epiphytes (See Figure 7). Notably, the conversations among plant hobbyists from bromeliad societies around the world, who recreationally grow bromeliads, discuss the problem of mosquitos in bromeliads that hold on water for periods long enough to sustain the life processes of mosquitos.
In a study regarding tropical cloud montane forest epiphyte’s strategies for survival, data collection finds the ability of epiphytes to absorb water from the atmosphere plays an “important role in nutrient cycling and water cycling” and, therefore, impacts regional hydrology and other forest processes (Gotsch 2015). This also reveals an independent ecological system prompted by the presence of epiphytes that is able to utilize resources (water and nutrients) that would not be immediately available to terrestrial systems. Because of epiphytes’ “High water storage capacity” and their contributions to a tree’s surface area, cloud, fog, and rainwater “interception” is increased, maximizing “inputs of water to the ecosystem” (Gotsch 2015).
Water & Epiphytes Because of their unique habitats, epiphytes “have given up any contacts with water and nutrient supplying substrates” (Luttge 1989. Epiphytes have a distinct relationship with water availability. Water availability, in fact, is a determinant of “epiphytic diversity and abundance” (Zotz 2009). Water collection abilities of epiphytes does not only benefit the plants, themselves, but also other organisms.
There are two main characteristics of epiphytes that allow them to survive without access to water-holding substrates. The first of them is their focus with foliar absorption of water, the uptake of water through leaves. This uptake of water through its leaves counteracts the water losses from transpiration (Gotsch 2015). Plants such as those in the genus Tillandsia have leaves covered with specialized hairs called trichomes which trap humidity and condense it to water available to leaf surfaces to uptake and store in their “hypodermis (water storage tissue)” (Isley 2009). The second is a plant’s specialization in water storage (Gotsch 2015). Plants with thick leaves and stems like orchid cactus, orchids, rhipsalis, are characterized by succulence, a term used to describe water storage within plant organs i.e. leaves, bulbs stem.
Water availability in the canopy of trees, through epiphytic plants, allows certain species the ability to spend their entire lives without leaving the canopy of trees. Some bromeliads, called tank bromeliads, have a unique arrangement of leaves, which help to capture and hold water (See Figure 6). Certain species of the tank bromeliads are able to carry up 20 gallons of water (DeChirico 2015). According to a Paul Isley, owner of Rainforest Flora Inc., a nursery specializing in epiphytic plants, epiphytes in certain locations throughout the world “provide the only source of water for part of the year” (Isley 2009).
Figure 7.Two tree
frogs find home on the
incurved leaves of this bromeliad.
13.

Colonization and Epiphytes: A survey in a tropical montane cloud forest in Costa Rica concluded that epiphytes comprised “30% of the total foliar biomass and 45% of the nutrient capital” (Gotsch 2015). While soil areas are quickly consumed by terrestrially linked plants, epiphytes take advantage of volumes of space beyond those that have access to soil. The term “colonization” describes the encrusting growth habits of epiphytes.The image to the right (Figure 8) displays bromeliads and orchids colonizing a tree trunk. The ability to colonize an aerial surface differentiates epiphytes from all other groups of plants.
The success of colonization begins with the success of dispersal mechanisms. Whether sexually, through seeds or spores, or asexually dispersed, dispersal of epiphytic offspring is a highly localized and usually “over short distances by already established species” (Bartels 2012). The presence of “old-growth stands and remnant trees” in serve as repositories for “propagule sources,” in other words, existing old and large host trees and their attached epiphytes serve as mother plants from which new offspring on new surfaces are generated (Bartels 2012). The distance of new, uncolonized habitat from existing, colonized habitat would contribute to the rate at which colonization of new habitat occurs.
Success of sexual reproduction is also determined by chance. An epiphyte’s seed or spore must be intercepted by an appropriate surface. One common characteristic among epiphytes is an emphasis on sexual reproduction. Their flower show can be spectacular, and seeds, accordingly, plentiful. This flower show often takes a toll on the plant; with all its energy going directly to flower and seed production. Sexual reproduction of
Figure 8. An assemblage of epiphytes of various genera and scales grow in niches over the bark of a tree.

Independent Bromeliad
Independent Bromeliad
Stolon
many seeds is essential as many seeds are not intercepted by appropriate surfaces that support the germination of the seeds the future life stages of the species.
Similar to human colonization, colonization among epiphytic plants involves competition, because real estate in the air is limited. Competition between species is rigorous as trees have been noted to host upwards of fifty different species of epiphytes (Lopez-Villalobos 2008). Figure 1 shows aquite a diversity of species on a over a relatively small area. As the name suggests, plants consume space along a host plant while it grows. While volumes of air in the correct habitat are seemingly limitless, surfaces with access to appropriate epiphytic habitat are not. Despite this a lack of space, certain epiphytes have abilities to increase growing space beyond the surface area of a tree.
Many epiphytes are considered monocarpic, a life process which ends with a flower, most recognizably showcased by agaves with their pinnacle flower at the end of their lifespan. While one flower may produce thousands of seeds, monocarpic epiphytes
also rely also on asexual reproduction via processes like pupping or offsetting, to create increasing colonies, also similar to agaves yet without attachment to soils.
Asexual reproduction through stoloniferous offsetting, the asexual generation of new offspring through horizontal stems called stolons, is a way a variety of epiphytes asexually reproduce. This reproduction process enables epiphytes to hang from themselves, grow off host surfaces, and even upside down. These qualities enable epiphytes to continue to grow despite a lack of inhabitable surfaces. Prolific growth of a “colony” eventually becomes a situation where individual plants which are connected to a colony are no longer attached to the host plant, but are attached to other individuals that are attached to the host plant. A clear example of this would be stoloniferous epiphytic bromeliads. Their offsets do not typically attach to host plants, but are only connected indirectly through the original individual. See Figure 9 and 10 for bromeliad stoloniferous reproduction.
15.
Figure 10: Stoloniferous bromeliad colony. Figure 9: Stolon Diagram

Another example in which epiphytes transcend the limits of host surfaces utilizes the leaf litter from host trees. Accumulations of debris within the host plants surfaces provides niches for more hydrophilic, or water-loving, epiphytic plants to establish. There are unique subgroup of epiphytes called “trashbasket epiphytes” which utilize their own old leaves and leaves of their host plants to increase the surfaces in which they can grow (Benzing 1990). Many trashbasket epiphytic ferns, such as staghorn ferns,
Figure 11. Images of Grammatophyllum elegans’supward-growing roots collect debris and creating a “basket”.
grow in size because of an accumulation of old dead leaves. Accumulated leaves decay and actually provide a nutrient-poor, soil-like media that stores water and increases humidity within its surrounding area. An orchid called Grammatophyllum elegans, is notable for its upward growing roots, which enable leaves to accumulate into the upward facing roots, eventually increasing humidity as collected leaves decay (Figure 11). It is not uncommon to see epiphytes growing in the decayed material of other epiphytes. As epiphytes increase in size, so do the surfaces which epiphytes can grow on, as epiphytes
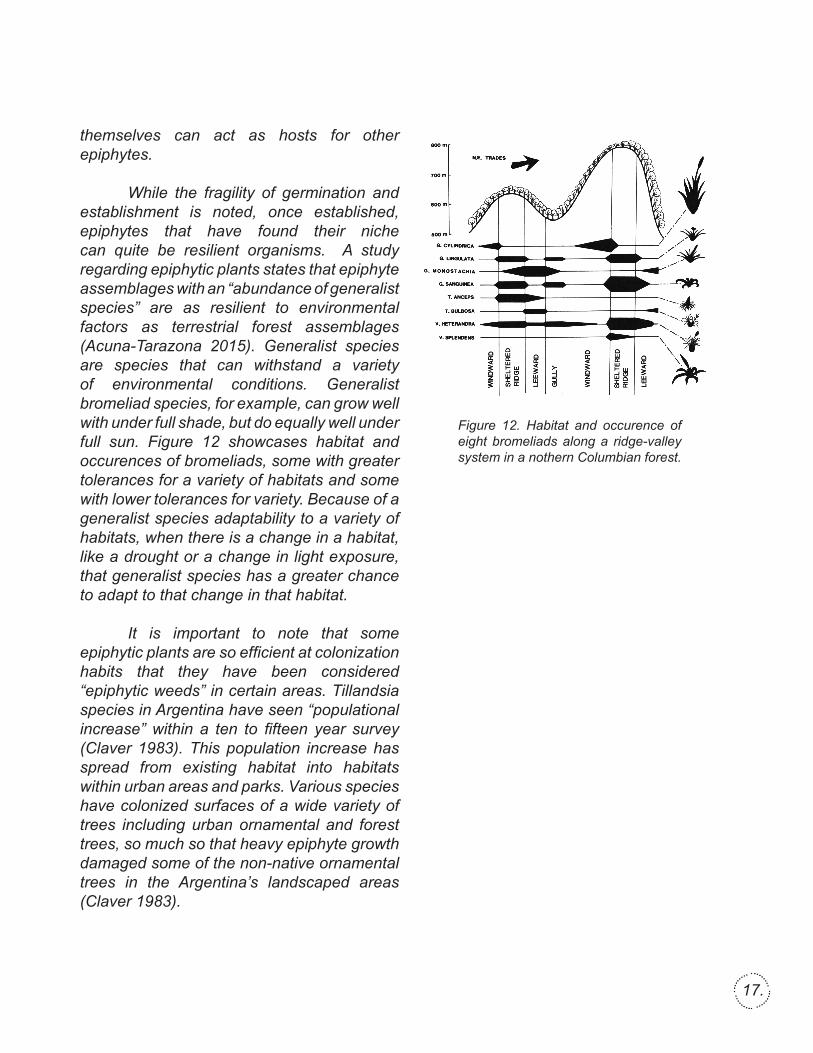
Figure 12. Habitat and occurence of eight bromeliads along a ridge-valley system in a nothern Columbian forest.
17.
themselves can act as hosts for other epiphytes.
While the fragility of germination and establishment is noted, once established, epiphytes that have found their niche can quite be resilient organisms. A study regarding epiphytic plants states that epiphyte assemblages with an “abundance of generalist species” are as resilient to environmental factors as terrestrial forest assemblages (Acuna-Tarazona 2015). Generalist species are species that can withstand a variety of environmental conditions. Generalist bromeliad species, for example, can grow well with under full shade, but do equally well under full sun. Figure 12 showcases habitat and occurences of bromeliads, some with greater tolerances for a variety of habitats and some with lower tolerances for variety. Because of a generalist species adaptability to a variety of habitats, when there is a change in a habitat, like a drought or a change in light exposure, that generalist species has a greater chance to adapt to that change in that habitat.
It is important to note that some epiphytic plants are so efficient at colonization habits that they have been considered “epiphytic weeds” in certain areas. Tillandsia species in Argentina have seen “populational increase” within a ten to fifteen year survey (Claver 1983). This population increase has spread from existing habitat into habitats within urban areas and parks. Various species have colonized surfaces of a wide variety of trees including urban ornamental and forest trees, so much so that heavy epiphyte growth damaged some of the non-native ornamental trees in the Argentina’s landscaped areas (Claver 1983).

Diagram showing the distribution of epiphytic orchids on photophytes in a West African rain forest. The number of orchid species found in the different zones of the phorophytes are given as a percentage of total number of species registered (101). A. Basal parts if the trunk up to 3 m above ground level. B. The trunk from 3 m above ground level up to the first ramifications. C. The basal parts of the large branches (1/3 of total length). D. The middle parts (1/3). E. The outer parts (1/3) of the large branches. (After Johansson 1975)Springer-Verlag 1989 148
Tree Architecture and Epiphytes: While the characteristics that enable epiphytes to grow in and contribute to an aerial ecosystem, the structure of their hosts, plants that epiphytes grow on, is critical to epiphyte diversity.
A study on epiphytic bromeliad distributions within a tropical dry forest states that a “forest that is vertically heterogeneous has more microhabitats” and, therefore, more opportunities for habitats for more “plants and animals” (Ruiz-Cordova 2014). These “microhabitats” are the product of “niche partitioning” which “[enables] the coexistence of a wide diversity of epiphytes” (Ruiz-Cordova 2014). The vertical stratification of types of plants and animals is visible throughout the layers of the forest. The higher in the forest a microhabitat is, “light availability vapor pressure deficit and wind speed generally increase” and therefore have a “considerably drier” microhabitat than lower microhabitats
in the understory (Ruiz-Cordova 2014). Besides the vertical stratification of forests, the architecture of trees contributes to the “vertical distribution of epiphytes among different forests” (Ruiz-Cordova 2014).
Characteristics of tree architecture including “size, age, branch quality, and bark texture” contribute to epiphyte distribution (Bartels 2012). For example, trees “have few large woody structures at the bottom and many smaller structures at the top” informing more surface area and a higher proportion of epipytes in the canopy area of trees (Riuz-Cordova 2014) (See Figure 13. for proportions of epiphyte distribution throughout a tree). Because trees are constantly growing, the dynamics of the forests are also changing as well. A change in the scale “of twigs or branches to shorter tree longevity” produces “immediate consequences for dependent flora” (Zotz 2009). As a tree ages, branches
Figure 13.

grow, and, therefore, the surface area increases, increasing habitat available for epiphytes. See Figure 14 for changes in epiphyte distribution throughout life stages of a tree.
When trees die, a decrease in habitat would follow. Noted, as well, is instability of “substrate,” in other words, instability the bark of an epiphytes host plant, is the “major cause of mortality” among epiphytes (Bartels 2012). For this reason, trees with peeling bark or branches that may break due to weight of epiphytes are also tree characteristics that influence epiphyte distributions. A diversity of trees is key to epiphyte diversity as some epiphytes have preferences for “particular host and host traits” (Bartels 2012).
A paper on mechanisms that regulate epiphyte diversity states that a “loss of preferred host species could translate into the loss of diversity” (Bartels 2012). On the contrary, the introduction of preferred host species could translate into an increase of diversity. A “heterogeneous” patterning of tree forms, meaning a patterning of trees with a variety of forms and the presence of openings in dense tree canopy, is an indicator of areas with high epiphyte diversity. The heterogeneity of a forest sections is associated with heterogeneity of microclimatic conditions within forest sections. Heterogeneity of tree species is the also an indicator of epiphyte diversity as “various tree species provide diverse substrates for colonization” (Bartels 2012).
Figure 14. Diagram showing the life stages of a tree. The dotted line represents areas where branches would fall off naturally. The black dots represent instances of epiphytes.
19.

There is a strong correlation between the existence of epiphytes and biodiversity. Epiphytes, according to a study regarding plant diversity, “constitute an important bioindicator group of species,” indicating the overall “ecosystem health and productivity” (Bartels 2012). Roles in forest hydrology, nutrient cycles within the forest, and the facilitation of animal life in tree canopies make epiphytic plants a crucial component to process which support biodiversity (Zotz 2009). Because epiphytic plants, themselves, are a “species-rich” life form there are opportunities with their own diversity to add to the biodiversity of an existing ecosystem. Because of niche relationships in which certain organisms only favor certain plant types, incorporating those plant-types brings in niche habitat for those certain organisms.
Changes in the landscape have consequences that impact epiphytic plants. Landscape changes from the scale of landuse changes to the scale of global climate changes will reveal a variety of responses.
Landuse Changes
Fairly similar to the presence of canopy gaps in forest sections, disturbances highly regulate epiphyte diversity. Disturbance events “disrupts prevailing microclimatic conditions in habitat” which disrupts in the prevailing epiphytic assemblages within those existing habitats (Bartels 2012). Disturbance can go either ways in terms of biodiversity in all species. For example distance in which a gap is created within the tree canopy would let more light into a uniformly shaded habitat, therefore, introducing possibilities for epiphytes more accepting of light. A disturbance, however, where there are removals of trees, actually is habitat loss, so diversity is inherently minimized, because of a loss of species. A loss in habitat limits introduces scarcity. Like all organisms, limited resources are associated with competition. Competition for water, light exposure, and most related to epiphytic plants, surfaces to grow over are determining factors for epiphytes. Destruction of epiphytes habitat,
Environmental Changes and Epiphytes:

especially in the tropical rainforest, is, also, the destruction of habitat for a plethora of other genera of organisms.
Global Climate Change
On a much larger scale, global climate change occur at a much slower rate than drastic landuse changes. Three options are available to plants in a changing climate: the first is to “remain stationary and evolve in situ;” the second is to “track appropriate niches spatially and migrate;” the third is to “go extinct” (Zotz 2009). Figure 15. showcases predicted movements as climate warms and potential losses where there are no habitats to move to. Tree loss, however, as stated previously, is a threat to epiphyte biodiversity. An increase in global tree loss would result in “the loss of preferred host species, propagules, and suitable [substrates]” (Bartels 2012). Microclimate change is also a determining factor in epiphyte biodiversity. With global climate change certainly impacting microclimate change, mortalities of various
tree species, global climate change impacts epiphytic plants. In a study regarding global climate change and cloud forests, global climate change was noted to impact “both the hydrology and ecology” of cloud forests (Jarvis 2010), a critical habitat for significant proportion of epiphytic life forms. Climate affects “forest type” and forest “stature” directly as well as indirectly “through its effect on soil properties” by water-logging and impacting its nitrogen content (Jarvis 2010).
Figure 15. How will climate change affect regions at the extreme ends of temperature or moisture conditions? Expressed here is an example: in tropical lowland forests, no species from warmer zones are avaliable to replace those species that may disappear, while in subalpine forests, no wetter/cooler forest habitats are available for species to move to.
21.

Despite their relative scale to larger landscape plants such as trees, epiphytes, benefit the natural landscape, people, and to man-made landscape systems. Regulatory roles in natural landscapes, contribution to a horticultural economy, and efficiency in human processes are just a few documented benefits of epiphytes.
In a study regarding epiphytes relationships to microclimate, a particular host plant, Eulychnia saint-pieana, was “significantly cooler and more humid during the day” and “slightly warm at night” than an another saint-pieana plant without (Stanton 2014). In areas of intense sun, epiphytes that grow in these high-light situations benefit host plants through photoprotection, shading aspects which protect host-plant epidermal tissues (Stanton 2014). Epiphytes, through this experiment, were noted to “influence the water economy of their host plants” (Stanton 2014). This instances of protection of hosts by epiphytic plants It is noted that epiphytes constitute up to “50% of the tree biomass” increasing foliage on the tree dramatically (Luttge 1989). Because epiphytic plants are essentially linked to host trees, host trees benefit from those the epiphytic relationship. A host of epiphytes would require less resources to support, yet have provide increased environmental services.
In addition to their environmental value, epiphytes also are notable for their various values to people around the world. While CITES has banned the exportation of wild plants, “epiphytic species are [still being] harvested from the wild for trade as ornamental plants” and for nutritional, medicinal and ceremonial purposes (Acuna-Tarazona 2015). Bromeliad and orchids
are “under high pressure due to extraction for trade and religious purposes” (Mendez-Castro 2014).
Many epiphytes are highly coveted by plant enthusiasts around the world as showcased by the popular orchid shows around the world (Figure 16). While wild-collect specimens were once common, a wild collected epiphyte purchase is usually an illegal transaction and a rare opportunity. With the correct permits, however, nurserymen are allowed limited collection of seeds. Because of the nature of many epiphytes, seeds are plentiful, and it is relatively easy to start growing many of them. In contrast, horticulturalists have streamlined the production of orchids within the orchid industry through tissue culture, an asexual propagation technique that provides more consistent, uniform, and reliable plants compared to less-reliable reproduction through seed.
Many epiphytes, horticulturally, are viable, reproducible commodities. Airplants, for example, are reproduced in greenhouses on pieces of window screen fabric as they require light and air to germinate. The orchid industry, particularly among Phalaenopsis
Epiphytes and their Benefits:
Figure 16. Horticultural display of orchids at the International Santa Barbara Orchid Show.

(Mendez-Castro 2015).
In a study about shade-coffee plantations’ relationship to epiphytes, a “high epiphyte load” was found to contribute to the ecological benefit of growing coffee in this manner. This interplay of human cultivation and natural systems produces a notable benefit to the promotion of biodiversity. Interestingly, the number of epiphytes was actually higher in shade-coffee plantations within cloud forest areas than in uncultivated cloud forest areas (Mendez-Castro 2015). This ability to coexist highlights a symbiotic relationship not only between epiphytes and their cultivated hosts, but also a relationship between epiphytes and human beings. See figure 17 for various coffee-growing systems in utilized in Mexico.
orchids, has streamlined production from tissue culture to a plant ready for storefronts in a relatively efficient manner. Epiphytes also take fertilizer and respond to them significantly, further shortening the timeline of their production.
While intentional absorption of fertilizer, or chemicals, increases production efficiency in the horticultural industry, epiphytes are also able to absorb pollutants in the air. Researchers looking into air quality of urban areas have used Spanish moss, an epiphytic bromeliad, common in the south eastern United States, to estimate “the degree of atmospheric contamination” (Shacklette 1973). The ability of epiphytic plants to filter contaminants mirrors that of plants that grow in soil. Researchers, however, prefer the use of epiphytic plants like Spanish moss and air plants, because they obtain their nutrients directly from the air. Plants with a greater “foliar surface in proportion to their weight” filter air than other plants because their “large surface area favors increased absorption and surface deposition of the airborne materials” (Shacklette 1973). Using plants, which require soil, would give flawed data as elements are absorbed from soil and water, rather than simply the air (Shacklette 1973).
Human intervention into natural systems does not necessarily have negative impacts to epiphytic diversity. Where humans have deforested areas of which once served as habitat for epiphytes, epiphytes are able to colonize “woody perennial, such as cacao, coffee, citrus, and some timber species” that replace original host trees (Zotz 2009). Shade-grown coffee, for example, is an ecologically sensitive method of growing coffee plants under the canopy of existing trees. Because coffee is grown best in “the geographic distribution of endangered ecosystems as tropical rain forest and montane cloud forest” coffee plantations often intersect with the habitat of epiphytic plants and their hosts
23.
Five coffee -growing systems in Mexico showing opportunities for epiphytic plants to coexist with agriculture. (Moguel 2009)

Landscape designers are constantly innovating new ways to create special places for people. Epiphytes are a unique life form, which naturally thrives in an aerial environment, yet can adapt to the necessary forms of human infrastructure. An image (Figure 18), taken by Patrick Weygoldt, captures an instance where human infrastructure hosts epiphytic plants, providing initial evidence for the potential hybrid role of epiphytic plants in the landscape.
The scope of the notable project, organized by SCAPE Landscape Architecture PLLC, Oyster-tecture, challenged the “domain of landscape design” when landscape architecture designed marine landscape infrastructure (Orff 2015). The project envisioned a polluted canal as a “seedling ground” for an oyster reef (Orff 2015). The use of marine life forms, in Oyster-tecture’s case, to create infrastructure is not only beneficial to the environment, but also, to the people who use that infrastructure. As the project redirects focus from the land to the sea, a redirection of focus from the land to air is of similar nature. Like oysters, epiphytes encrust surfaces. However, instead of encrusting surfaces within marine environments, epiphytes encrust surfaces within aerial environments. They can do this because epiphytes, in their natural habitats, obtain “all essential elements and water from the air” (Shacklette 1973). This unique ability offers great potential for landscape architects as they service areas where soils or soil-like media is prohibited.
It is not only important to preserve these plants for their contributions to global biodiversity, but also opportunities to harness the qualities of epiphytic plants as their habitats shift as climate changes. The best
way to harness the qualities of epiphytic plants is their utilization in designed landscapes. A study relating native epiphytic plants to urban areas in Singapore reveals the “viability of urban areas as repositories for biodiversity” (Izzuddin 2015). The best way to harness the qualities of epiphytic plants is their utilization in designed landscapes. As climate changes and natural environments decline, urban areas which are maintained by people can save some of this biodiversity. Similarly, to a botanical garden which preserves a biodiverse selection of plants, designed and maintained areas are opportunities for the same type of preservation. If landscape architects begin to consider epiphytes as landscape materials, the contextualization of epiphytes for the landscape architect would fundamentally precede those considerations.
The concept of epiphytic plant materials should follow the same principals which guide the selection of terrestrial plant materials. While designers often have their go to, fool-proof, robust plants that are plantable from the most inhospitable soils to more average soils, designers utlizing epiphytes should develop knowledge of good epiphytes to use. A conversation with Andy Phillips, a renowned grower of epiphytic orchids, reveals a great variety epiphyte very sensitive epiphytes to a hardy epiphytes, which would vary If the range of generalist epiphytic species, epiphytic species with a broad range of tolerances, intersects with human boundaries, the
Epiphytes and Design
Figure 18. Epiphytes growing on a telephone pole in

intersections of epiphyte habitat and human habitat can be coordinately designed. In a study regarding people’s perceptions of trees, “Oxygen …shade, aesthetics, weather regulation, and environmental quality improvement” were the main benefits that the survey group equated with trees and “garbage generation, dangerousness, and age” were the main problems associated with trees (Camacho-Cervantes 1999). On a different scale, epiphytes can provide similar benefits to those provided by trees without some of issues that trees provide in urban landscapes in terms of their size and their necessary connections to the soil. Epiphytes can act as an alternative to a tree canopy, where trees are not feasible.
Singapore’s Supertrees (Figure 19), designed by landscape architects of Grant Associates, environmental designers of Atelier Ten, and architects from Wilkinson Eyre, incorporate epiphytes into a highly manicured, irrigated, yet soil-less system. The super trees are “concrete and steel structures that form as part of the conservatories’ conditioning systems as well as the overall site energy management systems” which incorporate vertical gardens (Davey 2011). These vertical gardens are soil-less systems, which are vegetated with epiphytic plants. The name ‘supertree’ emphasizes the scale of this fabricated tree. Towering over a canopy created by real trees, this structure emphasizes the ability of epiphytic plants to grow where other plants cannot. The towers are significantly taller than surrounding exotic and native trees. With man-made structure and irrigation epiphytic plants grow well above the canopy of surrounding trees. The interplay of designed structures and natural elements, in this case, epiphytic plants, conditions designers to foster biodiversity. If the surfaces of trees are limiting factors of epiphytic habitat, recalling their ability to grow over non-natural materials, surely the creation of surfaces available for these plants would extend epiphyte habitat.
25.Figure 19. Epiphytes on Singapore’s Supertrees

Top: Figure 20. Dugmore Design Studio’s tree house incorporates real epiphytic plants.
2 Above: Figure 21. CMG Landscape Architecture’s installation process of SFMOMA’s lichen garden.

27.
While Singapore’s Supertrees enjoy the luxury of a rather tropical climate, installations, which involve epiphytic plants, have also seen success in temperate climates. In San Diego’s Botanic Garden’s Hamilton Children’s Garden by Dugmore Design Studio has a tree house play structure which they have incorporated durable epiphytic plants, including sun-tolerant and drought bromeliads, orchids, and staghorn ferns (See Figure 20). The success of this installation is due to the designer’s plant selection methods of irrigation (a drip micro spray). Through irrigation, epiphytic plants are able to grow in dryer climates than they are accustomed to, like other horticulturally exotic plants. Just like the irrigation of plants in traditional landscapes, the maintenance of epiphytic plants within designed areas will probably involve some irrigation, if at least during establishment. CMG Landscape Architecture’s SFMOMA Rooftop Garden incorporates a textured wall surface, which they inoculated with lichen with the instruction of a lichenologist (Figure 21). These examples highlight the importance of understanding the living materials’ needs as well as creating habitats that are similar to their habitats in nature.
Designers of urban green spaces that intersect epiphytic habitat not only anticipate epiphyte growth on the bark of planted trees, but also, on predict growth on man-made surfaces. Recalling the telephone wires covered with epiphytic plants, fabricated materials can act as the host plant surfaces. Because of this, there are opportunities for design to extend existing habitat therefore, create exciting, vibrant, slightly natural and unnatural landscapes. Nevertheless, with all design professions, designers must know their materials. Further research into specific types of epiphytes, their habitat preferences, etc. are essential to creative success with epiphytes. With design, to anticipate this epiphytic growth, epiphytes may coexist with informed and aesthetic planning.


Acuna-Tarazona, M, T Toledo-Aceves, A Flores-Palacios, VJ Sosa, and ML Martinez. “Post-stripping Recolonization of Vascular Epiphytes in Cloud- forest Fragments in Mexico.” Journal of Tropical Ecology, 31 (2015): 499-508.
Bartels, SF, and HYH Chen. “Mechanisms Regulating Epiphytic Plant Diversity.” Critical Reviews in Plant Sciences, 31.5 (2012): 391-400.
Benzing, David H. Vascular Epiphytes: General Biology and Related Biota. Cambridge: Cambridge UP, 1990. Print.
Camacho-Cervantes, Morelia, Jorge Schondube, Alicia Castillo, and Ian MacGregor-Fors. “How Do People Perceive Urban Trees? Assessing Likes and Dislikes in Relation to the Trees of a City.” Urban Ecosystems, 17.3 (2014): 761-773.
Claver, F. , Alaniz, J. , & Caldíz, D. (1983). Tillandsia spp: Epiphytic weeds of trees and bushes. Forest Ecology and Management, 6(4), 367-372.
Davey, Meredith. “Gardens by the Bay: Ecologically Reflective Design.” Architectural Design, 81.6 (2011): 108-111.
DeChirico, Gregg. “Exploring Ecuador: Visions of Ecuadorian Flora and Fauna.” CCCS Meeting. Odd Fellows Hall, San Luis Obispo. 10 Jan. 2015. Lecture.
Elliott, Robert M. Vegetated Infrastructure for Urban Stormwater Management: Advances in Understanding, Modeling and Design. n.p.: ProQuest Dissertations Publishing, 2015.
Gotsch, S. G., Nadkarni, N., Darby, A., Glunk, A., Dix, M., Davidson, K. and Dawson, T. E. (2015), Life in the treetops: ecophysiological strategies of canopy epiphytes in a tropical montane cloud forest. Ecological Monographs, 85: 393–412. doi:10.1890/14-1076.1
Isley, Paul T. Tillandsia: The World’s Most Unusual Air Plants. Gardena, CA: Botanical, 1987. Print.
Isley, Paul T. Tillandsia II : The World’s Most Unusual Airplants. Redondo Beach, Calif.: Botanical Press, 2009. Print.
Izuddin, Muhammad, and Edward Webb. “The Influence of Tree Architecture, Forest Remnants, and Dispersal Syndrome on Roadside Epiphyte Diversity in a Highly Urbanized Tropical Environment.” Biodiversity and Conservation, 24.8 (2015): 2063-2077.
Jarvis, Andy, and Mark Mulligan. “The Climate of Cloud Forests.” Hydrological Processes, 25.3 (2011): 327-343.
Kindlmann, Pavel, Elvia J Meléndez-Ackerman, and Raymond L Tremblay. “Disobedient Epiphytes: Colonization and Extinction Rates in a Metapopulation Of Lepanthes Rupestris (Orchidaceae) Contradict Theoretical Predictions Based on Patch Connectivity: Disobedient Orchids.” Botanical Journal of the Linnean Society, 175.4 (2014): 598-606.
López-Villalobos, Adriana, Alejandro Flores-Palacios, and Raúl Ortiz-Pulido. “The Relationship Between Bark Peeling Rate and the Distribution and Mortality of Two Epiphyte Species.” Plant Ecology, 198.2 (2008): 265-274.
Luttge, Ulrich. Vascular Plants as Epiphytes: Evolution and Ecophysiology. Berlin: Springer-Verlag, 1989. Print.
Méndez-Castro, Francisco, and Dinesh Rao. “Spider Diversity in Epiphytes: Can Shade Coffee Plantations Promote the Conservation of Cloud Forest Assemblages?.” Biodiversity and Conservation, 23.10 (2014): 2561-2577.
Moguel, P, and VM Toledo. “Biodiversity Conservation in Traditional Coffee Systems of Mexico.” Conservation Biology, 13.1 (1999): 11-21.
Nieder, Jurgen, Juliana Prosperí, and Georges Michaloud. “Epiphytes and Their Contribution to Canopy Diversity.” Plant Ecology, 153.1/2 (2001): 51-63.
Orff, Kate. “Shellfish as Living Infrastructure.” Ecological Restoration, 31.3 (2013): 317-322.
Ruiz-Cordova, JP, VH Toledo-Hernandez, and A Flores- Palacios. “The Effect of Substrate Abundance in the Vertical Stratification of Bromeliad Epiphytes in a Tropical Dry Forest (Mexico).” FLORA, 209.8 (2014): 375-384.
Stanton, Daniel E, Jackelyn Huallpa Chávez, Luis Villegas, Francisco Villasante, Juan Armesto, Lars O Hedin, Henry Horn, and Jim Dalling. “Epiphytes Improve Host Plant Water Use by Microenvironment Modification.” Functional Ecology, 28.5 (2014): 1274-1283.
Shacklette, Hansford. “Airborne Chemical Elements in Spanish Moss: Statistical Studies in Field Geochemistry.” Geological Survey Profession Paper 574-E.
Toledo-Aceves, T, JG Garcia-Franco, SL Lozada, MLL Mateos, and K MacMillan. “Germination and Seedling Survivorship of Three Tillandsia Species in the Cloud-forest Canopy.” Journal of Tropical Ecology, 28 (2012): 423-426.
Wagner, K, W Bogusch, and G Zotz. “The Role of the Regeneration Niche for the Vertical Stratification of Vascular Epiphytes.” Journal of Tropical Ecology, 29 (2013): 277-290.
Zotz, G, and M Bader. “Epiphytic Plants in a Changing World-Global: Change Effects on Vascular and Non-Vascular Epiphytes.” Progress in Botany, 70 (2009): 147-170.
References
29.

Bromeliads in Treehouse, San Diego Botanic Garden. Personal photograph by author. 2013.
Weygoldt, Patrick. Telephone Line in Castara, Tobago. Digital image. Facebook. N.p., n.d. Web. 11 Mar. 2016.
Vanda Orchid Photoshop, San Luis Obispo. Personal illustration by author. 2016.
Weygoldt, Patrick. Telephone Line in Castara, Tobago. Digital image. Facebook. N.p., n.d. Web. 11 Mar. 2016.
Weygoldt, Patrick. Telephone Line in Castara, Tobago. Digital image. Facebook. N.p., n.d. Web. 11 Mar. 2016.
Weygoldt, Patrick. Telephone Line in Castara, Tobago. Digital image. Facebook. N.p., n.d. Web. 11 Mar. 2016.
Weygoldt, Patrick. Telephone Line in Castara, Tobago. Digital image. Facebook. N.p., n.d. Web. 11 Mar. 2016.
Various Vascular Epiphytic Plants, San Luis Obispo. Personal Illustration by Author. 2014.
Isley, Paul T. Tillandsia duratii v. saxatilis. Photograph. “Tillandsia II : The World’s Most Unusual Airplants.” Redondo Beach CA: Botanical Press, 2009. P 55. Print.
Isley, Paul T. Tillandsia latifolia ‘Delgado’. Photograph. “Tillandsia : The World’s Most Unusual Airplants.” Gardena CA: Botanical Press, 1987. P 72. Print.
Lichen on branches, Big Sur CA. Personal photograph by author. 2016.
Moss growing on brick, San Luis Obispo CA. Personal photography by author. 2016.
Fern, moss, rock, Limekiln Sate Park CA. Personal photography by author. 2016.
Bromeliad section cut, San Luis Obispo. Personal illustration by author. 2016.
Tree Frogs in Bromeliads, San Luis Obispo. Personal photography by author. 2014.
Laman, Tim. Amazon Rain Forest Canopy View with Flowering Bromeliad Epiphytes Growing on a Branch of a Giant Ceiba Tree.
Neoregelia Zoe. Digital image. N.p., n.d. Web. 11 Mar. 2016. <http://ca-mord.src-nancy.fr/neoregelia-zoe&page=5>.
Isley, Paul T.Neoregelia Donger Ted and Jackie Johnson. Photograph. “Tillandsia II : The World’s Most Unusual Airplants.” Redondo Beach CA: Botanical Press, 2009. P 201. Print.
Rowe, Derrick. Grammatophyllum Elegans, Fiji Islands. Digital image. Forum for Epiphytic Myrmecodia. N.p., 21 Feb. 2014. Web. 11 Mar. 2016.
Rowe, Derrick. Grammatophyllum Elegans, Fiji Islands. Digital image. Forum for Epiphytic Myrmecodia. N.p., 21 Feb. 2014. Web. 11 Mar. 2016.
Rowe, Derrick. Grammatophyllum Elegans, Fiji Islands. Digital image. Forum for Epiphytic Myrmecodia. N.p., 21 Feb. 2014. Web. 11 Mar. 2016.
Benzing, David H. Figure 7.15. Diagram. “Vascular Epiphytes.” New York NY: Press Syndicate of the University of Cambridge, 1990. P 250. Print.
Tillandsia clump, San Luis Obispo CA. Personal photography by author. 2016.
Benzing, David H. Figure 7.15. Diagram. Vascular Epiphytes. New York NY: Press Syndicate of the University of Cambridge, 1990. P 250. Print.
Luttge, U. Fig. 6.5 A-E. Diagram. “Vascular Plants as Epiphytes and Ecophysiology.” Berlin: Springer-Verlag, 1989. P 148. Print.
Benzing, David H. Figure 7.22. Diagram. “Vascular Epiphytes.” New York NY: Press Syndicate of the University of Cambridge, 1990. P 260. Print.
Benzing, David H. Figure 7.21. Diagram. “Vascular Epiphytes.” New York NY: Press Syndicate of the University of Cambridge, 1990. P 258. Print.
Zotz, G, and M Bader. Fig. 1. Diagram. “Epiphytic Plants in a Changing World-Global: Change Effects on Vascular and Non-Vascular Epiphytes.” Progress in Botany, 70 (2009): P 159. Print.
Orchids at the Orchid Show, Santa Barbara CA. Personal photography by author. 2015.
Moguel, P, and VM Toledo. Figure 1. Diagram. “Biodiversity Conservation in Traditional Coffee Systems of Mexico.” Conservation Biology, 13.1 (1999): P 13. Print.
Weygoldt, Patrick. Telephone Line in Castara, Tobago. Digital image. Facebook. N.p., n.d. Web. 11 Mar. 2016.
Barce, G. Singapore: The SUPERTREES! Digital image. N.p., 24 Oct. 2012. Web. 11 Mar. 2016. <http://freakofnaturezzz. blogspot.com/2012/10/singapore-supertrees.html>.
Barce, G. Singapore: The SUPERTREES! Digital image. N.p., 24 Oct. 2012. Web. 11 Mar. 2016. <http://freakofnaturezzz. blogspot.com/2012/10/singapore-supertrees.html>.
Barce, G. Singapore: The SUPERTREES! Digital image. N.p., 24 Oct. 2012. Web. 11 Mar. 2016. <http://freakofnaturezzz. blogspot.com/2012/10/singapore-supertrees.html>.
Ohmygodwin10. Toni’s Tree House. Digital Image. N.p. 20 May 2014. 11 Mar. 2016. <https://ohmygodwin.files. wordpress.com/2014/05/sd-botanic-garden-46.jpg>
Gillin, Jaime. Sfmoma-lichen-installation-shadow. Digital image. World’s First Lichen Garden. Dwell, 26 Oct. 2010. Web. 11 Mar. 2016.
Gillin, Jaime. sfmoma-lichen-installation-closeup. Digital image. World’s First Lichen Garden. Dwell, 26 Oct. 2010. Web. 11 Mar. 2016.
Tillandsia ‘Curly Slim’, Rancho Santa Margarita CA. Personal photography by author. 2015.
Lichen on Rocks, Morro Bay CA. Personal photography by author. 2016.
Images Sources
Sources listed in order as they appear (From Left to Right, Top to Bottom)

31.

Experiment
Background While many factors impact the success of establishment of epiphytic plants, flexible elements of nature including exposure to sun, temperature, rainfall, humidity, would be difficult to replicate. Because of the stability of surface qualities, an experiment where a variety of surfaces serves as the independent variable within an environment with controllable conditions (i.e. water, humidity, light, temperature etc.) is much more feasible. In this experiment, the success of epiphyte establishment, measured by continued growth, and/or attachment to surfaces, would potentially provide evidence that would support the use of landscape materials intended to host epiphytic plants.
Experiment Summary
The experiment will test the growth of Rabbit’s foot fern, Davallia fejeensis, while attached to a variety of surfaces. Growth will be defined as any increases in rhizomes, new leaves, etc.) of the plant while rubber-banded to surfaces.
Subject Rabbit’s foot fern is one of the most widely grown ferns in the horticultural trade. It is an epiphytic fern from Fiji, and is the most commercially available fern from the genus Davallia. It is noted for its fast growth rate, durability, disease and pest resistance and rhizomatous succulence. In current nursery practice, the fern is grown in soil, despite its epiphytic nature. It is reproduced mainly through its rhizomes, the fuzzy root-like appendages which give the plant its common name. The reproducibility of epiphytic plants is an important consideration because wild collection is often disruptive and damaging to ecosystems. In Latin American countries, there is a high demand for the collection of epiphytes which involves careless extraction, via the “stripping” of epiphyte-hosting branches, of individual specimens, leaving “immature plants, which are unused and often simply abandoned on the forest floor” (Acuna-Tarazona 2015). As a horticulturally easily reproducible epiphyte, Rabbit’s foot fern, would be a feasible specifiable plant for landscape projects where site conditions allow.
Info about Davallia from below resources:
“Rabbit’s Foot Fern Culture Sheet.” Arc Ferns, LLC., n.d. Web. 19 Apr. 2016.“Rabbit’s Foot Fern.” House Plant 411 LLC, n.d. Web. 19 Apr. 2016.

Fig. 1 Rabbit’s Foot Fern
33.

Limitations
Geographic Limitation
Landscape applications of epiphytic plants should utilize local flora, as those plants would be most acclimated to the environmental conditions of corresponding sites. Because San Luis Obispo is in an area that is not a biodiversity hotspot for epiphytic plants; i.e. rainfall and humidity is lower comparatively, the experiment selects an exotic plant, Davallia fejeensis, grown in conditions similar to that of its origin, Fiji.
Biodiversity Limitation
In a designed landscape solution, it is likely that multiple epiphyte species would be utilized. Because of the sheer diversity of epiphytes and the niche/ site-specificness of epiphytic plants, however, one experiment focusing on a single species would not provide blanket evidence for the successful attachment of all epiphytic plants, but solely evidence for attachment of the tested epiphytic species. Limitation in terms of access to sources of potential complementary epiphyte plants also leads the experiment to only one plant.
Time Limitation
The timeline of this project spans over 28 days, with observations taken every week. Germination of epiphytes from seeds would surpass the timeline of the experiment. Because of this, an experiment relating to the asexual propagation of an existing epiphytic specimen. As asexual reproduction would visualize attachment in a much shorter time frame.
Delimitations The study of epiphtytic plant establishment to selected surfaces only a part of a greater potential reseach effort to support the idea of a landscape system that incorporates epiphytic plants. The set up of this experiment to keep all variables consistent except those of the selected surfaces, may oversimplify the
This experiment assumes either consistent irrigation or consistent rainfall until establishment, consistent temperature, and consistent light exposure. Nature, however, is not consistent and, therefore, bringing up questions about the fragility of epiphytic systems against fluctuating elements of nature. Wind and air pollution are elements, too, that are not addressed within the greenhouse space.
The timeline of this experiment only allows for the observation of establishment. Landscape plant installations, however, are long-lasting, maintained, and changing elements. More research into the landscape systems which incorporate epiphytic plants within a longer time-frame would provide clarity in terms of expectations. Research into fertilizers of epiphytic plants may enhance potential epiphytic plant systems, something that the experiment does not harness.

Surface Materials There are a total of 10 materials within the experiment to be tested for potential epiphyte hosts. Materials that will be used in the experiment are: cork oak bark, rebar, galvanized sheet steel, steel wire, PVC coated steel wire, welded wire fencing, PVC coated wire fencing, window screen, redwood, and brick. See Figure 2 for materials.
Cork oak bark was selected as a material because of extensive use in the horticultural trade for the mounting of many epiphytic plants. It can be viewed as a control material as it represents actual tree bark that an epiphyte might grow on in nature.
Cork Oak
Galvanized Sheet Metal
RebarSteel Wire
PVC Coated Steel Wire
Redwood
Brick
PVC Coated Wire Fencing Welded Wire
Fencing
Window Screen
Fig. 2 Materials
Surface Type Form
Cork Oak Bark N/ARebar 1/4 in. Galvanized Sheet Steel 1/16 inSteel Wire 3/32 inPVC Coated Wire 152 mWelded Wire Fencing N/APVC Coated Fencing N/AWindow Screen N/ARedwood N/ABrick
Experiment Controls
35.

Cuttings
Cuttings taken consist of a piece of the fern’s hairy aerial rhizome (rhizomes not touching soil) and leaves. Before propagating the plant, the mother plant was watered daily (soil watered and aerial rhizomes watered) to ensure vitalility of cuttings. Cutting uniformity was determined by approximate length and also by approximate number of leaves. Each cutting is around 4-6”, with originally 3-4 fern fronds. Each cutting (20 total), was reduced to its largest and healthiest leaf. The leaves of each cutting were trimmed to 3/4 their original length to reduce water loss through transpiration. See Figure 3.
Fig. 3 Cutting Process Images

Attachment
Cuttings were attached to each of the surfaces via rubber-bands (twisted and/or tied). In current horticultural practice, the mounting of epiphytes to cork oak bark or wood usually involves a compressed sphagnum moss for moisture and humidity purposes. Because of this, two cuttings are attached to each surface type: one with and one without moss. Cuttings are then attached to moss with rubber bands. Cuttings attached with moss are touching both the surface and moss. Moss pieces are held together with rubber bands (See Figure 4). See Figure 5 for attachment example.
Fig. 4 Moss Held with Rubber Bands
Fig. 5 Attachment to steel wire with and without moss.
37.

Fig. 6 Experiment Location
Location
The experiment will take place at the tropical foliage greenhouse of the horticulture unit at Cal Poly San Luis Obispo. The tropical foliage greenhouse is kept under conditions most ideal for tropical plant growth. The allocated is space is against a south-facing wall, so the area is slightly warmer than the rest of the greenhouse. Plants that are growing at the greenhouse include, bromeliads, tillandsia, ferns, sansevieria, pothos, draceana, begonia,
ficus, philodendron, and other tropical indoor plants. Plants growing in the green house receive a thorough watering and misting on a regular basis, keeping the overall humidity of the greenhouse fairly high. Humidity is kept consistently greater than 50%. The windows facing south are painted white for shading purposes. See Fig. 6 for visual.

3’-0”
6’-0
”
2’-0”Fig. 7 Wooden Frame and Metal Fencing
5’-0
”
Surface Orientation:
Each surface to be tested will be hung vertically and oriented uniformly via paper clipson a , 2’ x 5’ piece of metal fencing attached to a wooden frame (Figure 7).
39.

Care
The only care that this experiment involves are two to three mistings daily. Each misting drenches hairy fern rhizomes completely until rhizome hairs are saturated. Water should drip off of rhizomes when fully saturated. There should be a noticeable color change after watering. See Figure 8. to see differences in non-saturated rhizomes and saturated rhizomes.
Fig. 8 Non-saturated Rhizomes vs. Saturated Rhizomes
Documentation
Progess of cuttings will be documented via photographs. Each surface will be photographed adjacent to a ruler prior to that day’s watering/ misting. Notable differences between measurment dates will be notable after review of photographs. Measurements to be taken: April Tuesday 5th, 12th, 19th, 26th, May Tuesday 3rd.

41.

Surface Type GrowthApril 5 April 12 April 19 April 26 May 3
Cork Oak Bark No No No No NoCork Oak Bark w/ Moss No No No Yes YesRebar No No No Yes YesRebar w/ Moss No No No No NoGalvanized Sheet Steel No No No Yes YesGalvanized Sheet Steel w/ Moss No No No Yes YesSteel Wire No No No No NoSteel Wire w/ Moss No No No Yes YesPVC Coated Wire No No No No NoPVC Coated Wire w/ Moss No No No No YesWelded Wire Fencing No No No Yes YesWelded Wire Fencing w/ Moss No No No No NoPVC Coated Fencing No No No Yes YesPVC Coated Fencing w/ Moss No No No No NoWindow Screen No No No No NoWindow Screen w/ Moss No No No No YesRedwood No No No Yes YesRedwood w/ Moss No No No Yes YesBrick No No No Yes YesBrick w/ Moss No No No No Yes
Growth Data Table

43.
Results of Experiment / Discussion:
Within the time frame of 28 days, growth on all surfaces was observed. Observations of growth, with and without moss, was only seen on galvanized sheet steel, redwood, and brick.
The presence of moss did not consistently translate to growth on the different materials being tested. A reason for this inconsistency is likely due to the individual condition of the cutting. If this experiment were to utilize multiple individuals for each trial, growth across all surfaces regardless of moss is foreseeable. The unsuccessful growth while attached to cork oak bark, the material most representative of the natural habitats of epiphytes, further supports this reason for growth inconsistency.
The growth on more than half of the individual cuttings, reveals evidence for the feasibility of rabbit’s foot fern in landscape design where site conditions permit. This experiment indicates that the species is unfastidious in its host surface preferences and would most likely be installable on a wide variety of surfaces. Despite the success of this plant on all surfaces, this may not be applicable to all other epiphytic genera and/or species. Future experiments are needed to test the alternative use of epiphytic plants in the built landscape.

Cork Oak Bark No No No No NoCork Oak Bark w/ Moss No No No Yes Yes
Surface Type GrowthApril 5 April 12 April 19 April 26 May 3
Experiment Documentation Photos
April 5
April 19 April 26 May 3
April 12

April 26
May 3
45.

Rebar No No No Yes YesRebar w/ Moss No No No No No
Surface Type GrowthApril 5 April 12 April 19 April 26 May 3
April 5
April 19 April 26 May 3
April 12

April 26
May 3
47.

Galvanized Sheet Steel No No No Yes YesGalvanized Sheet Steel w/ Moss No No No Yes Yes
Surface Type GrowthApril 5 April 12 April 19 April 26 May 3
April 5
April 19 April 26 May 3
April 12

April 26
April 26
May 3
May 3
49.
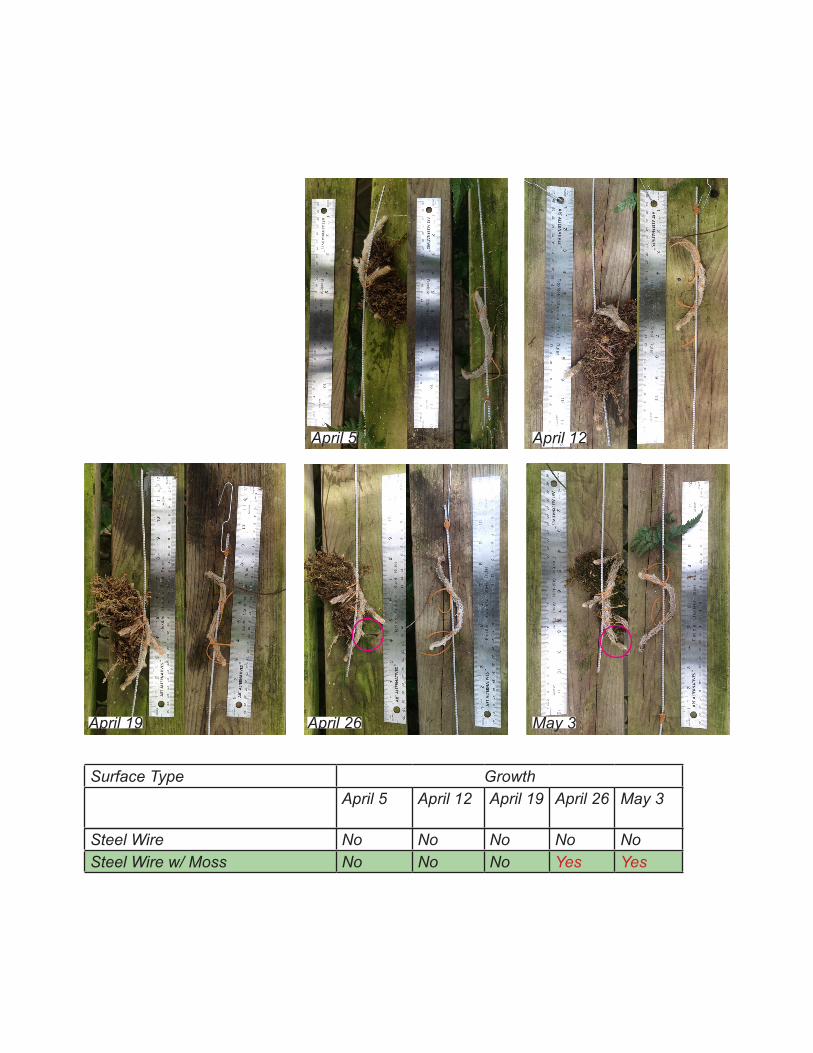
Steel Wire No No No No NoSteel Wire w/ Moss No No No Yes Yes
Surface Type GrowthApril 5 April 12 April 19 April 26 May 3
April 5
April 19 April 26 May 3
April 12

April 26
May 351.

PVC Coated Wire No No No No NoPVC Coated Wire w/ Moss No No No No Yes
Surface Type GrowthApril 5 April 12 April 19 April 26 May 3
April 5
April 19 April 26 May 3
April 12

May 3
53.

Welded Wire Fencing No No No Yes YesWelded Wire Fencing w/ Moss No No No No No
Surface Type GrowthApril 5 April 12 April 19 April 26 May 3
April 5
April 19 April 26 May 3
April 12

April 26
May 3
55.

PVC Coated Fencing No No No Yes YesPVC Coated Fencing w/ Moss No No No No No
Surface Type GrowthApril 5 April 12 April 19 April 26 May 3
April 5
April 19 April 26 May 3
April 12

April 26
May 3
57.

Window Screen No No No No NoWindow Screen w/ Moss No No No No Yes
Surface Type GrowthApril 5 April 12 April 19 April 26 May 3
April 5
April 19 April 26 May 3
April 12

May 3
59.

Redwood No No No Yes YesRedwood w/ Moss No No No Yes Yes
Surface Type GrowthApril 5 April 12 April 19 April 26 May 3
April 5
April 19 April 26 May 3
April 12

April 26
April 26
May 3
May 3
61.

Brick No No No Yes YesBrick w/ Moss No No No No Yes
Surface Type GrowthApril 5 April 12 April 19 April 26 May 3
April 5
April 19 April 26 May 3
April 12

April 26
May 3
May 3
63.


sphagnum moss, fishing line, and the epiphyte propagule. He says that the moss is used mainly for humidity purposes, not really for roots to grow within. His use of chaparral wood is for purposes of longevity, as the oils in the wood works against rot. Through his experience, however, orchids are capable of growing on quite a scale of materials. The only materials he suggests against would be materials with short lifespans and tendencies to peel, such as plastics and easily peelable bark. While this research focused on a asexual form of reproduction, Andy revealed a particular orchids species that he simply dusts its seeds over surfaces and they “grow like weeds.” Throughout his nursery, there are occasional instances of volunteerism, the unplanned germination of plant seeds in areas not intended for epiphytic plants. This observation at his nursery, consistent with the literature review, exibits established populations giving rise to new populations of epiphytes on unpopulated habitats.
The timeline for maturity from a propagule to a flowering-sized plant ranges anywhere from a couple years to many years. It all depends on the species of plants. Because of this there are definite species that would most likely be most feasible in a landscape design which is expected to “fill in” within a reasonable amount of time. Despite this, Andy suggests that slower growing species are worthwhile and can be incorporated with faster-growing species. That being said, Andy concludes that there would definitely be “go-to” generalist species that would be more feasible in a wide variety of landscape scenarios.
Andy Phillips interviewed May 5, 2016
Expert Commentary A conversation with Andy Phillips, a renowned grower of epiphytic orchids, and visit to his nursery, Andy’s Orchids in Encinitas, California, reveals a wealth of opportunity with epiphytic plants.
With a history of time spent in Central and South America, Andy has brought thousands of plant species into horticulture, some of which are endangered in their native habitats. His experiences within epiphyte habitat revealed plants for every habitat scenario. Because of the scope of the nursery, he keeps a variety of greenhouse environments to grow a variety of epiphytes with different environmental preferences. Some orchids are even grown outdoors under within the local weather conditions, as some of the orchids are accustomed to similar climactic conditions of Encinitas.
What enables Andy to grow orchids from all over the world is an understanding of their water requirements. If planted in their natural habitats, these plants would not need supplemental irrigation. When growing epiphytes outside of their habitats, the grower requires a knowledge of their watering preferences. With knowledge of water preferences and modern technology in irrigation, it is possible to grow epiphytes outside of their native habitats, as many exotic terrestrial landscape plants are. Andy says that with technology, design can enable epiphytes to grow beyond the canopy of trees.
Andy’s process of propagation involves a piece of wood (he uses wood from chaparral plants cleared for land development),
65.

With deforestation replacing critical trees in tropical rainforest with an agricultural landscape, design utilizing epiphytes can enable native arboreal ecosystems to exist above.

67.


This parking structure is given new life and aesthetic appeal, through color and texture, with epiphytic plants.
69.


71.
This parking lot without planters to allow for any sort of tree canopy, receives shade and aesthetic appeal through epiphytes.


















